
{kind=link}
河南夏大豆区近30年主要大豆品种产量改良的遗传进展
[黄中文 , 徐新娟, 王伟, 梅沛沛]
, 徐新娟, 王伟, 梅沛沛]
, 徐新娟, 王伟, 梅沛沛]
|
|


第一作者联系方式: E-mail: huangdou373@126.com
提高产量是大豆育种的主要目标。研究大豆产量及其相关性状的遗传进展, 对于今后制定高产育种策略有重要参考意义。本研究随机选择近30年河南主要育成品种中的18个大豆品种, 进行两年产量评价试验的研究。结果表明, 产量随育种年份增加总体呈递增趋势, 遗传进展为17.39 kg hm-2, 年递增率是0.7%; 有效分枝、主茎节数、百粒重、株高有弱的正向遗传进展, 而单株荚数和每荚粒数表现弱的负向遗传进展, 但均不显著。百粒重、主茎节数和株高与产量有显著的遗传相关与表型相关, 环境相关均不显著, 表明这3个性状具有较大的遗传力, 随产量性状的遗传改良, 这3个性状均协同提高, 且不易受环境条件的影响; 而有效分枝、单株荚数、每荚粒数与产量的遗传相关和表型相关均没达到显著水平, 这是它们与百粒重、主茎节数、株高有极显著负遗传相关所致。
High yield is a main objective in soybean breeding. This study aimed at guiding soybean breeding by understanding genetic gain of yield and its related traits. 18 soybean cultivars randomly selected from cultivars released in Henan province during the last 30 years were evaluated in two years. The results showed an increasing trend of yield with the average annual rate of 0.7%, and the genetic gain was 17.39 kg ha-1. Effective branch number, node number of main stem, seed weight, and plant height showed a positive genetic gain, whereas pod number per plant and seed number per pod showed a negative genetic gain; however, these genetic gains were weak with no significance. Soybean yield had significantly genetic and phenotypic correlations with seed weight, node number of main stem, and plant height but had no significant correlation with environment. This result indicated that seed weight, node number on main stem, and plant height had high genetic heritability and were selected simultaneously with the genetic improvement of yield, with minor effect of environment. In contrast, effective branch number, pod number per plant, and plant height had no significant correlation with yield, mainly because of their negatively genetic correlations with seed weight, node number of main stem and plant height.
