






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
亚洲棉bZIP蛋白家族的鉴定及 GaFD基因的组织表达分析
[张彦楠, 蔡大润, 黄先忠*  ]
]
]
|
|


第一作者联系方式: E-mail: zhangyn0513@163.com, Tel: 0993-2057262
碱性亮氨酸拉链(basic leucine zipper, bZIP)是真核生物中数量最多并且最具多样性的转录因子之一, 参与植物生长发育及响应生物和非生物胁迫。本研究利用亚洲棉( Gossypium arboreum)全基因组数据库, 通过生物信息学分析分布在13条染色体上的159个 bZIP家族基因的全序列。系统进化、基因结构和保守基序分析表明这些基因分成13个亚家族。其中, A亚家族有3个 GaFD基因 GaFD1、 GaFD2和 GaFD3, 通过实时荧光定量PCR分析3个 GaFD基因在不同组织中的表达, 结果表明 GaFD1和 GaFD2在SAM中的表达量最高, GaFD3在茎中表达量最高。研究表明棉花基因组中具有数量众多的bZIP家族成员, 不同基因结构及 FD基因不同的表达特征表明bZIP基因在棉花生长发育中可能具有不同的功能, 这些结果为进一步解析棉花bZIP家族基因的功能和作用机理积累了有价值的资料。
The basic leucine zipper (bZIP) is one of the largest and most diverse transcription factors in eukaryotes, and is involved in various processes of plant growth and development and in response to biotic and abiotic stresses. In this study, 159 bZIP family genes were identified and their complete gene sequences were obtained by using bioinformatics analysis method, based on Gossypium arboreumwhole genome database, and 159 bZIP genes were loaded on 13 chromosomes. The 159 bZIP genes were categorized into 13 groups based on their phylogenetic relationships, gene structures and conserved motifs. In addition, three GaFD homologous genes GaFD1, GaFD2, and GaFD3 were identified, which belongs to A subfamily in G. arboretum. The expression patterns of GaFD genes in different tissues were determined by using quantitative Real-time reverse transcription PCR (qRT-PCR) method. The results showed that GaFD1and GhFD2were preferentially expressed in the shoot apical meristem (SAM), whereas GaFD3was preferentially expressed in stem. The results revealed that a number of bZIP family members exist in cotton genome, and FD genes with different structures and expression patterns play different roles in the development of cotton, which provides valuable information for dissecting the function and mechanism of bZIPs in cotton.
