

{kind=link}
{kind=link}
{kind=link}
江苏主栽粳稻品种的遗传与食味结构分析
[龚红兵1, 2 , 曾生元2 , 李闯2 , 左示敏1 , 景德道2 , 林添资2 , 陈宗祥1 , 张亚芳1 , 钱华飞2 , 余波2 , 盛生兰2 , 潘学彪1, *  ]
]
]
|
|


第一作者联系方式: E-mail: hongbinggong973@sina.com.cn, Tel: 13812369841
利用SSR分子标记和主要食味性状分析江苏主栽粳稻品种的食味结构。50对SSR引物中有41对存在多态性, 41对多态性标记在86个品种间共扩增出113个等位基因, 每个标记的等位基因数目为2~6个, 平均2.76个; 多态信息含量(PIC值)变异范围为0.02~0.84, 平均为0.48; Nei’s基因多样性指数变幅在0.02~0.68之间, 平均为0.37; 遗传聚类分析可将86个品种分为3个类群。在正季长日照情况下, 品种间直链淀粉及蛋白质含量在遗传聚类的I、II、III类中呈现出递减的趋势, 食味分值则反之; 而在短日照条件下, 第II、第III大类型品种的直链淀粉及蛋白质含量较正季显著升高, 食味值显著下降, 而第I类型品种降幅不大; 总体而言, 江苏主栽粳稻品种的食味一般且遗传背景相近, 拓宽遗传基础是培育环境钝感型优质食味粳稻的重要途径。
Fifty indica-japonicaspecific SSR markers were employed to genotype 86 conventional japonica rice cultivars mainly planted in Jiangsu province and 41 loci showed polymorphism among them. After amplifying in 86 rice varieties, a total of 113 alleles were detected at the 41 SSR loci, and the allele number ranged from two to six, with an average of 2.76 in each locus. Polymorphism information content (PIC) value of these 41 SSR markers ranged from 0.02 to 0.84, with an average value of 0.48; Nei’s expected heterozygosity value of the 41 markers ranged from 0.02 to 0.68, and the average value was 0.37. Based on the coefficient index ranged from 0.72 to 1.00, we divided the 86 cultivars into three groups by UPGMA. The analysis of taste quality traits