河南夏大豆区是我国黄淮海大豆主产区之一。河南省地处中原腹地, 生态资源优越, 其特定的生态环境生产的大豆具有高蛋白、高油“ 双高” 、适宜食用等优势, 育成的品种还可以直接推动豫、皖、苏、鲁等省的大豆生产发展。但由于河南省以及我国大豆在国际大豆市场竞争中处于不利地位, 河南省每年需进口大豆120万吨以上, 因此增加生产是缓解目前大豆供给矛盾的最有效手段[1, 2, 3]。但是通过扩大种植面积实现增产的策略没有太大的空间, 提高大豆品种的产量潜力, 是增加大豆生产的有效途径。
据美国植物生理学家Johnson[7]估计, 大豆的产量潜力可达15 t hm-2。2009年Tefera等[8]报道了尼日利亚稀树草原(Nigerian Savannas)的大豆在过去的20年时间内增加了1117~1710 kg hm-2; Cooper等[9]选育的半矮杆新品种Apex在高产环境下, 最高产量达5.98 t hm-2。意大利是世界上大豆单产最高的国家, 全国平均单产为3.6~3.7 t hm-2。
产量及其相关性状是典型的数量性状, 易受环境影响[10, 11, 12, 13]。计算作物产量遗传进展的方法很多, 其中把不同历史时期育成的品种种植在相同环境和统一栽培管理措施下进行产量比较是最常见的方法。在统一的环境下评价育成品种的遗传进展, 可以为制定育种策略提供重要的参考信息, 也可以为将来的育种实践提供现有品种农艺性状潜在利用价值[8], 这种研究方法已在几种主要作物上应用过。Tefera等[8]报道尼日利亚的大豆在过去20年内共增加了1117~1710 kg hm-2, Specht等[11]报道美国的大豆在1972— 1997年间年均增加了31.4 kg hm-2, Kamara等[12]报道尼日利亚地区1970— 1999年间释放的玉米品种遗传进展为0.41%。周阳等[6]报道30年中我国北部麦区小麦品种产量的遗传进展为1.2%, Jin等[14]对东北地区1950— 2006年育成的45个代表品种的研究表明, 产量遗传进展年递增率为0.58 %。
河南省大豆品种从20世纪60年代初期至今, 经过5次更新换代, 截止2007年, 共有54个品种通过河南省审定[15]。其主要农艺性状中, 生育期在106 d左右, 株高因品种而异, 变化较大; 产量构成因素中, 单株荚数总体呈增加趋势, 百粒重则相对变化较小[16]。然而尚未见系统研究分析其产量遗传进展、产量及产量相关性状演变规律的报道。本研究随机选择河南省近30年育成的主要大豆品种, 研究产量及主要农艺性状的遗传进展, 旨在为提高品种产量潜力和培育高产夏大豆品种提供一定的理论依据。
根据河南省5次更新换代的主要品种目录, 随机选择34个品种进行一年的产量及相关性状的评价, 作为本试验的前期预备试验。根据预试结果, 和在本地试验区的产量稳定性表现, 再从中随机选取18个代表河南夏大豆区大豆品种演变和发展的品种(表1)。
于河南科技学院乔谢大豆试验基地, 采用随机区组设计, 3次重复。每小区4行, 行长2.7 m, 行距0.4 m, 密度为19.5万株 hm-2, 2年分别于6月上旬种植。田间管理给予充足的肥水管理控制。10月上旬收获。
收获前, 在小区内选择连续生长的10株测定活体株高。从每小区收获中间2.5 m行长的植株, 随机选择10株单独装袋, 用于室内考种, 获得单株荚数, 每荚粒数, 主茎节数, 有效分枝、百粒重等性状, 其余装入网袋中, 风干, 脱粒, 获得的籽粒产量与10株产量相加即为小区产量。
采用SAS 9.4软件进行两环境联合方差分析及相关分析。