在生物体中, 转录因子(TF)负责调控基因的表达, 碱性亮氨酸拉链(bZIP)转录因子是最大并且最具多样性的转录因子家族之一。bZIP转录因子根据它们共有的bZIP保守结构域而被命名。bZIP结构域由一个碱性区域和一个亮氨酸拉链区域组成, 包含60~80个氨基酸, 其中碱性区域具有一个含18个氨基酸残基的保守基序N-x7-R/K-x9, 这个区域的作用是参与细胞核定位和DNA结合; 亮氨酸拉链结构域的特征是每7个氨基酸的第7位含有一个亮氨酸(Leu), 或者还含有其他疏水性氨基酸, 比如异亮氨酸(Ile)、缬氨酸(Val)、苯丙氨酸(Phe)和甲硫氨酸(Met), 该结构域参与bZIP蛋白与DNA结合之前的二聚体化[1]。植物bZIP蛋白能与含有AGCT核心区域的DNA序列元件特异性结合, 并且优先与G-box (CACGTG)、C-box (GACGTC)和A-box (TACGTA)结合[2]。当bZIP蛋白与DNA序列相互作用时, 其碱性结构域的N端插入DNA双链的大沟, 同时亮氨酸拉链的C端二聚体化而形成一个叠加的卷曲螺旋[3, 4]。随着更多真核生物基因组测序的完成, 越来越多物种的bZIP转录因子家族已被鉴定或者预测[5, 6, 7, 8, 9, 10, 11, 12]。但在植物中只有很少一部分bZIP转录因子的功能被确定。已有的研究表明植物bZIP蛋白参与许多器官和组织的分化、种子萌发[13]、花序及叶片的发育[14]; 另一方面, bZIP蛋白还参与信号传递和响应生物、非生物刺激, 比如脱落酸(ABA)信号、渗透作用、组织缺氧、干旱、高盐和冷胁迫[15], 以及病原体防御等[16]; bZIP蛋白也响应光效应, 并且参与光形态建成[17]。
根据碱性结构域和其它保守基序的特征, 将拟南芥的75个bZIPs基因划分为A、B、C、D、E、F、G、H、I和S类10个亚家族[9]。其中A亚族中有少部分bZIPs基因已被克隆, 如ABF和AREB主要在ABA和胁迫信号调控网络中发挥重要作用[15, 18]; ABI5主要参与种子萌发和早期幼苗的发育[13, 19]。A亚家族中的FD基因对植物侧枝发育、花序形成和开花起到重要作用[14, 20]。拟南芥开花位点T控制基因FT编码成花素(Florigen)蛋白, 成花素在叶片中合成, 经过韧皮部到达顶端分生组织(SAM), 与在SAM产生的FD蛋白结合, 促进下游开花身份相关基因如AP1等表达, 进而促进开花[20]; 水稻Hd3a编码的蛋白是水稻中的成花激素, 在水稻SAM中, Hd3a、成花素受体蛋白14-3-3与OsFD1相互作用, 形成一个成花素激活复合物FAC来促进水稻开花[21], 而Hd3a、14-3-3和OsFD2蛋白形成一个FAC来调节叶片的发育[14]。在玉米C亚家族中, O2通过和PBF蛋白相互作用调控胚乳发育[22]。D亚族参与防御病害和生理生长两种不同进程[23, 24, 25, 26, 27]。G亚家族的成员参与光应答启动子的调节[28]和种子的成熟[29]。H亚家族HY5参与光形态发生和光信号转导, 进而影响下胚轴和根的发育[18]。I亚家族中, 烟草RSG和番茄VSF-1基因调节植物的生长发育[30, 31]。S亚家族是拟南芥bZIP家族中成员最多的, 但是只有ATB2基因功能研究的报道, 表明ATB2与平衡碳水化合物的供需有关[32], 此外, 一些S亚家族的同源基因在胁迫后能激活转录和在花的特定部位特异性表达[33, 34]。
异源四倍体陆地棉(Gossypium hirsutum)占全世界栽培种棉花的90%以上, 是可再生的纺织用纤维的主要来源, 还能产生油料种子[35]。陆地棉的2个现存的祖亲属亚洲棉(Gossypium arboreum)和雷蒙德氏棉(Gossypium raimondii)的基因组测序的完成[36, 37], 为二倍体棉花bZIP转录因子家族的分析提供了数据支持, 并且随着异源四倍体陆地棉[38, 39]、海岛棉基因组的测序[40], 进一步加快了棉花生物技术的进步。bZIP蛋白在棉花整个发育过程中起重要的作用, 尤其是bZIP家族的FD基因, 与棉花花器官的发育和开花有关。本研究利用二倍体棉花基因组数据库鉴定亚洲棉bZIP家族成员, 并查找bZIP家族FD基因序列, 分析FD基因在亚洲棉不同组织中的表达特征, 为进一步研究棉花bZIP家族基因的功能奠定了基础。
