


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
基于生物量的冬小麦越冬前植株地上部形态结构模型
[陈昱利1 , 杨平1 , 张文宇2 , 张伟欣2 , 诸叶平3 , 李世娟3 , 巩法江1 , 毕海滨1 , 岳霆1 , 曹宏鑫2, *  ]
]
]
|
|


第一作者联系方式: E-mail: luckydogcyl@163.com
越冬前植株地上部形态建成是冬小麦株型构建和可视化的重要基础。为定量分析冬小麦越冬前植株地上部形态结构参数与器官生物量的关系, 以济麦22、泰农18和鲁原502为材料, 于2013—2014和2014—2015年度开展品种和施氮试验。利用2013—2014年度越冬前植株地上部形态结构参数和器官生物量等数据构建了冬小麦越冬前植株地上部形态结构模型。经2014—2015年度数据检验, 除叶鞘长和叶弦长模型精度略低外, 叶长、最大叶宽、叶切角和叶弦角模型精度均较高, 所建模型可较好地模拟不同品种与各施氮水平冬小麦越冬前植株地上部形态结构。
The aboveground morphogenesis is an important basis of plant morphological construction and visualization for winter wheat before overwintering. For quantitatively analyzing the relationship between the aboveground architectural parameters and organ biomass of winter wheat before overwintering, field experiments with different varieties (Jimai 22, Tainong 18, and Luyuan 502) and nitrogen levels were carried out in 2013-2014 and 2014-2015 wheat growth seasons. Simulation models for aboveground architectural of winter wheat before overwintering were built with the 2013-2014 dataset of aboveground architectural parameters before overwintering and organ biomass and validated by the 2014-2015 dataset, showing the models exhibited satisfactory predictions for leaf blade length, leaf maximum blade width, leaf blade tangent angle, and leaf blade bowstring angle, except for leaf sheath length and leaf bowstring length. The models built in this study are suitable to simulate the aboveground architecture of winter wheat varieties before overwintering under different nitrogen levels.