环境相关系数 
遗传相关系数 
表型相关系数 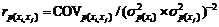
性状遗传进展按照下列公式计算[17]。
Yi=a+bCi+u (1)
ln(Yi)=a+bCi+u (2)
式中, Yi为品种i的表型值, Ci是品种i的育成年份, a为方程的截距, b是遗传进展, u是回归残差。方程(1)的b是性状的遗传进展, 方程(2)的100b是性状遗传进展的年递增率(%)。
2.1.1 产量 品种之间产量有极显著差异, 品种之间产量与环境的互作达到显著水平。参试品种平均产量为2641 kg hm-2, 豫豆27产量最高, 达到3310 kg hm-2, 其次是商丘1099、豫豆2号、周豆11、郑交107和郑交9525, 最低的是1978年推出的周7327-118, 仅为2160 kg hm-2 (表1)。随品种育成年份增加, 产量总体呈递增趋势, 遗传进展为17.390 kg hm-2 (R2= 0.29, P < 0.05)(图1), 年递增率是0.7% (R2= 0.33, P < 0.05)。
试验范围内, 大豆产量总趋势有两个高峰, 即1985和1988年推出的豫豆2号和豫豆8号为代表的第一个高峰, 以及以1999年推出的豫豆27为代表的第2个高峰, 30年产量变化总趋势是升高、降低、再升高。由表1和图1可见, 品种产量潜力变化可以分为3个变化阶段, 1990年以前育成的周7327-118, 商丘7608, 豫豆2号和豫豆8号为一个阶段, 其中豫豆2号产量最高, 达到2885 kg hm-2, 而周7327-118, 商丘7608和豫豆8号产量显著低于豫豆2号, 产量潜力呈增长趋势。第二阶段为1994— 1999年, 产量呈现先上升后下降再上升的趋势, 1999年育成品种豫豆27产量最高, 为3310 kg hm-2, 有效分枝也较高, 为3.3, 和豫豆21的有效分枝(3.7)差异不显著, 其株高也最高, 为107 cm, 该品种在1995— 1998年63点次试验中, 较对照豫豆8号平均增产9.5%, 其适应范围广, 对肥力要求不严, 中等以上肥力均可种植, 在河南省各地和安徽北夏大豆产区, 江苏、湖北、山东、山西、陕西等省与河南省接壤地区均可种植。第3个阶段为2000— 2004年, 2002年育成的品种商丘1099产量较高, 为2857 kg hm-2, 与之后育成的品种产量差异不显著, 品种产量潜力递增不明显。从品种产量潜力演变阶段分析, 第1和第2阶段大豆育成品种产量潜力有显著提高, 2000年后育成品种产量进展缓慢。
| 表1 产量及相关性状的方差分析 Table 1 ANOVA of yield and related traits |
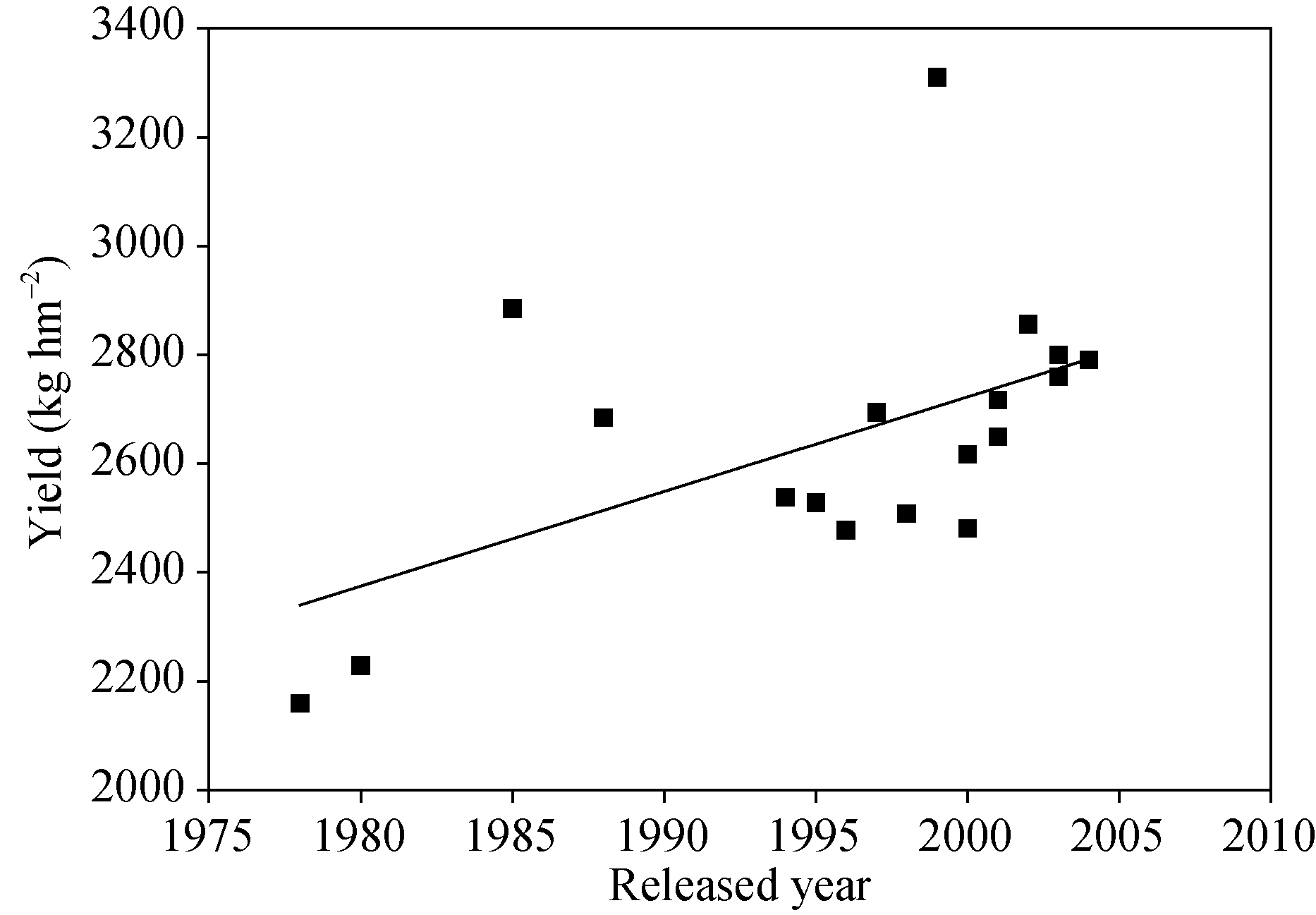 | 图1 产量对品种育成年份的回归Fig. 1 Regression of yield on released year |
2.1.2 产量相关性状 方差分析结果显示, 单株荚数、每荚粒数和百粒重品种间、基因型与环境互作均有极显著差异; 株型性状中有效分枝、主茎节数和株高品种之间有显著或极显著差异, 但基因型与环境互作均不显著。
单株荚数呈负遗传进展, 为-0.242, 但未达到显著水平。2001年育成的郑9007单株荚数最多, 达到71.8, 其次为1978年育成的周7327-118和2001年育成的豫豆29。1978、1980、1985、1988育成的4个品种中, 商丘7608和周7327-118差异显著, 其他的差异不显著; 1994— 1999年育成的6个品种中, 豫豆27、豫豆22、豫豆21和周7327-118差异不显著, 豫豆25、豫豆16、豫豆19和周7327-118差异显著。豫豆19单株荚数最少, 仅为41.2。