亚洲棉(G. arboreum)中亚1号A由国家种质三亚野生苗圃提供。2015年4月26日将其种植于石河子大学试验农场。播种后40 d采集根、茎、真叶, 在解剖镜下剥去叶片将SAM分离出来; 采集开花当天的花、雌蕊和雄蕊以及20 d的棉铃, 将胚珠从棉桃中剥离。所有棉花组织立即浸没于液氮中, -80℃保存备用。
通过NCBI的全基因组鸟枪(whole-genome shotgun contigs, WGS)数据库(http://www.ncbi.nlm. nih.gov/)获得亚洲棉(G. arboreum)全基因组序列, 分别从拟南芥转录因子数据库(DATF) (http://datf.cbi. pku.edu.cn/)和植物转录因子数据库v2.0 (Plant TFDB) (http://planttfdb.cbi.edu.cn/)获得拟南芥和雷蒙德氏棉bZIP转录因子序列。分别以拟南芥和雷蒙德氏棉的bZIP基因的开放阅读框序列(ORF)为查询序列(query), 运行TBLASTX, 选择数据库wgs、物种G. arboreum, 其他为缺省值, 搜索亚洲棉基因组上的bZIP基因序列, 选择比对结果中E值小于E-10的序列。通过MegAlign在线软件比对分析雷蒙德氏棉和获得的亚洲棉的bZIP基因序列, 获得亚洲棉bZIP基因的ORF序列。用SMART (http://smart. emblheidelberg.de/)和Pfam (http://pfam.sanger.ac.uk/)软件鉴定亚洲棉bZIP基因编码的蛋白序列是否存在bZIP结构, 参数设置为缺省值, 去除非全长的短片段以及相同基因的冗余序列, 保存完整且具有表达信息的基因序列用于下一步分析, 最终确定亚洲棉基因组中bZIP基因的序列。
利用ClustalX2程序比对159个亚洲棉、13个拟南芥[9](https://www.arabidopsis.org/)和11个水稻[10](http://rice.plantbiology.msu.edu/)的bZIPs蛋白的序列, 通过MEGA5.1软件分析系统发育和分子进化, 以邻位相连法(Neighbor-Joining, NJ)构建进化树[41], 运用p-distance模型, BootStrap参数设置为1000次重复, 使分支结果更为可靠。
利用在线MEME分析工具(MEME version 4.8.1) (http://meme.sdsc.edu/meme/cgi-bin/meme.cgi)鉴定亚洲棉bZIP转录因子除bZIP结构以外的保守基序, 设置基序的最小宽度、最大宽度和最大数量, 分别限制为6、200和25。由最低E-value < E-48值确定基序的最终数量, 根据这些基序在MEME中的位置顺序和出现的频率确定亚洲棉bZIP家族序列所属的亚家族。
利用在线软件Gene Structure Display Server (GSDS) (http://gsds.cbi.pku.edu.cn/)分析亚洲棉bZIP基因外显子/内含子(intron-exon)的分布和内含子的剪接位点。
以拟南芥的1个bZIPA亚家族基因AtFD (GenBank登录号为AT4G35900)的ORF序列为query, 运行TBLASTX, 选择数据库wgs、物种G. arboreum, 搜索亚洲棉FD基因, 选择比对结果中E值小于E-10的序列。利用NCBI的BlastN工具(参数设置为缺省值)比对分析GaFD基因与GenBank数据库中其他物种FD基因的相似性, 同时运用Clustal X2程序比对GaFD和其他物种FD蛋白的氨基酸序列, 并且通过进化分析和氨基酸序列的相似性将GaFD基因命名。
采用杭州博日科技有限公司(BIOER) Biospin多糖多酚植物总RNA提取试剂盒, 参照其说明书, 提取亚洲棉不同组织的总RNA; 利用北京百泰克公司的Supermo III M-MLV反转录酶, 参照试剂盒说明书, 合成cDNA第1链。
