{kind=link}
{kind=link}
{kind=link}
亲本籼粳成分与两系杂交粳稻杂种优势的关系及遗传基础
[于亚辉1, 2 , 刘郁1 , 李振宇1 , 陈广红1 , 徐正进2 , 唐亮2, *  , 毛艇
, 毛艇1 , 徐海2 ]
, 毛艇|
|


第一作者联系方式: E-mail: yyh666@sina.com
以籼粳重组自交系(秋光×七山占, RIL)和粳型光温敏核不育系(GB028S)及其杂交F1为材料, 利用程氏指数法和分子标记法分析亲本的籼粳成分与杂种优势的关系及遗传基础。结果表明, 采用程氏指数法和分子标记法在群体籼粳分类的结果上比较一致; RIL偏粳系数与F1产量及其杂种优势均呈显著或极显著的二次曲线关系; F1产量在偏粳系数0.55~0.70区间内出现高峰值, 杂种优势在偏粳系数0.50~0.65区间内出现高峰值, 即RIL偏粳系数为0.55~0.65时F1有形成较高产量及杂种优势的潜力。Chr.8、Chr.11和Chr.12的籼粳成分与F1产量及杂种优势关系密切, 双亲的遗传距离与F1产量和相关性状及杂种优势没有明显的关系。
, MAO TingIn order to further reveal the relationship between composition of subspecies lineage and heterosis in Oryza sativa L., we built a bridge for morphological index, Cheng’s index (Chi) and molecular index, japonica discrimination values ( Dj) based on two populations, a recombinant inbred lines (RILs), derived from the cross between indica cultivar Qishanzhan and japonica cultivar Akihikari, and japonica photo-thermo sensitive genic male sterile (PTGMS) line GB028S and their F1 hybrids. Two indexs achieved relatively consistent results in two populations independently. Significant curvilinear relationship appeared among Dj values, F1 yield and yield heterosis. The peaks of F1 yield and yield heterosis were detected in Dj interval from 0.55 to 0.70 and from 0.50 to 0.65 respectively, indicating that 0.55-0.65 is the preferential Dj interval for producing high yield and heterosis of F1 hybrids. Compositions of subspecies lineage on Chr.8, Chr.11, and Chr.12 were closely related with yield and heterosis of F1,however, the relationship among parent’s genetic distance, F1 yield and heterosis was not significant.