investigated under different environments indicated that the taste quality of conventional japonica rice cultivars in Jiangsu was correlated with the genetic clustering. Under long-day (LD) condition, the amylose content (AC) and protein content (PC) manifested a tendency of group I > group II > group III, while the taste value (TV) was opposites. And under short-day (SD) condition, taste quality traits of group II and group III varied significantly compared with that under LD condition, while the traits of group I had no significant change. The results indicated that the main cultivars grown in Jiangsu have a general taste quality and narrow genetic background, broadening the genetic basis is an important approach to breed japonica rice cultivars with environment- insensitivity and good eating quality.
江苏是我国粳稻米的主产区, 约生产全国粳稻谷年总产量的1/3 [1], 江苏粳稻单产长期维持在8.25~9.00 t hm-2, 平均单产水平全国最高。2000年以来, 江苏省粳稻育种的目标已从高产、优质、多抗转向优质、高产、多抗, 优质成为第一位[2]。通过常规育种结合分子标记辅助选择的手段, 近年来育成了一批理化品质优异的粳稻, 以及南粳46、南粳5055为代表的优质食味“ 软米” 品种[3, 4], 但是从饮食习惯来看, 软米为半糯性, 多吃易腻, 而真正食味优良品种不多。武育粳3号作为20世纪90年代育成的代表品种, 其抗性、稳产性已经面临诸多挑战, 但是由于其具有较为突出的优质食味特性, 近年在生产上仍具有较大的应用面积[5], 这也从一个侧面反映出江苏粳稻在食味品质改良方面滞后。
传统的水稻育种主要是通过肉眼来选择杂交后代分离群体中的优良个体。然而, 稻米品质由大量的数量性状位点(QTL)控制, 在杂交后代表现出连续的表型变异, 肉眼难以有效判断, 也缺乏连续型性状分离的有效统计和选择方法, 因此采用传统的方法来提高水稻品质较为困难。此外, 我国国土面积宽广, 不同地域间生态环境存在显著差异, 直接形成了适宜不同生态类型的粳稻品种, 粳稻米品质受环境影响变异幅度大, 在不同地区难以保持其“ 优质食味” 特性, 成为限制许多优良粳稻品种进一步推广应用的重要因素。因此, 选育综合品质好, 对温光反应钝感的优质食味粳稻品种仍是江苏育种工作者需要长期重视和解决的问题。