根据上述分析得到GaFD基因的序列, 设计实时荧光定量qRT-PCR分析的引物(表1), 内参基因为棉花的Ubiquitin7。以亚洲棉不同组织的cDNA为模板, 采用北京康为世纪有限公司FASTSYBR Mixture (With ROX)试剂盒, 利用7500 Fast实时荧光定量PCR仪(Life Technologies, Foster City, CA, USA)检测基因表达量。检测每份样品目的基因和内参基因的CT值(循环阈值), 每份样品3次重复, 并且进行3次独立的实验。
| 表1 本研究所用的引物 Table 1 PCR primers used in this study |
qRT-PCR总体系含cDNA 30 ng、2 × FASTSYBR混合物5.0 μ L、基因特异正向引物(10 μ mol L-1) 0.2 μ L、反向引物(10 μ mol L-1) 0.2 μ L, 用RNase-Free H2O补足到10.0 μ L。采用二步法PCR, 反应程序为95℃预热10 min, 95℃ 15 s, 60℃ 1 min, 共40个循环, 熔解曲线阶段为仪器默认程序。采用2-Δ CT法分析试验数据, 先分别计算出每组的Δ CT = CT目的基因- CT内参基因, 再根据Δ CT值求出2-Δ CT及标准误, 使用Microsoft Excel 2010软件处理数据。
通过比对分析亚洲棉基因组数据库, 共获得159个GabZIPs基因, 它们分布于94个基因组鸟枪序列(shotgun contigs)上, 被命名为GabZIP1~GabZIP94, 由相同shotgun contigs编码的不同的bZIP蛋白共享一个基因名(用额外的小数部分区分), 比如GabZIP1.1~GabZIP1.3。159个GabZIPs基因分布于13条染色体上, 其中第1染色体上有6条序列, 第2染色体上有6条序列, 第3染色体上有12条序列, 第4染色体上有11条序列, 第5染色体上有19条序列, 第6染色体上有13条序列, 第7染色体上有9条序列, 第8染色体上有18条序列, 第9染色体上有11条序列, 第10染色体上有23条序列, 第11染色体上有13条序列, 第12染色体上有6条序列, 第13染色体上有8条序列; Gabzip28、Gabzip49.1、Gabzip49.2和Gabzip75这4条序列的染色体定位情况未知。
由进化分析可以看出(图1), 除GabZIP64外, 其余182条bZIP序列分为A、B、C、D、E、F、G、H、I、S、U、V、W 13个亚家族, 其中A~S 10个亚家族同拟南芥和水稻bZIP家族分类一致。AtbZIP62、AtbZIP72和AtbZIP60在拟南芥bZIP家族不属于任何亚家族, GabZIP74.1和GabZIP74.2与AtbZIP62和OsbZIP80蛋白划分为同一个进化分支, 把它们归为一个新的亚家族, 命名为U亚家族; GabZIP12和GabZIP66与AtbZIP72蛋白划分为同一个进化分支, 归为一个新的亚家族, 命名为V亚家族; GabZIP19.1、GabZIP19.2、GabZIP19.3和GabZIP83与AtbZIP60蛋白划分为同一个进化分支, 归为一个新的亚家族, 命名为W亚家族。
 | 图1 亚洲棉bZIP家族蛋白进化分析Fig.1 Phylogenetic analysis of the GabZIP family proteins in G. arboretum 利用MEGA5.1软件及邻位相连法NJ构建进化树, BootStrap参数设置为1000次重复, 有13个进化分支被命名为A~I和S~W, 13种不同的颜色代表13个亚家族, 数字代表每个分支的成员。 The phylogenetic tree was generated using the neighbor-joining criteria in MEGA 5.1 with 1000 Bootstrap Replicates. The 13 distinct clades were named as A-I and S-W. The 13 different color boxes represent 13 clades. Number represents members in each clade. |