自1908年Shull提出“ 杂种优势” 以来, 杂种优势在提高作物产量上被广泛利用。水稻特别是亚洲栽培稻主要分为籼稻和粳稻两个亚种, 其遗传分化程度较大, 在育种研究中广泛认为籼粳交具有较强杂种优势。自1951年以来, 沈阳农业大学杨守仁教授团队开始从事籼粳稻杂交育种的基础研究, 认为籼粳交是产生杂种优势的有效途径[1]。经过育种实践, 育成了应用价值很高的沈农98366、沈农127和沈农1033等。在相同理论的指导下, 我国育种家利用籼粳杂交陆续育成多个新品种, 如浙江的矮粳23、T209、城特232等, 辽宁的辽粳5号和辽粳326, 江苏的紫金粳和南粳35, 北京的中作系列, 湖北的鄂育晚5号等[2]。同时, 国外学者对籼粳杂交育种技术亦高度重视, 20世纪60年代, 韩国育种家利用籼粳杂交育成了统一、水原、密阳等高产矮秆品种。日本于1981开始籼粳交育种, 先后育成了中国91、北陆125、关东146等新品种[2]。籼粳杂交理论不仅在常规育种上得到广泛应用, 更是杂交稻育种的重要手段。20世纪90年代, 我国育种家直接利用籼粳杂交强杂种优势成功培育出两优培九、Y两优2号、Y两优900等一系列两系法杂交水稻组合, 其中Y两优900最高单产达14.82 t hm-2。截至2012年, 两系杂交水稻累计推广3.33× 107 hm2, 增产稻谷1.11× 1010 kg[3, 4, 5]。而在我国北方, 两系杂交水稻受气候环境条件的限制发展较慢。籼粳杂交F1具有大穗、大粒、生育期偏长和生长量大等特点, 较适宜在温光条件较好的南方稻区种植; 但在北方稻区生育期偏长和抗寒性差却是籼粳杂交F1最大的种植障碍, 易引起结实率低、早衰和倒伏等弊病, 使高产稳产受到极大的影响。所以, 在提高籼粳交强优势利用的同时, 避免籼稻血缘引入造成的适应性下降已成为困扰北方两系杂交水稻育种的重要问题。为此, 探讨籼粳交的遗传基础, 研究调节搭配亲本组合及其籼粳成分, 使北方两系杂交水稻能获得高产稳产成为了现阶段研究的重点。本文通过籼粳重组自交系(RIL)与粳型光温敏核不育系杂交获得杂种F1, 利用程氏指数法和分子标记法分析RIL和不育系的籼粳成分及遗传距离, 研究其与F1杂种优势的关系, 旨在为北方两系杂交水稻的发展打下理论基础。
2012年以粳型光温敏核不育系GB028S和高代籼粳交重组自交系(秋光× 七山占) RIL F10为亲本, 杂交构建F1群体, 该杂种群体F1共98个。2013年将杂种F1群体与RIL种植于辽宁省盐碱地利用研究所试验地, 每株系2行, 每行2 m, 行距为30.0 cm, 株距为13.3 cm, 每穴单株, 2次重复, 栽培管理同当地生产田。
杂种F1群体与RIL齐穗后测量株高, 成熟后收获每株系5株, 网室风干后考种, 调查有效穗数、穗长、穗粒数、千粒重、结实率及单株产量等性状。利用中亲优势代表杂种优势, HMP = (F1-MP)/MP× 100, 式中HMP为中亲优势, MP为中亲值。
调查分析不育系GB028S和籼粳交重组自交系的抽穗时壳色、叶毛性状、1~2穗节长、籽粒长宽比、稃毛、酚反应, 计算程氏指数(Cheng’ s index, Chi)。程氏指数在划分籼粳成分上分4级, 0~8为籼、9~13为偏籼、14~17为偏粳、18~24为粳。
均匀选取142个SSR引物用于标记亲本的基因型, 参照毛艇等[6]的方法鉴定籼粳成分, 将与秋光带型相同的带视为一个粳性位点, 以B来表示, 与七山占带型相同的带视为一个籼性位点, 以A来表示。基因型差异程度用粳性位点所占比例Dj值表示, 即每一株系的Dj值(%) = B位点数/(A位点数+B位点数)× 100。按Dj值的含义, 0~25%为籼型、25%~50%为偏籼型、50%~75%为偏粳型、75%~100%为粳型。按照Nei计算标准遗传距离[7], 亲本间的遗传距离GD (genetic distance) = -ln [2Mxy/(Mx+My)], 公式中Mx和My分别为X和Y两材料的总片段数, Mxy为两材料的共片段数。采用SPSS 19.0和Microsoft Excel 2013软件统计分析试验所得数据, 并绘制图表。