合理株型结构是小麦高产、稳产、优质的有效保证, 地上部形态结构是小麦生长发育过程与株型结构的全面展现, 而小麦生长模型与形态结构模型则是小麦生长发育过程与株型结构的定量表达。因此, 将小麦生长模型与形态模型有机结合, 建立基于生物量的小麦植株形态结构参数模型是功能-结构小麦模型研究的重要内容, 对小麦株型设计与栽培调控具有重要参考意义。
目前, 关于小麦形态结构模型的研究, 主要集中在小麦形态与环境因素的定量关系和小麦植株可视化方面。Evers等[1, 2]通过分析群体密度和遮阴对春小麦生长发育的影响, 构建了春小麦形态参数模型, 并对小麦发育的三维结构模型ADEL-wheat参数进行了量化; Fournier等[3]在ADEL-wheat模型基础上, 分析了冬、春小麦初始参数的差异。张文宇等[4, 5]利用系统分析方法和动态建模技术, 以生长度• 日为尺度, 构建了小麦主茎叶片及茎鞘夹角动态模拟模型; 并通过分析小麦主茎叶型和茎型指标与环境因素的定量关系, 模拟分层叶面积、叶向值等株型指标动态变化规律。陈国庆等[6]通过分析小麦茎鞘生长过程与环境因素的关系, 构建了小麦叶鞘和节间生长过程的动态模拟模型。Mabille等[7]构建了小麦籽粒形态参数模型。伍艳莲等[8]借助于OpenGL图形平台, 实现了小麦器官-个体-群体三层次的形态可视化; 谈峰等[9]在构建基于形态特征参数的小麦根系三维形态模型基础上, 实现了根系生长可视化。雷晓俊等[10]通过对小麦穗形态结构观测分析, 提出了基于形态特征参数的麦穗几何模型及可视化实现方法。上述研究中, 所建小麦形态模型大多未考虑同化物分配与小麦植株形态模型的定量关系, 缺乏冬小麦生长模型和形态模型的有效结合。
随着作物形态结构模型不断发展, 功能-结构小麦模型已成为小麦形态模型与可视化研究的一个重要方向[11]。目前, 基于生物量的作物功能-结构模型已有研究, 主要是通过确定作物器官生物量与形态参数的定量关系和作物形态参数之间的内在联系, 进而建立形态结构参数模型和实现植株可视化[12, 13, 14, 15, 16], 而基于生物量的小麦植株地上部形态结构参数模型报道较少。本文在前人研究基础上, 以器官生物量为尺度, 通过分析冬小麦越冬前地上部植株形态参数与器官生物量的定量关系, 将冬小麦生长模型和形态模型结合, 建立冬小麦越冬前地上部植株形态结构模型。为冬小麦生长模型与植株地上部形态结构模型结合, 进而为建立功能— 结构小麦模型奠定基础。
选取3个代表性小麦品种, 即济麦22 (株型紧凑, 山东省农业科学院作物研究所育成)、泰农18 (中间型, 泰安市泰山区瑞丰作物育种研究所、山东农业大学农学院育成)和鲁原502 (株型松散, 山东省农业科学院原子能农业应用研究所、中国农业科学院作物科学研究所育成)。
于2013年10月至2015年6月在山东省淄博市农业科学研究院试验农场开展品种与施氮试验, 设3个品种, 5个氮肥处理, 3次重复, 共计45个小区。土壤为褐土, 0~30 cm耕层含有机碳32.14g kg-1、全氮1.62 g kg-1、速效磷14.40 mg kg-1、速效钾150.32 mg kg-1、pH 8.16。采用裂区设计, 主区为品种(V), 即V1 (济麦22)、V2 (泰农18)和V3 (鲁原502), 副区为施氮水平(N)。小区面积10.0 m × 1.5 m = 15.0 m2, 行道宽0.5 m。基施有机肥15 000 kg hm-2、P2O5 112.5 kg hm-2、K2O 112.5 kg hm-2。其他管理同大田高产栽培管理。