亚洲棉bZIP A亚家族是最大的一个进化分支, 包括38条GabZIPs蛋白序列。B亚家族和H亚家族的成员最少, 分别只有1条GabZIP蛋白序列。C亚家族有11个成员, D亚家族有29个成员, E亚家族有8个成员, F亚家族有5个成员, G亚家族有14个成员, I亚家族有17个成员, S亚家族有26个成员, U亚家族有2个成员, V亚家族有2个成员, W亚家族有4个成员。
亚洲棉bZIPs蛋白一共有25个保守基序(E-value < 1E-1.0), 包括保守的bZIP基序(motif 6), 不同亚家族bZIP蛋白的保守基序具有一定的分布规律(图2)。motif 8存在于除了D亚家族外的所有亚家族中, 但有些基序仅共存于个别亚家族中, 比如motif 1只存在于A、S亚族中, motif 4存在于D和E亚族中, motif 5存在于A和D亚族中, motif 7存在于A、C和S亚族中, motif 14存在于A和G亚族中, motif 17存在于E和I亚族中, motif 18存在于D、S和W亚族中, motif 22存在于D和I亚族中, motif 24存在于E、I和V亚家族中。此外, 还有一些保守基序只存在于特殊的亚家族中, 比如, motif 2、3、9和27只存在于A亚族中, motif 10、11、12、13、15和16只存在于D亚族中, motif 23只存在于F亚族中, motif 19只存在于G亚族中, motif 20和25只存在于I亚族中。这一现象表明每个亚家族中存在的特殊基序决定了该亚家族中成员的功能。
 | 图2 亚洲棉bZIP家族结构分析Fig.2 Structural characterization of GabZIP family in G. arboretum 25种不同的颜色代表25个不同的保守基序, 外显子和内含子分别由黄色方块和黑色线表示。 Twenty-five different color boxes represent 25 different motifs. Orange box represents exon, and black line represents intron. |
Intron和exon的数量和分布情况暗示了亚洲棉bZIP家族基因的进化印记, 同一个亚家族中的成员具有相似的基因结构, 不同亚家族和成员之间intron-exon的数量和位置是不同的(图2)。其中有19条基因序列没有内含子, 占整个GabZIP基因的11.95%, 这种现象出现在A、F和S亚族, 分别占7.69%、20.00%和57.69%。剩余的140个GabZIP基因有内含子, 内含子的数量在1~12之间变化, D和G亚家族中的成员内含子数目变化程度最大, 分别在5~12和6~11之间变动, 其余亚家族内含子的数量存在小范围的变化, 一般在1~4之间变动。
bZIP区域的碱性区(Basic region)和铰链区(Hinge region)是最保守的(图3), 根据Basic region和Hinge region中内含子的数目、位置和拼接位点, 将GabZIP基因划分为7种模式: a、b、c、d、e、f和g (图4)。其中模式a (包含31个基因)和b (包含39个基因)是最普遍的, 模式a在Hinge region 的-5位置有一个内含子(P0), 只有A亚家族的31个基因。模式b在Basic region的-22位置有一个内含子(P2), 它出现在C、E、I和W亚家族中。模式c有2个内含子(都在P0), 1个在Basic region的-22位置, 另一个在Hinge region的-6位置, 只出现在D亚家族。模式d在Hinge region的-6位置有一个内含子(P0), 只存在于G亚家族。模式e在Basic region的-19位置有1个内含子(P2), 只包含I亚家族的1个基因。模式f在Basic region的-20位置有一个内含子(P0), 只出现在V亚家族。模式g在Basic region和Hinge region都没有内含子, 有42个基因属于这个模式, 这42个基因分别包含在A、B、F、H、S和U亚家族中。由图4可以看出在Hinge region内含子只出现在P0处, 在Basic region内含子出现在P0/P2处。由于bZIP基因结构中Basic region和Hinge region中的重要性和保守性, 使GabZIP与OsbZIP和ZmbZIP蛋白的功能可能相一致。