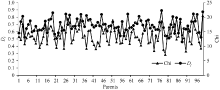
图1的RIL和GB028S程氏指数整体偏粳分布, 程氏指数主要分布在14~18之间, 其中GB028S的程氏指数为20.5。偏粳系数主要集中在0.60~0.90之间, 整体偏粳, GB028S的偏粳系数为0.875。程氏指数Chi和分子标记的偏粳系数Dj值呈极显著正相关(相关系数r = 0.450), 由此可见利用程氏指数法及分子标记法的偏粳系数在衡量群体籼粳成分上具有较高的相似性, 可以利用它们协同划分目标群体的籼粳成分。
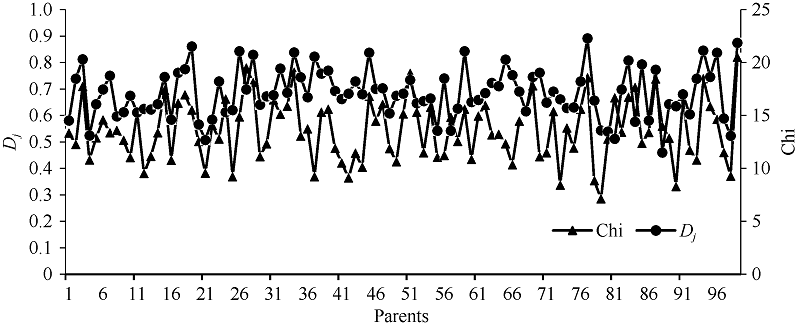 | 图1 RIL和GB028S的偏粳系数及程氏指数比较Fig. 1 Comparison of parental Chi and Dj |
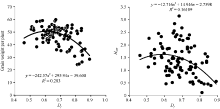
由图2可以看出单株产量与偏粳系数存在二次曲线关系, 父本RIL的偏粳系数Dj值在0.55~0.70区间时, F1的单株产量较高, Dj值为0.67时产量达峰值。单株产量杂种优势与偏粳系数也存在二次曲线关系(图2), F1的产量杂种优势的高峰区出现在父本RIL Dj值0.50~0.65区间内。两个高峰区在偏粳系数0.55~0.65重合, 说明就母本GB028S, 当父本偏粳系数为0.55~0.65时有形成较高产量及杂种优势的潜力。父本RIL的程氏指数和Dj值都与杂种F1的每穗粒数呈极显著负相关(表1), 父本的程氏指数与杂种F1的千粒重呈极显著正相关, 与结实率呈显著正相关, 与着粒密度呈极显著负相关。父本RIL的Dj值与杂种F1的单株产量呈极显著负相关。
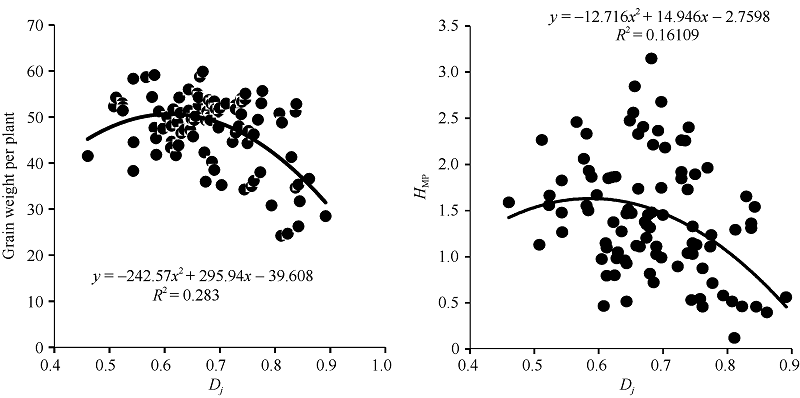 | 图2 RIL的Dj与F1单株产量和杂种优势的相关分析Dj: 偏粳系数; HMP: 中亲优势。Fig. 2 Relationship of RILs’ Dj with yield and HMP in F1Dj: Deviation japonica coefficient; HMP: Mid-parent heterosis. |
| 表1 RIL籼粳成分及亲本间遗传距离与F1产量性状和杂种优势的相关分析 Table 1 Relationship of RILs’ indica-japonica index and parental genetic distance with traits and heterosis in F1 |
由表1可以看出, F1产量的杂种优势与父本RIL的偏粳系数Dj值的负相关达到显著水平。F1穗数的杂种优势与父本RIL的程氏指数呈极显著负相关。F1结实率的杂种优势与父本RIL的偏粳系数Dj值呈显著负相关。F1其他农艺性状的杂种优势与父本RIL的程氏指数和Dj偏粳系数的相关均未达到显著水平。亲本间的遗传距离与F1单株产量和部分性状及其杂种优势的相关都未达到显著水平。