虽然培育优质食味水稻新品种在实际育种工作中仍面临很大的困难与挑战, 但随着水稻基因组研究和现代制造业技术的发展, 分子生物学及诸多快速测定稻米品质技术的发展为我们提供了新的机遇。遗传多样性是衡量种内或种间遗传变异度的重要指标, 是作物遗传改良的物质基础, SSR标记能够准确有效地鉴定出水稻序列的差异, 具有较高的多态性、共显性、稳定性高、技术简便快捷等特点, 被广泛应用于水稻群体遗传结构分析、物种进化、遗传图谱构建、基因定位及分子标记辅助育种等研究领域[6, 7, 8]。在对稻米食味的鉴定方面, 采用快速食味计实现高效快捷的筛选已逐渐成为可能。本研究利用SSR标记并结合快速食味仪测定稻米食味性状进而分析江苏主栽品种食味性状的遗传结构, 以期为培育环境钝感型优质食味粳稻品种提供理论依据。
常规粳稻品种87份, 包括2001— 2012年江苏省审定的常规粳稻品种83份, 典型的对照品种4份。82份材料中中熟中粳类型品种23份, 含1997年通过审定的镇稻88; 迟熟中粳类型品种29份, 对照为1992年通过审定的优质食味品种武育粳3号; 早熟晚粳类型品种22份和中熟晚粳类型品种12份, 对照均为1998年通过审定的高产但食味一般的武运粳7号(供试品种名称等信息见附表1), 另有食味测定对照日本品种越光。
2012与2013年正季、2012年海南两年3季分别在江苏丘陵地区镇江农业科学研究所句容市后白试验基地和陵水南繁基地适期播种87份亲本材料, 湿润育秧, 按随机区组方式每份材料种植, 3个重复, 每重复100株。行株距25.0 cm × 13.3 cm, 按常规方法田间管理。
1.3.1 DNA提取和PCR反应 参照McCouch等[9]的CTAB法略有改动提取水稻基因组DNA。PCR扩增含模板DNA 1.0 μ L、10× PCR的缓冲液2.0 μ L、25 mmol L-1的MgCl2 2.0 μ L、2 mmol L-1的dNTP 2.0 μ L、0.3 μ mol L-1的引物2.0 μ L、Taq酶0.5 U, 加ddH2O补足20 μ L; PCR扩增条件为: (1) 94℃预变性5 min; (2) 94℃变性50 s, 55~60℃退火50 s (温度因引物不同而异), 72℃延伸50 s (不同长度的预期产物按1 kb每分钟调整延伸时间), 扩增32个循环; (3) 72℃延伸10 min, 4℃保温。PCR产物经3%琼脂糖凝胶电泳检测及8%垂直平板聚丙烯酰胺凝胶电冰, 0.1%硝酸银染色后观测。
1.3.2 SSR引物与数据分析 参照已发表的分子标记遗传图谱, 选用均匀分布于水稻全基因组12条染色体上的50对SSR引物, 由生工生物技术(上海)股份有限公司合成。
根据PCR扩增结果, 在相同迁移位置上, 有带记为1, 无带记为0, 缺失数据赋值为9。统计每个SSR位点多态性信息量(polymorphism information content, PIC)[10], 并利用Popgene软件计算SSR位点的多态性信息[11], 采用NTSYS-pc ver2.1 0软件计算遗传相似矩阵, 按UPGMA法聚类[12]。