2013年10月至2014年6月, 设N1 (0 kg hm-2)、N2 (84.375 kg hm-2)、N3 (168.75 kg hm-2)、N4 (253.125 kg hm-2)和N5 (337.5 kg hm-2) 5个施氮水平。50%作基肥, 在播种前施入; 50%作追肥, 在拔节期施入。2013年10月7日播种, 基本苗225万hm-2。2014年10月至2015年6月, 根据上年度试验结果, 为了更切合生产实际, 设N1 (0 kg hm-2)、N2 (75 kg hm-2)、N3 (150 kg hm-2)、N4 (225 kg hm-2)和N5 (300 kg hm-2) 5个施氮水平。50%作基肥, 在播种前施入; 50%作追肥, 在拔节期施入。2014年10月12日播种, 基本苗225万 hm-2。第1年试验数据用于建立模型, 第2年试验数据用于模型检验。
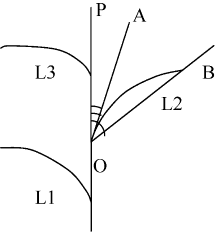
自三叶期标记叶位, 每隔10 d选取每处理长势一致的植株5株, 用直尺、游标卡尺等测定主茎叶片长度、叶片最大宽度、叶鞘长、叶弦长、茎叶夹角(叶切角、叶弦角)(图1)等形态指标。然后, 分不同叶位将叶片、叶鞘等分装, 105℃下杀青30 min, 再80℃烘至恒重后称重。
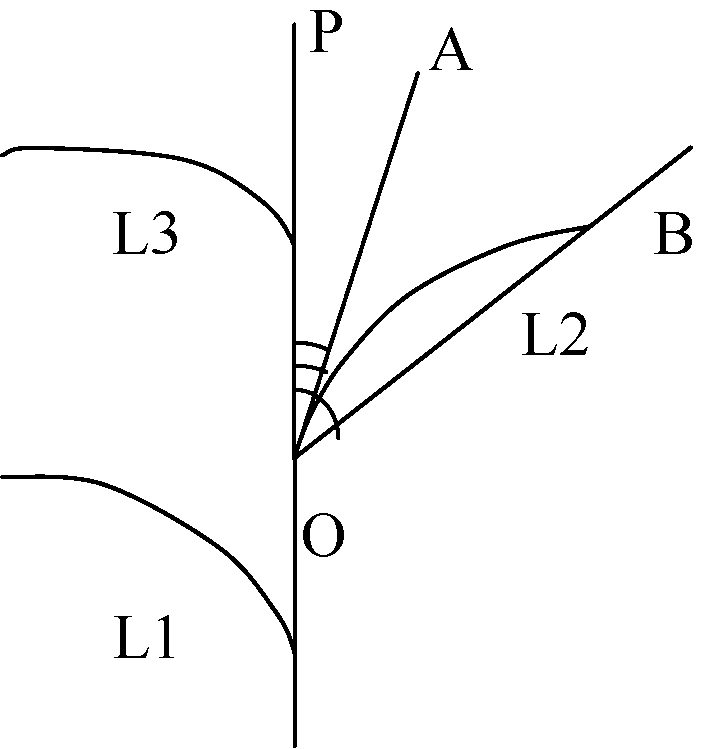 | 图1 叶片角度结构示意图OP代表主茎, L1、L2和L3代表主茎第一、第二和第三叶位叶片。Fig. 1 Schematic diagram of leaf blade anglesLine OP stands for the main stem and L1, L2, and L3 stand for the 1st, 2nd, and 3rd leaf, respectively. |
叶片长度为叶片伸直状态下自叶片基部至叶尖的直线距离; 叶片最大宽度为叶片的最大叶宽值, 一般在叶片中部; 叶弦长为叶片自然伸展状态下自叶片基部至叶尖的空间直线距离; 叶切角为叶片基部切线与主茎的夹角(AOP, 图1); 叶弦角为叶弦与主茎的夹角(BOP, 图1)。各参数均以3次测量取平均值。
采用Microsoft Excel 2007和Sigmaplot v10.0统计分析软件处理试验数据。
利用根均方差(RMSE)、平均绝对误差(da)和平均绝对误差与观察值平均数的比值(dap)统计量检验模型, 并绘制观测值与模拟值的1︰1关系图, 以检验模型。
 |
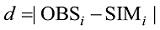 |
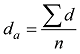 |
 |
式中, 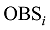

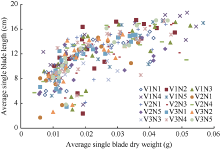
2.1.1 叶长模型 生物量作为小麦叶片生长发育的物质基础, 是小麦叶片形态形成的关键。2013— 2014年试验数据表明, 越冬前冬小麦不同叶位叶片长度随其生物量增加呈线性增长的趋势, 品种间有较大差异(图2)。根据叶片伸长与叶片生物量变化的关系, 主茎叶片长度可由如下公式计算。
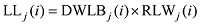 |
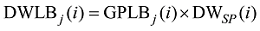 |
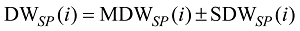 |
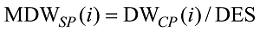 |