通过对比对分析亚洲棉bZIP基因的序列, 共得到3个GaFD基因, 它们分别位于第1、第4和第10染色体上(表2)。根据与拟南芥AtFD基因的进化关系和蛋白氨基酸序列的比对分析, 将亚洲棉GabZIP13命名为GaFD1, 将GabZIP47命名为GaFD2, 将GabZIP70命名为GaFD3。Blast比对分析这3个GaFDs基因的CDS序列, 其中亚洲棉GaFD1与雷蒙德氏棉预测GrFD (GenBank登录号为XM_012613834.1)基因的相似性为97.1%, GaFD2与预测GrFD(GenBank登录号为XM_012632374.1)基因的相似性为97.4%, GaFD3与预测GrFD(GenBank登录号为XM_012586433.1)基因的相似性为97.3%。比较亚洲棉3个GaFD基因的ORF及ORF相对应基因组区域的序列, 分析这3个基因的外显子和内含子结构, 结果显示和拟南芥AtFD基因结构相似, 均由3个外显子和2个内含子组成(图5)。将3个GaFD蛋白与其他6种物种的FD蛋白, 即拟南芥AtFD (GenBank登录号为BN000021)、葡萄(Vitis vinifera) VvFD (GenBank登录号XP_ 003635259.1)、欧洲山杨和河北杨的杂交品种(Populus tremula × Populusalba) PtFD (GenBank登录号AGM48561.1)、马铃薯(Solanum tuberosum)
 | 图3 亚洲棉bZIP蛋白碱性区域和铰链区的比对Fig. 3 Alignment of basic and hinge regions of GabZIP protein in G. arboreum |
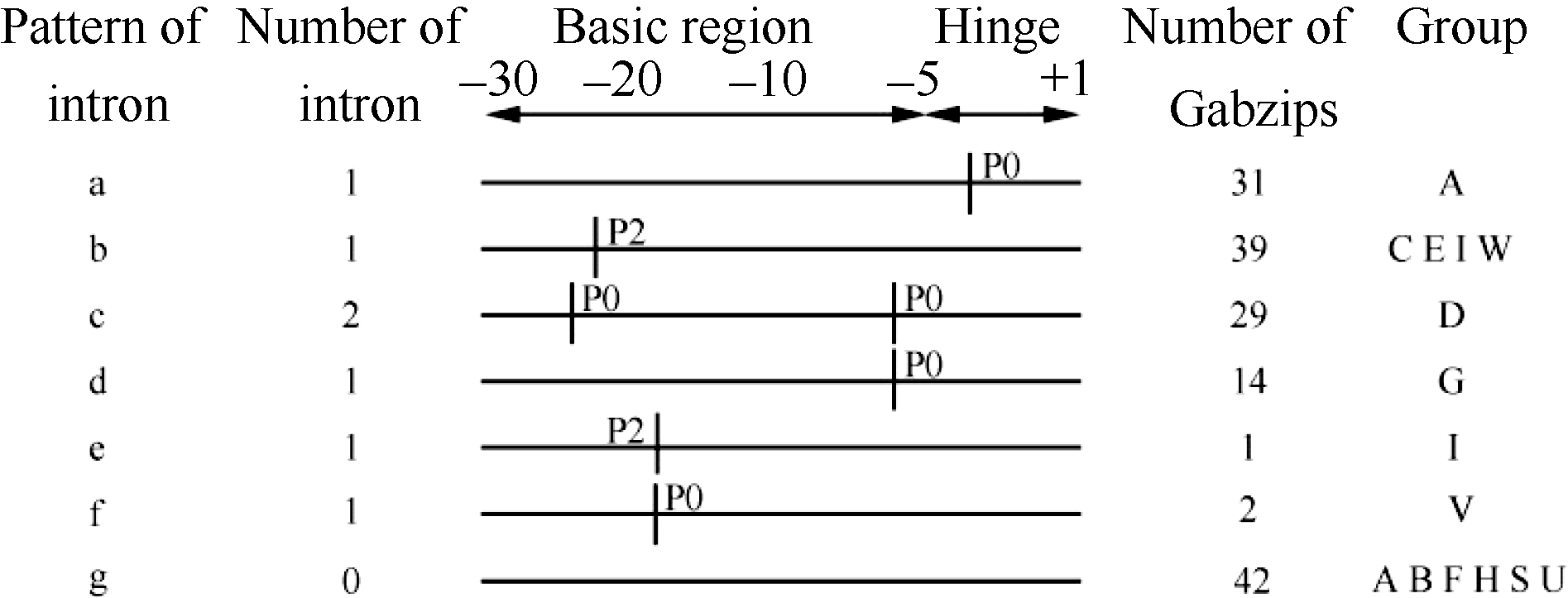 | 图4 亚洲棉bZIP基因碱性区域和铰链区内含子的分布模式Fig. 4 Intron distribution patterns within the basic and hinge regions of the GabZIP genes in G. arboreum +1是第1个亮氨酸(每7个氨基酸的第7个是亮氨酸), 碱性区域和铰链区用水平线表示, 内含子处用垂直线分隔。P0代表内含子拼接位点发生在2个密码子之间, P2代表内含子拼接位点发生在1个密码子的第2个和第3个核苷酸之间。 The first leucine in the amino acid heptad repeats is numbered +1, the basic and hinge regions are depicted by horizontal solid lines, Intron interrupted by vertical bars. P0 represent the intron splicing site between two codons, P2 means the intron splicing site located between the second nucleotide and the third nucleotide in one codon. |
StFD (GenBank登录号为AGM48561.1)、大豆GmFD (GenBank登录号为XP_006573542.1)和野草莓(Fragaria vesca) (GenBank登录号为XP_ 004289074.1)的氨基酸序列比对分析, 发现亚洲棉
GaFD蛋白中都存在A、LSL和SAP这3个保守基序(图6), 表明这3个序列是GaFD基因。
本研究通过信息学手段分析亚洲棉全基因组序列, 获得了159个亚洲棉bZIP基因, 它们分别位于第1~第13染色体上。通过与拟南芥和水稻bZIP基因家族聚类分析[9, 10], 将亚洲棉bZIP基因分为13个亚家族(图1), 其中A~S 10个亚家族同拟南芥分类
| 表2 亚洲棉GaFD基因信息 Table 2 Information for GaFDin G. arboreum |