根据分子标记所在染色体不同, 分别计算父本RIL各家系每条染色体偏粳系数, 分析父本RIL单一染色体籼粳片段分布与杂种F1产量等性状及杂种优势的关系(表2)。杂种F1的产量与父本RIL的Chr.8和Chr.11的偏粳系数Dj值呈显著负相关, 在产量构成因素中, F1的穗数与父本RIL的Chr.8和Chr.11的Dj值呈显著负相关; F1的千粒重与父本RIL的Chr.1、Chr.11和Chr.12的Dj值的负相关, 及与Chr.9的Dj值的正相关均达到显著或极显著水平, 可见控制千粒重表达的关键籼粳成分基因多分布于Chr.1、Chr.9、Chr.11和Chr.12, 但作用方向各有不同; F1的结实率与父本Chr.3的Dj值呈极显著正相关, 与Chr.4和Chr.12的Dj值呈显著正相关; F1的株高与父本Chr.3和Chr.5的Dj值呈显著负相关, 与Chr.12的Dj值呈极显著负相关, 杂种F1的着粒密度与父本RIL的Chr.1和Chr.11的Dj值呈极显著负相关。
| 表2 RIL染色体偏粳系数与F1单株产量及部分性状的相关系数 Table 2 Correlation coefficients between chromosomes’ Dj and F1 traits |
表3表明, F1产量的杂种优势与父本Chr.12的偏粳系数Dj值呈极显著负相关。F1穗数的杂种优势与父本Chr.2的Dj值呈显著负相关, F1穗粒数的杂种优势则与父本Chr.4的Dj值呈显著负相关, F1千粒重的杂种优势与父本Chr.2的Dj值的负相关, 及与Chr.4的Dj值的负相关都达到显著水平。F1着粒密度的杂种优势与父本Chr.1和Chr.6的Dj值呈极显著负相关。
| 表3 RIL染色体成分与F1杂种优势的相关系数 Table 3 Correlation coefficients between chromosomes’ Dj and HMP |
以12条染色体的籼粳成分为变量, 对98份F1单株产量进行主成分分析, 经分析提取5个主成分, 分别用X1、X2、X3、X4、X5表示(表4), 第1到第5主成分贡献率分别为17.95%、12.00%、11.11%、9.61%和8.51%, 共累积贡献率为59.20%。第1主成分X1中Chr.5和Chr.8载荷最大, 第2主成分X1中Chr.4载荷最大, 第3主成分X1中Chr.11载荷最大, 第4主成分X1中Chr.10载荷最大, 第5主成分X1中Chr.12载荷最大。从表4可以看出贡献率最高的第1主成分内染色体的载荷值比较接近, 贡献差异较小。
| 表4 RIL染色体籼粳成分主成分载荷矩阵 Table 4 Load matrix of principal component analysis on chromosome elements of RILs |
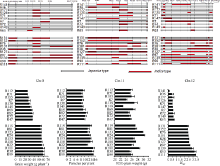
综合表2、表3和主成分分析可知, F1的产量及其构成要素和杂种优势与父本RIL Chr.8、Chr.11、Chr.12的偏粳系数Dj值关系密切。根据杂种F1的产量高低和产量杂种优势强弱, 挑选出极端高和极端低两组, 对其父本RIL的Chr.8、Chr.11和Chr.12染色体上的籼粳片段分布情况进行分析(图3)。如图所示, 当父本RIL染色体Chr.8上的分子标记RM408-RM25区间上存在籼性片段时, F1具有较高产量和穗数。当父本RIL染色体Chr.11上的分子标记RM286-RM4和PSM366-RM224之间存在籼性片段时, 杂种F1具有较高的产量、穗数和千粒重。当父本RIL染色体Chr.12上的RM519-RM270区间存在籼性片段时, 杂种F1具有较高的产量杂种优势。
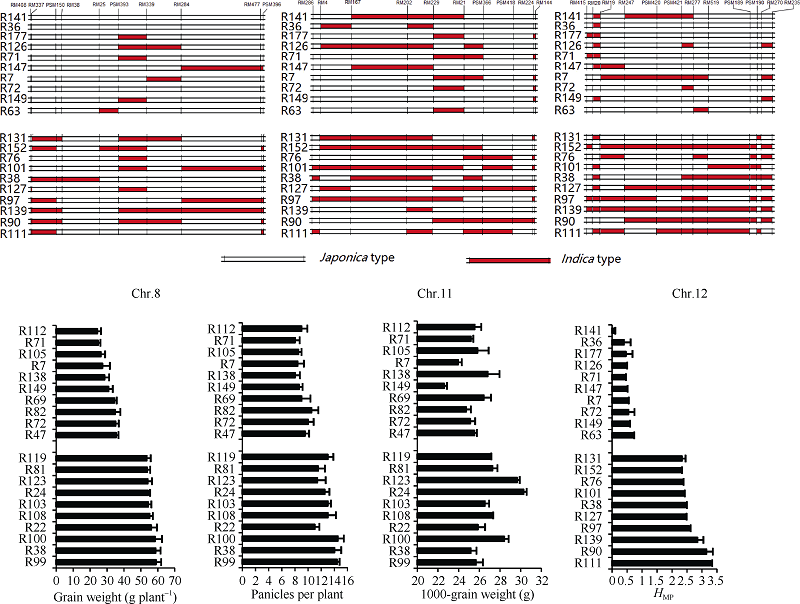 | 图3 RIL染色体籼粳成分与杂种F1产量及杂种优势相关的标记位点分布Fig. 3 Markers position for correlation of chromosomes’ Dj with yield and HMP in F1 |