式中, j表示主茎叶位, i表示出苗后的天数, LLj(i)为出苗后第i天主茎第j叶位叶片长度(cm), DWLBj(i)为出苗后第i天主茎第j叶位叶片干生物量(g), RLWj(i)为出苗后第i天主茎第j叶位叶片长度和干生物量的比值(cm g-1), CPLBj(i)为出苗后第i天主茎第j叶位叶片干生物量与单株地上部干生物量的比值(g g-1), DWSP(i)为出苗后第i天单株地上部干生物量(g), MDWSP(i)为出苗后第i天单株地上部平均干生物量(g), SDWSP(i)为出苗后第i天单株地上干生物量标准误差(g), DWCP(i)为出苗后第i天单位面积植株地上部干生物量(g m-2), DES为种植密度(m-2)。
 | 图2 2013-2014年不同处理叶片长度随叶片干重的变化趋势Fig. 2 Changes in the average single blade length by the average single blade dry weight for different treatments in 2013-2014 |
随着叶位的增加, 越冬前冬小麦主茎不同叶位RLW逐渐变小, 呈指数函数变化趋势, 品种间无显著差异(图3-A); 而越冬前主茎不同叶位CPLB先变大后变小, 在第3和第4叶位达到最大, 呈现二次曲线变化趋势, 品种间无显著差异(图3-B)。不同叶位叶片RLW和CPLB可用如下公式表示。
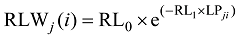 |
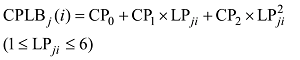 |
式中, RLWj(i)为出苗后第i天主茎第j叶位叶片长度和干生物量的比值(cm g-1); CPLBj(i)为出苗后第i天主茎第j叶位叶片干生物量与单株地上部干生物量的比值(g g-1); LPji为出苗后第i天主茎第j叶片的叶位; RL0、RL1、CP0、CP1、CP2为模型参数, 由试验数据获得(表1)。
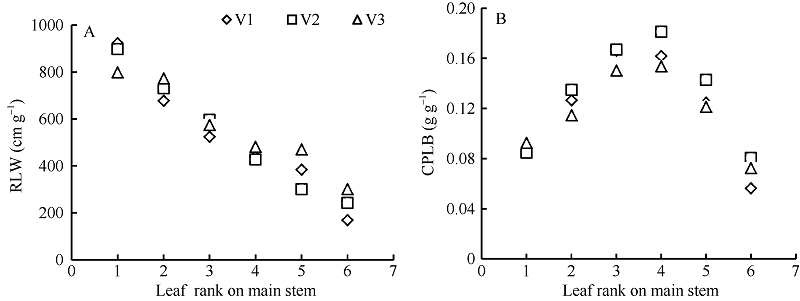 | 图3 叶片长度与地上部干生物量比值(A)和叶片干生物量与单株地上部干生物量比值(B)随叶位的变化趋势(2013-2014)V1、V2和V3分别表示品种济麦22、泰农18和鲁原502。Fig. 3 Changes in RLW(A) and CPLB (B) with the leaf rank on main stem (2013-2014)V1, V2, and V3 represent varieties Jimai 22, Tainong 18, and Luyuan 502, respectively. RLW: ratio of blade length to leaf dry biomass; CPLB: ratio of leaf dry biomass to shoot dry biomass. |
| 表1 各模型参数值及其统计检验 Table 1 Parameters of various models and their statistical test |
2.1.2 最大叶宽、叶鞘长和叶弦长模型 越冬前冬小麦主茎不同叶位叶片最大叶宽、叶鞘长和叶弦长均随该叶位叶片伸长呈一次函数的增长趋势, 品种间无显著差异(图4)。
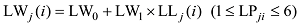 |
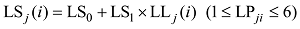 |
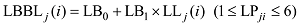 |
式中, LWj(i)、LSj(i)和LBBLj(i)分别为出苗后第i天主茎第j叶片的最大叶宽(cm)、叶鞘长(cm)和叶弦长(cm); LLj(i)为出苗后第i天主茎第j叶位叶片长度(cm); LPji为出苗后第i天主茎第j叶片的叶位; LW0、LW1、LS0、LS1、LB0、LB1为模型参数(表1)。