每荚粒数呈现先增加后减少的趋势, 有弱的负向遗传进展。1995年育成的豫豆19每荚粒数最多, 达到2.7个, 其次为2003年育成的郑交107和2002育成的商丘1009, 每荚粒数和豫豆19差异不显著, 均为2.5个, 其他年份育成的15个品种每荚粒数和豫豆19差异均显著, 其中豫豆29每荚粒数最少, 仅为2.1个。
百粒重均值为20.2 g, 遗传进展为0.005。品种年份之间百粒重的变化没有一定的规律。2003年育成的周豆11百粒重最大, 达到27.2 g, 其他年份育成的17个品种百粒重和周豆11差异均显著, 其中2003年育成的郑交107百粒重最少, 仅为14.9 g。
有效分枝随着品种育成年份呈先增加后减少的趋势, 遗传进展为0.008。1996年育成的豫豆21最多, 达到3.7, 之后育成的豫豆系列品种中, 豫豆28最少, 有效分枝仅为1.9。1988年之前育成的4个品种中, 豫豆2号有效分枝为3.1, 与豫豆21差异不显著; 2000— 2004年育成的8个品种中, 郑交9525有效分枝为3.6, 与豫豆21差异不显著, 商丘1099和豫豆28差异显著, 有效分枝分别为2.3和1.9。
主茎节数随着品种育成年份呈先增后减再增的趋势, 遗传进展为0.049。1985年育成的豫豆2号主茎节数最多, 达到17.6, 之后育成的豫豆系列品种中, 豫豆19、豫豆22、25、29、28无显著变化, 豫豆8最少, 主茎节数仅为13.4。2000— 2004年育成的8个品种中, 郑豆11主茎节数为17.0, 与豫豆2号差异不显著, 差异不显著的还有郑90007、豫豆28、商丘1099、郑交15.8和郑92116, 郑交107差异显著, 主茎节数仅为13.6。
株高均值为87 cm, 遗传进展为0.042。1999年育成的豫豆27株高最高, 达到107 cm, 其次为2000年育成的豫豆28和2002年育成的商丘1009, 株高和豫豆27差异不显著, 分别为99 cm和97 cm, 其他年份育成的15个品种株高和豫豆27差异均显著, 其中2003年育成的郑交107株高最矮, 仅为65 cm。
从表2可以看出, 产量的表型相关、遗传相关和环境相关的数值大多为正值, 其中百粒重、主茎节数和株高的遗传相关和表型相关均大于环境的相关, 达到极显著水平(P < 0.01), 说明百粒重、主茎节数和株高与产量有显著的遗传相关与表型相关, 表明这3个性状具有较大的遗传力, 对杂种后代产量性状的改良起着关键作用, 受环境条件影响不大。有效分枝、单株荚数及每荚粒数与百粒重的表型相关、遗传相关和环境相关均为负值, 有效分枝、单株荚数、每荚粒数与产量的遗传相关和表型相关均没达到显著水平, 但与产量的偏相关均达到极显著水平, 这是因为它们与百粒重、主茎节数、株高有极显著的负的遗传相关性, 选择育种材料的时候, 首先选择百粒重、主茎节数和株高性状优良的品种作为母本进行杂交育种, 以提高大豆产量。
有效分枝与单株荚数的表型相关、遗传相关和环境相关为正相关, 有效分枝性状对单株荚数影响大, 其中表型相关(0.551)和环境相关(0.492)达到极显著水平, 遗传相关(0.360)不显著。其中有效分枝与主茎节数、株高和每荚粒数的表型相关、遗传相关和环境相关为负偏相关。
主茎节数与株高的表型相关、遗传相关和环境相关为正值, 株高对主茎节数呈正相关, 其中表型相关(0.692)和环境相关(0.500)达到极显著水平, 遗传相关(0.102)不显著。其中主茎节数与每荚粒数的表型相关、遗传相关和环境相关为负值, 呈偏相关, 遗传相关极显著, 说明主茎节数越多, 每荚粒数越少, 大豆偏于营养生长, 从而影响了产量的增加。
单株荚数与每荚粒数遗传相关为正值(0.080), 呈正相关, 单株荚数影响每荚粒数, 其中环境相关达到显著水平, 单株荚数与株高遗传相关为正值(0.098), 呈正相关, 株高影响单株荚数, 每荚粒数和株高的表型相关、遗传相关和环境相关为负值, 呈偏相关, 说明株高越高, 每荚粒数偏少, 另外从大豆植株特性分析, 株高偏高, 主茎节数偏多, 与前面分析的主茎节数与每荚粒数的表型相关、遗传相关和环境相关为负值, 呈偏相关, 结果一致。