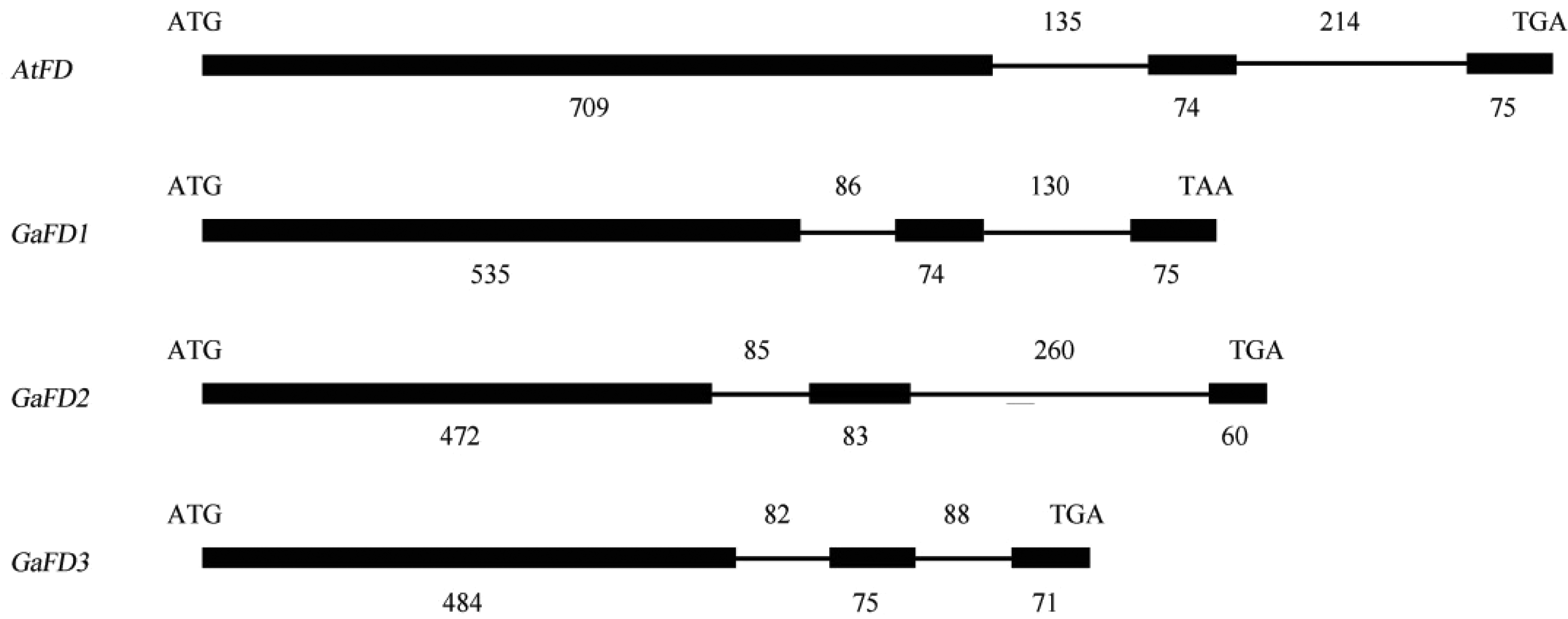 | 图5 亚洲棉GaFD基因外显子及内含子分析Fig. 5 Exons and introns analysis of GaFDgenes in G. arboreum ATG和TGA/TAA分别代表起始密码子和终止密码子, 黑色方框和直线分别代表外显子和内含子, 数字表示外显子和内含子的长度(bp)。 ATG and TGA/TAA represent initiation codon and termination codon, respectively. Black box and lines represent exonic and intronic regions, respectively. Numbers indicate the length of exons and introns (base pairs). |
 | 图6 亚洲棉与其他物种FD蛋白氨基酸序列比对Fig. 6 Amino acid sequences alignment of FD proteins from G. arboreum and other plant species A、LSL和SAP代表不同植物FD蛋白氨基酸序列中的3个保守基序。 A, LSL, and SAP represent three conserved motifs in FD protein amino acid sequences of different plant species. |
 | 图7 亚洲棉GaFD基因在不同组织中的表达特征Fig. 7 Expression patterns of GaFD genes in different tissues of G. arboreum R: 根; S: 茎; L: 叶; SA: 幼苗顶端分生组织; Fl: 花; Pi: 雌蕊; St: 雄蕊; O: 花后20 d的胚珠。 R: root; S: stem; L: leaf; SA: SAM; Fl: flower; Pi: pistil; St: stamen; O: the ovule at the 20 day after flowering. |
一致[9], U亚家族与大麦(Hordeum vulgare)和玉米(Zea mays)分类一致[42, 43], V和W属于新的亚家族。在拟南芥、水稻和玉米中, bZIP亚家族成员最多的分别是S (17个AtbZIP), A (17个OsbZIP)和D (40个ZmbZIP), 成员最少的分别是H、E、U (每个亚家族有2个AtbZIP), U (1个OsbZIP)和U (2个ZmbZIP)[9, 10, 43]; 在亚洲棉中, A亚家族是最大的一个进化分支, 包括38条GabZIP蛋白序列, B亚家族和H亚家族的成员是最少的, 分别只有1条GabZIP蛋白序列。与拟南芥bZIP基因比对发现与AtbZIP基因同源的GabZIP基因, 在进化上可能与AtbZIP基因功能相近, 如, A亚家族有38个GabZIP, 其中GabZIP31.3、GabZIP68.3和GabZIP90.1是拟南芥AtABF的同源基因, 主要参与ABA和逆境胁迫的调控表达[18]; B亚家族只有一个基因, GabZIP86; C亚家族有11个GabZIP基因, 其中GabZIP46.1和GabZIP18分别是AtbZIP10和AtbZIP25的同源基因, AtbZIP10和AtbZIP25的功能是调控特定种子基因的表达[44], 而GabZIP46.1和GabZIP18是否同样具有此功能还有待于验证; D亚家族有29个GabZIP基因, 该家族通过与TGACG (TGA)蛋白序列结合发挥它的作用[23, 25], 其中GabZIP53.3、GabZIP82和GabZIP87是AtTGA的同源基因; E和F亚家族分别有8个和5个GabZIPs基因; G亚家族有14个GabZIPs成员, 由于该家族成员与G-box序列结合而命名, GabZIP29、GabZIP44.3、GabZIP49.1是AtGBF的同源基因, 参与响应光启动子的调控[28, 45]; H亚家族只有一个成员, GabZIP59, 它是拟南芥AtHY5的同源基因, AtHY5基因调控在刺激诱导下下胚轴和根的发育[17]; I亚家族有17个GabZIP成员; S亚家族有26个GabZIP成员, 由于S亚家族成员的蛋白序列长度较短, 它可能需要与其它因子相互作用来完成转录激活[33]。