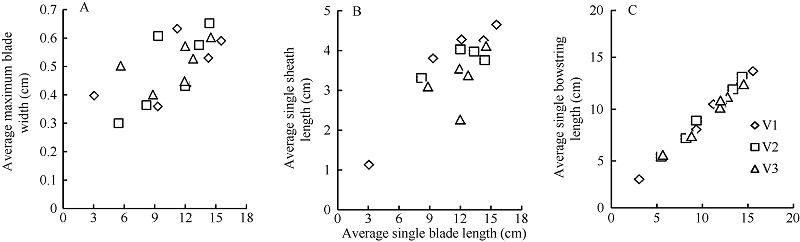 | 图4 最大叶宽(A)、叶鞘长(B)和叶弦长(C)随叶长的变化趋势(2013-2014)V1、V2和V3分别表示品种济麦22、泰农18和鲁原502。Fig. 4 Changes in the maximum single blade width (A), single sheath length (B), and single bowstring length (C) with the leaf blade length (2013-2014)V1, V2, and V3 represent varieties Jimai 22, Tainong 18, and Luyuan 502, respectively. |
2.1.3 叶片角度模型 叶片生物量是叶片角度形成的重要生理学和物理学基础[10], 因此将某叶位叶片叶切角和叶弦角引入模型, 并计算该叶位叶片干生物量与叶切角和叶弦角的比值RTWj(i)和RBWj(i)。越冬前冬小麦主茎不同叶位叶片的RTWj(i)和RBWj(i)随叶位的增加而降低, 呈指数函数变化趋势(图5)。
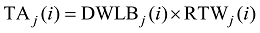 |
 |
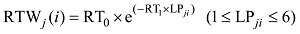 |
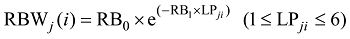 |
式中, TAj(i)和BAj(i)分别为第j叶第i天的叶切角(° )和叶弦角(° ), DWLBj(i)为出苗后第i天第j叶位叶片干生物量(g), RTWj(i)和RBWj(i)分别为第i天第j叶的叶切角和叶弦角与叶片干物重的比值(° g-1), LPji为出苗后第i天主茎第j叶片的叶位, RT0、RT1、RB0、RB1为模型参数(表1)。
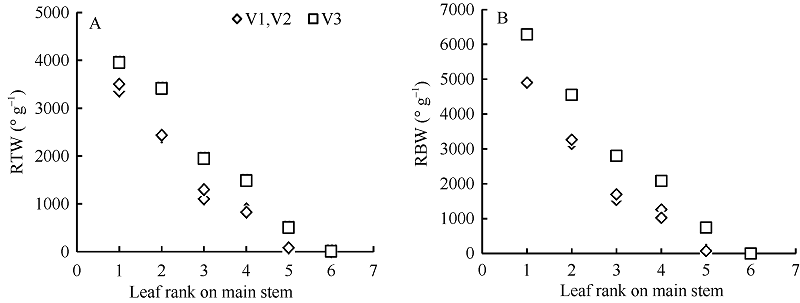 | 图5 叶片叶切角与叶片干生物量比值(A)和叶弦角与叶片干生物量比值(B)随叶位的变化趋势(2013-2014)V1、V2和V3分别表示品种济麦22、泰农18和鲁原502。Fig. 5 Changes in RTW (A) and RBW (B)with the leaf rank on main stem (2013-2014)V1, V2, and V3 represent varieties Jimai 22, Tainong 18, and Luyuan 502, respectively. RTW: ratio of blade tangent angle to leaf dry biomass; RBW: ratio ofblade bowstring angle to leaf dry biomass. |
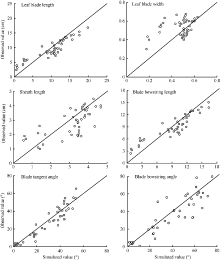
采用独立试验资料对所建冬小麦越冬前植株形态参数模型进行检验, 结果冬小麦越冬前植株地上部主茎形态结构参数叶长、最大叶宽、叶鞘长、叶弦长、叶切角和叶弦角的实测值与模拟值的吻合程度较好(表2和图6), 各模型r值在0.708~0.998之间(P < 0.001)。叶长、最大叶宽、叶弦长以及叶切角模型的dap值在10%~20%之间, 表明模型精度较好, 而叶鞘长及叶弦角模型精度较低(dap> 20%), 表明模型有改进空间。