在Microsoft Excel、SPSS17.0软件上进行数据整理和方差分析, 采用Duncan’ s新复极差检测法多重比较。
利用日本佐竹公司生产的食味仪(RCTA-11A)测定食味品质。称取每一品种500 g, 出糙后用JY7132型精米机碾成精米, 称量130 g放进样品槽, 1 min后读出食味分和直链淀粉、蛋白质、水分的含量。每个样品被重复测定3次, 取平均值。
选用每条染色体4~5对, 共50对SSR引物, 其中9对引物(RM71、RM438、RM413、RM267、RM152、RM219、RM409、RM258、RM147)为单态位点, 其余41对具有多态性, 多态性位点百分率为82.0%。
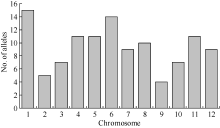
41对引物在86个品种中共扩增出113个等位基因, 品种间不同位点等位基因数目不等, 范围为2~6个, 平均2.76个, 以等位基因数目频次为2的引物最多, 有19对, 其次为3个的, 有16对, RM430在86份品种间扩增出6个等位基因。41对SSR 标记的多态信息含量(PIC)平均为0.48, 其变异范围为0.02~0.84, 其中低多态位点(PIC < 0.25)有10个(表1)。经Popgene计算平均Nei’ s基因多样性指数为0.37, 略低于PIC指数, 变幅在0.02 (RM219) ~ 0.68 (RM336)之间。12条染色体中第1染色体等位基因总数最多, 第9染色体最少(图1)。
| 表1 41对有多态的SSR引物的位置以及在86份江淮稻区主栽品种中的遗传多样性信息 Table 1 Location, number of alleles and genetic diversity of 41 SSR loci in 86 Jianghuai-river area major rice varieties |
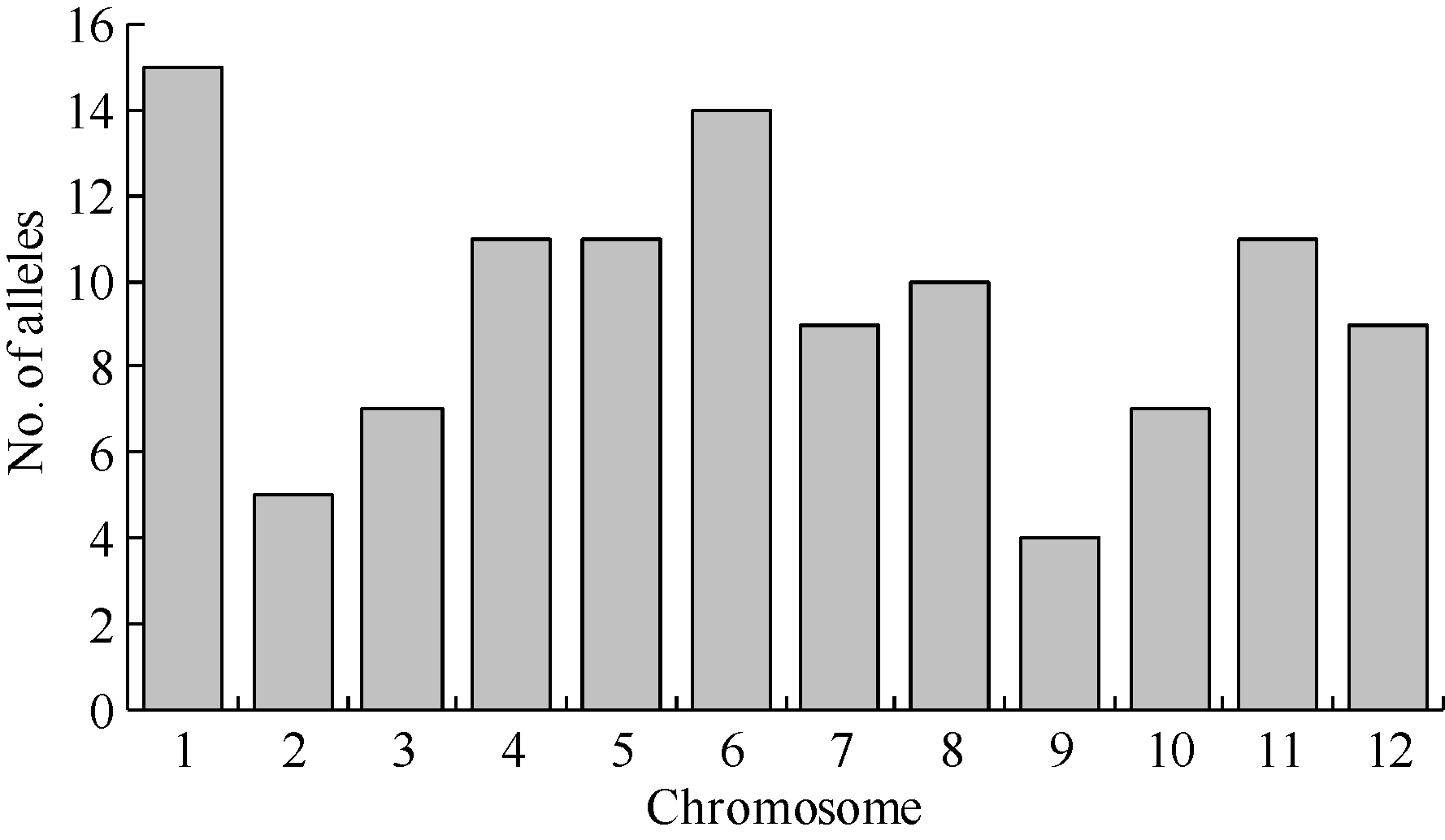 | 图1 12条染色体的等位基因数目分布Fig. 1 Frequency of allele number in each chromosome |
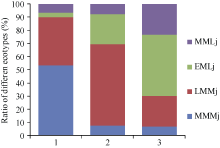
41对SSR引物在86份供试亲本中扩增得到113个等位基因, 利用NTSYS-pc2.0统计分析软件计算86个亲本间的遗传相似系数(coefficient), 其范围在0.72~1.00之间。以遗传相似系数为原始数据, 用UPGMA法对86个亲本聚类分析, 由图2可知, 以相似系数0.73为阈值, 可将86个亲本分为3大类群。其中镇稻88、连粳4号等30个品种划分为第I大类群, 武育粳3号等13个品种为第II大类群, 武运粳7号、淮稻5号等43个品种属于第III类群(见附表1和图2)。