程侃生的形态指数法(程氏指数法)在籼粳分类上得到众多学者广泛的认可, 能较圆满地区分籼粳品种, 但无法得知籼粳交育成品种的籼粳成分比例和构成。随分子生物学的飞速发展, 利用分子标记来研究亲本血缘关系及判定籼粳分类成为可能。本研究表明, 利用以形态指标的程氏指数及以分子标记为主的偏粳系数(Dj值)在分类群体籼粳成分上具有一定的相似性, 可作为划分目标材料籼粳成分的依据, 这与陈跃进等[8]的研究结果较一致。卢宝荣等[9]研究认为基于高通量InDel分子标记鉴定籼粳稻与基于程氏指数法鉴定结果在很大程度上表现一致。刘丹等[10]、孙健等[11]研究认为利用InDel、SSILP标记鉴定北方粳稻籼粳成分高效准确, 与程氏指数法鉴定结果较一致。而张培江等[12]、王彦荣等[13]研究认为程氏指数法与SSR分子标记对籼粳分类结果不完全一致。分析认为程氏指数法主要利用形态及部分生理特性, 受环境及人为操作影响较大, 有一定的误差。分子标记能直接反应DNA的差异, 相对准确率较高, SSR、ILP、InDel和SNPs[14, 15, 16, 17]等分子标记被认为是鉴定籼粳成分快速、高效的方法。
在父本籼粳成分构成与F1产量关系的分析中, 杂种F1单株产量及杂种优势与父本的程氏指数的相关分析不显著, 却与父本的偏粳系数呈极显著负相关, 进一步分析表明二者存在二次曲线关系, 说明基于粳型光温敏核不育系GB028S (Chi: 20.5, Dj: 0.875)组配的杂交组合, 当父本偏粳系数在0.55~ 0.65范围时具有形成杂种优势实现高产的潜力。在染色体的籼粳成分与单株产量等性状及相应杂种优势的相关分析中, F1单株产量与父本Chr.8和Chr.11的偏粳系数呈显著负相关, 当父本Chr.8上的RM408-RM337、RM337-PSM150、PSM150-RM38、RM38-RM25和Chr.11上的RM286-RM4、PSM366- PSM418、PSM418-RM224标记之间存在不同程度籼性片段时, 组配后的杂种F1具有较高的产量。很多研究也在上述区间附近发现很多调控产量及其相关的基因或QTL, Luo等[18]在研究杂交籼稻杂种优势上位性遗传基础时将穗数qPP8定位在RM38附近, 陈深广等[19]将控制结实率杂种优势的qHSF-8定位在RM337附近, 赵秀琴等[20]将穗粒数qGN8定位在RM38。陈冰嬬等[21]研究籼稻恢复系蜀恢527籽粒性状QTL时将千粒重qTGW11定位在RM286附近, Li等[22]和Chai等[23]将单株产量qGY11-2和qGYP11.7、赵芳明等[24]和王智权等[25]将千粒重qTGW11、徐建龙等[26]将单株有效穗数qPN11、赵秀琴等[20]将穗粒数qGN11b均定位在RM224附近, 可见标记RM224附近可能存在与产量相关的重要基因, 这需要在以后的研究中进一步筛选挖掘。单株产量的杂种优势与Chr.12的偏粳系数呈极显著负相关, 通过分析筛选出Chr.12上的RM519-PSM189、PSM189-PSM190和PSM190-RM270标记之间存在籼性片段时, F1具有较高的产量杂种优势。辛业云等[27]在研究两系杂交籼稻两优培九杂种优势中将产量qHY12定位在RM519附近, 王智权等[25]将每穗粒数qHSPP12和结实率qHSSR12及穆平等[28]将有效穗数qPN12定位在RM270附近。总之, 北方两系杂交粳稻利用籼粳交提高产量杂种优势, 其亲本选育上应侧重挖掘研究Chr.8、Chr.11和Chr.12在上述标记附近的籼性基因, 有效导入目标基因, 提高分子标记在育种实践上的应用价值。