| 表2 冬小麦越冬前植株地上部形态结构模型观察值与模拟值比较的统计参数 Table 2 Comparison of statistical parameters of simulation and observation in winter wheat aboveground architectural parameter models before overwintering |
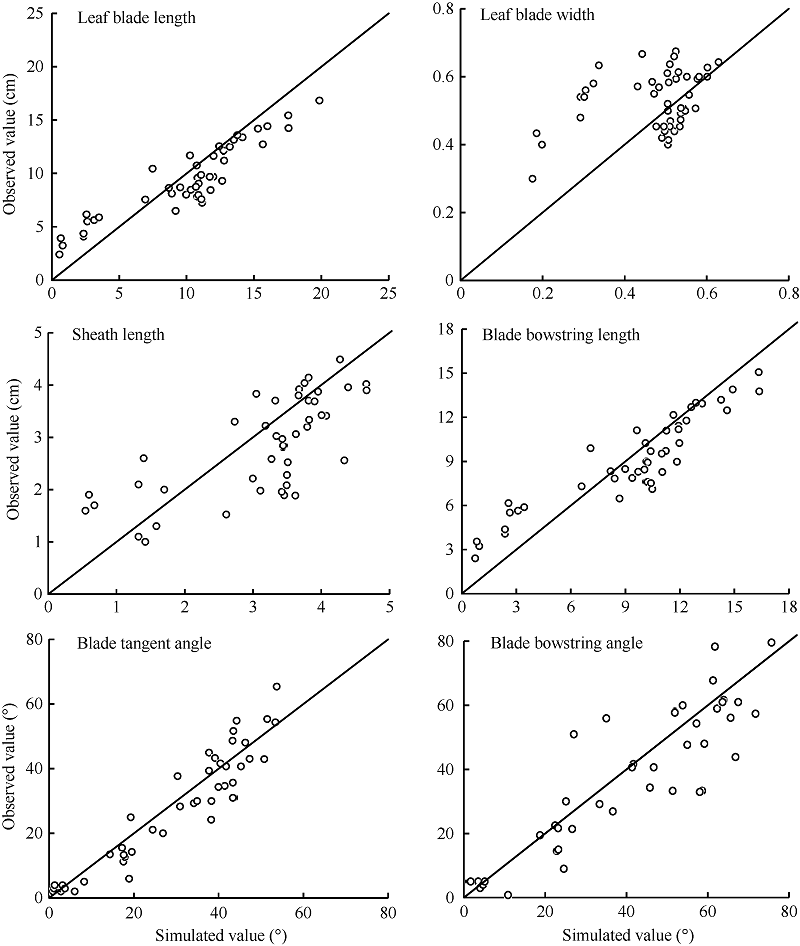 | 图6 实测值与模拟值比较(2014-2015)Fig. 6 Comparison between observed and simulated values (2014-2015) |
随着作物形态模型不断发展, 功能— 结构植物模型(functional-structural plant model, FSPM)已经成为作物形态模型与可视化研究的一个重要方向[17]。FSPM是作物生长模型之功能与作物形态模型之结构的有机结合, 通过将作物生理过程描述与3D显示相结合, 不断强化对作物生理机制、品种基因型与环境响应研究[11]。本研究通过分析小麦地上部植株形态参数与生物量的定量关系, 将小麦生长模型和形态模型结合, 建立了基于生物量的小麦植株地上部植株形态结构参数模型。生物量形成受小麦品种和环境因素(光、温度、水分、CO2、施肥等)综合影响, 而生物量可通过小麦生长模型估算并响应品种和环境因素差异[18]。目前, 针对小麦形态结构参数与环境因素的定量关系以及植株可视化已开展了较多形态结构模型方面的研究[19, 20, 21], 例如, 谭子辉[19]通过分析小麦叶、茎、蘖、穗等形态建成与其生理发育时间的关系, 初步研究了小麦植株形态建的模拟模型。然而, 现有形态结构模型缺少与生长模型的结合, 而生物量作为小麦生长发育的一个重要指标, 对小麦植株形态建成具有重要作用, 是小麦生长模型和形态结构模型有机结合的桥梁。除生物量外, 本研究中在模型中引入了RLW、CPLB、RTW、RBW等参数, 均具有一定生物学意义。
本研究表明, 冬小麦叶片长度和生物量存在线性关系, 这与曹宏鑫等[12]在水稻和张伟欣等[18]在油菜上的研究结果相似。叶片角度是小麦株型结构的一个重要指标, 随着分蘖的发生, 叶鞘与主茎之间也会形成一定角度[3], 使叶片角度发生变化, 故在叶片角度(叶切角和叶弦角)的模拟上存在一定误差。由于在越冬前, 小麦植株分蘖较小, 本文只涉及小麦单株主茎形态结构, 所建模型是否适合小麦分蘖, 还有待检验; 小麦植株形态结构除了受品种、氮素等因素影响外, 还受水分[22]、播期[23]、密度[4]等多种因素影响。特别是播期和种植密度对小麦形态结构有较大影响[24], 下一步拟将把播期和密度作为研究重点, 在不同种植密度下, 确定小麦不同叶位叶片干物质积累与分配的变化规律并建立相应的形态结构参数模型。此外, 叶形和叶曲线模型也是小麦叶片形态结构模型研究的重要内容[25, 26, 27, 28, 29], 本文尚未涉及。
冬小麦越冬前地上部植株形态结构指标主要包括数量指标和质量指标2个部分。本研究只考虑了冬小麦越冬前地上部植株形态数量指标与生物量的定量关系, 并未涉及冬小麦越冬前地上部植株形态质量指标(叶片含氮量、含水量、叶片厚度等)的差异。
在研究冬小麦植株地上部器官生物量分配的基础上, 通过分析冬小麦越冬前植株地上部各形态参数指标与生物量的定量关系及其各形态参数之间的内在联系, 建立了基于生物量的冬小麦越冬前植株地上部各形态参数模型。除叶鞘长和叶弦角模型精度较低外, 所建其他形态结构参数模型准确性均较高, 具有一定的机理性和普适性。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|