从遗传相似系数可以看出, 江苏粳稻品种间存在高度的同质性, 这与前人研究结果相似[13, 14]。本研究的分子标记聚类结果与江苏粳稻的生态型划分高度相关(表2和图3), 究其原因, 粳稻具有强烈的地域性, 在不同的适宜种植区域已经形成了对基本营养生长性、感温性、感光性的定向选择。从总体上看, 本研究的类群划分结果与周振玲等[14]的分类结果大体一致。
 | 图2 江苏主栽粳稻品种的遗传聚类图 MMMj: 中熟中粳; LMMj: 迟熟中粳; EMLj: 早熟晚粳; MMLj: 中熟晚粳。Fig. 2 Molecular coefficient index and the clustering of main cultivars grown in Jiangsu province MMMj: medium maturing mid-season japonica rice; LMMj: late maturing mid-season japonicarice; EMLj: early maturing late japonica rice; MMLj: medium maturing late japonica rice. |
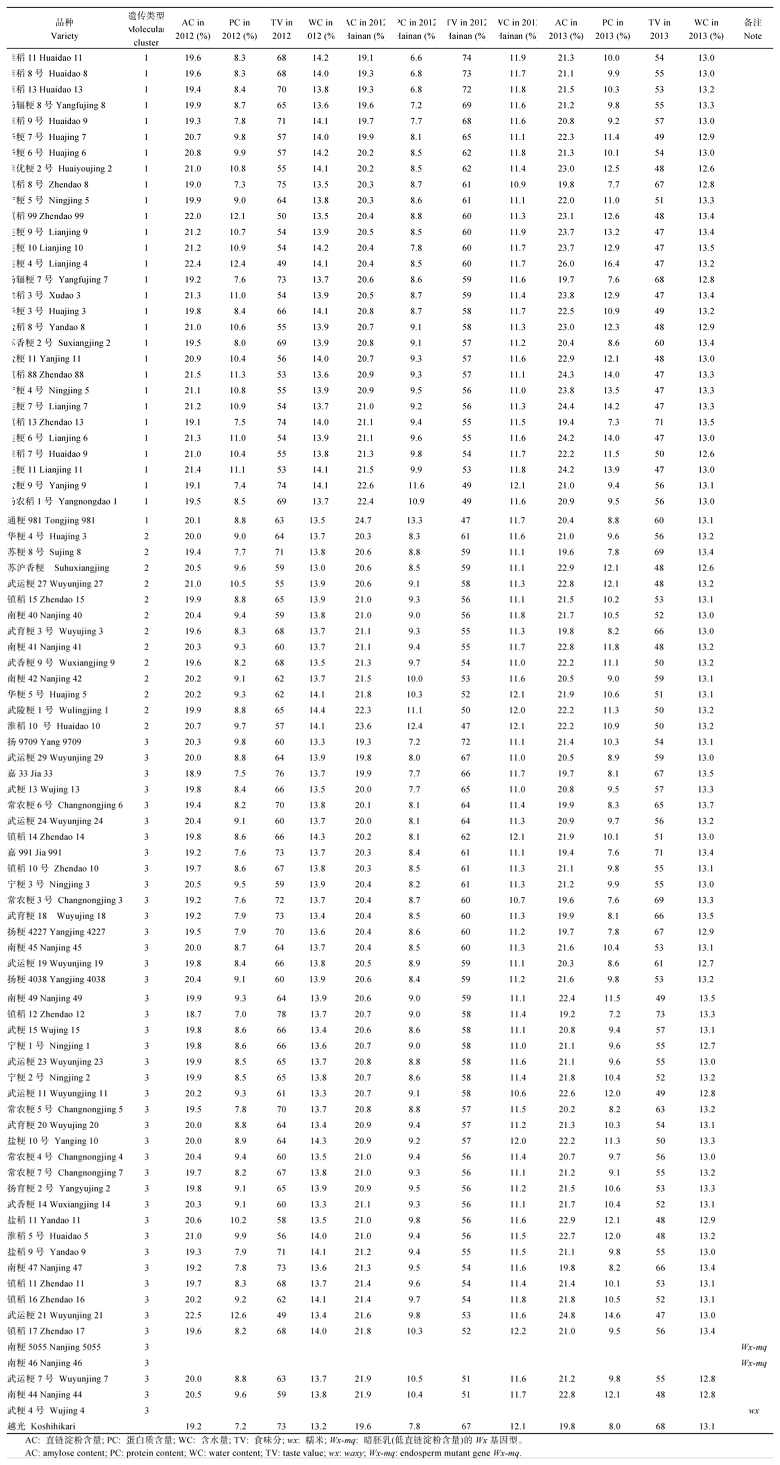
通过两年两地三季对86个江苏主栽粳稻品种的食味品质测定发现, 在正季情况下虽未达到统计学的显著水平, 但可以看出品种的直链淀粉含量以及蛋白质含量呈现出分子标记聚类I、II、III类递减的趋势, 食味分值则反之(表2), 其中对照品种越光为2012年的食味分值为73, 2013年为68; 镇稻88的2012年食味分值为53分, 2013年为47分; 武育粳3号2012年食味分值为68, 2013年为66; 武运粳7号2012年食味分值为63, 2013年为50; 而在短日照条件下, 第II、第III大类型品种的直链淀粉及蛋白质含量较正季显著升高, 食味值显著下降, 而第I类型品种降幅不大, 例如镇稻88的2012年海南食味分值为57, 而武育粳3号为55、武运粳7号为51。
此外, 与2012年相比, 相同品种2013年的直链淀粉以及蛋白质含量均有较大幅度的增加, 食味值显著下降, 其主要原因是2013年抽穗期(7月底至9月上旬)当地气温长时间持续在33℃以上, 罕见的异常高温年份(http:// www.tianqi.com/)导致了食味品质的下降(见附表2)。
镇稻12、镇稻13、嘉991、苏粳8号、常农粳3号等5个晚粳品种以及迟熟中粳品种扬辐粳7号2012年和2013年正季的食味值都优于对照越光, 或者与越光相当, 但是在海南, 这些品种的食味分值都显著低于越光, 而在海南表现出高食味值(分值高于越光)的则是淮稻11、淮稻8号、淮稻13、淮稻9号、扬9709等几个中熟粳稻品种, 除扬9709属于第3类群外, 其余4个品种均属于遗传聚类的第I大类群, 且来源相似(大多淮稻系列品种)(见附表2)。
| 表2 不同遗传类群江苏主栽粳稻品种的食味品质特征 Table 2 Characteristics of taste quality between different genetic groups of japonica rice varieties |