| 表2 产量及产量相关性状间的遗传相关系数 Table 2 Coefficients of genetic correlation between yield and related traits |
从本试验表现出的河南省大豆品种产量演变的阶段性特点分析, 1985年豫豆2号品种的育成体现出大豆产量的增高, 之后育成的品种产量提高较慢, 在1999年育成的豫豆27产量达到了近30年育成的大豆品种产量的最高峰, 产量达到了3310 kg hm-2, 年递增率为0.7% (P < 0.05)。Jin等[14]以东北地区1950— 2006年育成的45个代表品种试验表明, 产量年平均进展0.58%, 说明黄淮夏大豆区相比东北区域育成的大豆品种, 产量的遗传进展略有增加, 但低于美国大豆遗传进展的相关报道[11]。我国大豆产量曾经在19世纪50— 70年代停滞不前, 在80年代才开始大量相关研究, 大豆产量出现不断增加的趋势[2, 5, 16, 18], 大豆品种遗传改进的明显趋势在于抗倒伏性显著增强、单株粒重提高、每节荚数、每荚粒数增多、粒重变大、茎秆增粗、株高降低[18]。
影响大豆产量提高的因素很多。首先, 很多研究者从大豆的主要农艺性状入手, 研究不同性状对大豆产量的贡献, 王振民等[19]研究吉林省生产上曾大面积推广的42个大豆品种的主要农艺性状的演变趋势发现, 产量与分枝数呈负相关; 产量与有效节数、每荚粒数、百粒重呈正相关, 与本研究结果类似。本试验从主要产量的遗传因素来看, 在这期间育成的品种改良的农艺性状着眼点不同, 1996年育成的豫豆21的有效分枝有明显提高, 达到3.7, 1995年育成的豫豆19每荚粒数最多, 为2.7。谢甫绨等[20]研究了辽宁省育成的50个品种的主要农艺性状遗传改进的趋势, 其中分枝荚数和三粒荚数的变化最大, 单株粒重、百粒重、主茎节数及分枝荚数变化中等, 其余各性状变化较小。郑洪兵等[21]对吉林省不同年代大豆品种株型性状的演变研究表明, 产量、叶面积、叶面积指数、茎直径、比叶重随着年代的增加而增加。
其次, 开展了大量关于大豆产量相关性状的遗传分析, 研究了大豆各个性状的遗传力及其相关性, 大豆株高、主茎节数、有效荚数、荚粒数、株粒重、分枝数与百粒重等是影响和构成大豆产量的重要因素[22, 23, 24, 25, 26]。本试验发现, 近30年育成的大豆品种产量变化主要集中在主茎节数、百粒重和株高上, 产量与三者成正相关。可能因为夏大豆生长期短, 光周期反应明显, 不易形成分枝, 另外, 7月和8月大豆开花期的持续阴雨对有限结荚习性的品种非常不利, 因为有限结荚品种开花期时间短而集中。刘萌娟等[26]研究发现, 小区产量和主茎节数遗传相关显著, 选育高产品种的重要辅助性状为主茎节数和百粒重, 因此在田间选择时, 重点选择主茎节数多、籽粒中等偏小的品种, 有望获得理想产量育种效果。近年来, 随着分子育种技术研究的不断发展, 与产量、品质、抗性有关的QTL数目越来越多, 基于分子水平的聚合育种技术也日趋完善[27, 28, 29], 研究大豆产量的相关因素和遗传进展, 也为大豆分子育种技术提供重要参考信息。
河南夏大豆区近30年大豆育成品种产量的年均遗传进展为17.390 kg hm-2 (P < 0.05), 年递增率为0.7%。在产量相关因素中, 主茎节数的年均进展相对较大, 其中有效分枝、主茎节数、百粒重、株高的正向遗传进展较弱, 而单株荚数和每荚粒数是负向遗传进展, 均不显著。百粒重、主茎节数和株高与产量有显著的遗传相关与表型相关, 环境相关均不显著, 表明这3个性状具有较大的遗传力。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|