保守结构分析表明, 亚洲棉GabZIPs家族成员共有25个保守基序(图2), 这些保守基序可能与蛋白的功能有关, 也可能参与bZIP蛋白功能的激活。拟南芥、水稻和玉米的bZIP家族蛋白保守基序的鉴定都已经完成[9, 10, 43]。拟南芥A亚家族的保守基序中有酪蛋白激酶(CKII)磷酸化位点(S/TxxD/E), ABA能引起AtAREB1/2蛋白在此位点磷酸化进而诱导下游基因的表达, 在亚洲棉A亚家族的保守基序中也包含这个序列。拟南芥G亚族中有3个保守基序是富含脯氨酸激活结构域的一部分, 在亚洲棉G亚族中同样包含这一保守基序。说明不同物种bZIP家族之间具有一定的结构保守性, 这也是基因功能相似的原因。此外, 不同物种bZIP亚家族保守基序之间也存在着差异, 这也是物种之间保持特异性的原因。基因结构分析表明(图2), 亚洲棉有19条GabZIPs基因序列没有内含子, 这个现象出现在A、F、S和U亚族中, D和G亚族中内含子变化的数目最大, 在玉米[43]中, 没有内含子的序列出现在F和S亚族中, 内含子变化数目最大的也是D和G亚族, 说明在单双子叶植物中bZIP家族的基因结构具有相似性。在亚洲棉中, bZIP的Basic region和Hinge region是最保守的, 一般认为在这个区域中内含子的位置和拼接位点相对于其它区域对探索bZIP基因的同源进化更有意义[10, 46], 根据这个区域中内含子的数目、位置和拼接位点将bZIP基因划分为7种模式(图4), 其中内含子的数量与水稻[10]、玉米[43]、大麦[42] bZIP家族的分类基本一致, 拼接位点有所差异, 但和蓖麻[47]内含子拼接位点比较相似, 可能是单双子叶物种的差异所致。
bZIP A亚家族中的FD蛋白通过与FT基因编码的Florigen蛋白相互作用, 促进下游开花身份相关基因表达, 进而促进植物开花。亚洲棉GabZIP A亚家族有3个GaFDs基因, 分别位于第1、第4和第10染色体(表2)。结构分析表明, 亚洲棉3个FD基因与拟南芥FD基因的结构相同(图5), 并且蛋白序列中含有FD蛋白的3个保守基序(图6)。研究表明, 拟南芥fd-1突变体是明显晚花的, 在fd-1突变体中过量表达FD可以恢复它的正常表型[20], 说明FD基因具有促进开花的作用。但是亚洲棉3个FD基因是否也具有促进开花的功能, 或者各自起着不同的作用还需要进一步的验证。对拟南芥[10]和水稻[14]FD基因的研究表明, 成花素FT基因在子叶的维管组织和叶片中转录为mRNA并合成FT蛋白, FT蛋白再由韧皮部经过长距离运输到顶端分生组织, 与在SAM合成的FD蛋白相互作用激活下游信号, 促进植物开花, 在本研究中, 3个GaFD基因均在SAM中的表达量很高, 在叶片中只有微弱的表达, 或者不表达(图7), 这符合FD蛋白与FT蛋白的作用模式。但是, GaFD1和GaFD2在SAM中的表达量最高, 而GaFD3在茎中的表达量最高, 其次在SAM中表达, 这个现象说明GaFD3与GaFD1和GaFD2可能存在功能上的差异, 推测除具促进开花的作用外, 还有其他重要的功能。
总之, 在二倍体棉花G. arboreum (A2)和G. raimondii (D5)基因组中均存在bZIP家族基因, 四倍体陆地棉由A与D基因组进化而来, 四倍体棉花也可能具有A与D两套bZIP家族基因, 同时也不排除四倍体棉花bZIP基因家族在进化过程中由于重复基因间的相互作用只有一组二倍体基因组(A或D)来源的bZIP基因表达。随着陆地棉[38, 39]和海岛棉[40]基因组测序的完成, 为我们对四倍体棉花中究竟存在多少个bZIP基因, 相应的基因序列与二倍体有什么不同提供了数据支持, 对它们在棉花发育过程中的具体功能还需要进一步的深入研究。本研究获得的有关亚洲棉bZIP家族基因的信息, 为进一步分析棉花bZIP转录因子特别是棉花FD同源基因的功能和作用机制积累了有价值的资料。
基于亚洲棉(G. arboreum)全基因组数据库, 发掘出159个bZIP家族基因, 属于13个亚家族, 分布在13条染色体上。bZIP A亚家族存在3个GaFD基因, GaFD1和GaFD2在SAM中表达最高, GaFD3在茎中表达最高, 不同的表达特征表明它们可能具有不同的功能。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|