亲本间的遗传差异被普遍认为是子代产生杂种优势的基础, 杂种优势的产生在一定范围内取决于双亲的遗传差异性和性状互补性等因素。而遗传距离作为定量描述遗传差异的方法之一, 被许多学者用于杂种优势的预测。本研究中亲本间的遗传距离与F1单株产量和部分性状及杂种优势的相关分析都未达显著水平, 表明分子标记遗传距离与F1的杂种优势没有明显和必然的联系, 这与李任华等[29]、廖伏明等[30]、孙传清等[31]、朱作峰等[32]和张涛等[33]研究结果比较一致。而蔡建等[34]和赵庆勇等[35]研究认为分子标记遗传距离与杂种优势呈显著正相关。王友林等[36]研究认为Indel标记遗传距离在不同类型品种间具有较好的杂种优势预测能力, 可以用于单穗重的优势预测, 即随着遗传距离的扩大, 杂种优势主要体现在单穗重的增加上。罗小金等[37]认为不同材料、不同遗传距离范围F1的单株产量及杂种优势与遗传距离的相关性存在很大差别, 表现出很复杂的关系, 在特定遗传距离范围内存在相关性。游书梅等[38]研究认为基于单一性状的QTL标记的遗传距离与杂种优势存在正相关。Melchinger等[39]研究认为杂种优势和遗传距离为二次曲线相关, 一定范围内杂种优势因亲本间遗传距离的增加而增强, 但超出这个范围, 杂种优势呈现减弱的现象。也有的研究认为, 同一杂种优势群内的亲本组合中, 亲本遗传距离与杂种一代呈显著相关; 而在不同杂种优势群之间较远的组合中, 相关不显著[40, 41, 42]。Saghai等[43]、Zhao等[44]也认为分子标记遗传距离与杂种优势的关系受材料来源、亲本数目、亲缘关系及所用标记位点的数量等因素的影响, 故而预测性不足。结合上述研究分析主要原因[45, 46, 47]: ①研究的性状及标记的数目较少, 有些性状对杂种优势的表现贡献较低或无贡献。②大多数量性状的表达易受环境影响, 环境(包括同一地点不同年份)的差异导致结果的不同, 使亲本间遗传差异估计的准确性大大降低。③由于上位性的作用, 增加标记的数目可能并不能增加上述相关性, 检测结果只能代表少量基因型, 因此所得遗传差异并不能全部代表亲本间的遗传差异, 而应该寻找与所研究性状有紧密连锁的标记位点。④性状之间相关的复杂性, 也导致遗传距离和杂种优势之间的复杂化。近年来, 洪德林等[48, 49, 50, 51, 52]率先在杂交粳稻上开展亲本产量性状优异配合力的标记基因型筛选工作, 研究得出RM152-165/170和RM2439-150标记是单株日产量和单株有效穗数优异配合力效应最大的标记基因型, RM8254-120/180标记对增加穗部性状的枝梗数贡献较大, 进而为提高粳稻杂种优势预测提供了参考。赵彦宏等[53]研究把杂种优势精确分担到各个QTL位点上, 估算控制性状的QTL 遗传效应进而预测杂种优势。
综上所述, 杂种优势的形成来源于亲本间的遗传差异, 而亲本间的遗传差异可用遗传距离来衡量。虽然基于分子标记的遗传距离用于杂种优势预测在理论和实践上尚无定论, 但普遍认为遗传距离的深化研究是预测杂种优势的有效途径, 只是在度量遗传距离的方法上还有待探讨, 找到能准确有效地度量遗传距离的手段, 将提高遗传距离在杂种优势预测上的应用价值。同时, 挖掘筛选与目标性状杂种优势密切相关的功能基因座位, 利用高通量分子标记精确定位, 分析主效基因及微效基因的作用, 对目标性状开展杂种优势的预测, 提高品种选育的目标性。
利用程氏指数法及分子标记法的偏粳系数在群体籼粳分类上具有较高的一致性。父本RIL的偏粳系数Dj值与单株产量及其杂种优势存在二次曲线关系。当父本的偏粳系数为0.55~0.65时, 杂种F1单株产量和杂种优势潜力较大。父本Chr.8、Chr.11和Chr.12上部分区段的籼粳成分与杂种F1产量及杂种优势关系密切。双亲的遗传距离与F1产量和相关性状及杂种优势没有明显的关系。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|