直链淀粉和支链淀粉含量的比例影响淀粉粒的结构和特点[15], 而直链淀粉含量的高低则是稻米品质优劣的决定因素[16]。研究表明直链淀粉合成主要由Wx基因控制, Wx基因第一内含子剪切位点上游55 bp处具有一个(CT)n重复的多态性位点, 现已证明该多态性位点也与稻米的直链淀粉变异存在显著的相关性[17], 万映秀等[18]研究表明SSR标记RM190揭示的水稻Wx基因多态性可以解释直链淀粉含量变异的59.3%, 且RM190至少已存在8种多态型, 即(CT)20、(CT)19、(CT)18、(CT)17、(CT)14、(CT)11、(CT)10和(CT)11/(CT)18等。本研究的聚类结果中也采用了RM190标记, 但只在江苏粳稻中筛选出2种等位变异, 其大小均在110 bp左右, 参照万映秀等[18]的结果[其结果中镇稻99的基因型为(CT)17], 本研究的RM190为(CT)17、(CT)18两种基因型, 且大多属(CT)17, 因此, 除了遗传相似系数显示江苏粳稻品种间具有较高的同质性外, 从这一侧面亦可以看出江苏粳稻品种的品质遗传基础狭窄。本研究表明除个别软米品种(如南粳46、南粳5055)直链淀粉含量低于13%外, 其余大部分品种食味仪测定值均在18.7%~22.5% (2012年数据)之间, 且上述2种基因型间直链淀粉含量的整体差异不大(详细数据见附表2); 而2013年为19.2%~26.0%, 变幅较大很可能的原因是2013年7月下旬至9月上旬出现了连续的异常高温天气, 部分不耐高温的中粳品种结实率及品质严重下降。
日本佐竹公司生产的RCTA-11A食味仪与理化测定方法不同, 它是通过近红外原理来测定稻米食味品质的。本研究测定的精米中直链淀粉含量、蛋白质含量均高于理化法测定值, 一般高出2个点(参照品种审定通告数据), 但是二者趋势一致, 即理化测定值高的食味仪测定也是高, 反之亦然。前人研究结果表明食味仪测定的食味值与食味品尝综合值之间存在极显著相关性[19], 本研究结果进行相关分析也发现, 食味值与直链淀粉及蛋白质含量的相关系数达到极显著的-0.979、-0.967。大量研究已证明直链淀粉及蛋白质含量是水稻的食味品质主要决定因素, 因此, 除了对糯稻(如武粳4号)、低直链淀粉含量品种(如软米品种南粳5055等)的测定不准确外(数据明显偏离正常值), RCTA-11A食味仪可以作为粳稻米食味评价的快捷、有效工具。
本研究表明, 江苏粳稻品种的品质遗传基础狭窄, 以第I大类型的品种食味品质对环境较为钝感但是在江苏本地普遍较差, 而第II、第III类型的品种对环境敏感, 其“ 品质广适性” 还很脆弱, 且大多与越光相比仍存在一定差距, 而对照品种越光无论是在海南还是江苏正季均能表现出较好的食味品质。因此, 改良江苏粳稻食味品质较为合理的途径是, 在扩大遗传背景的基础上, 选用食味品质好的品种与第I大类型的本地优良品种配组, 结合多年多点的食味测定, 选育对环境钝感的优质食味新品种。此外, 通过分子标记辅助的方法, 精确选择诸如低直链淀粉、低蛋白、具BADH2香型基因型的新品种也是改良江苏粳稻食味品质的一条快捷途径。
| 附表1 江苏主栽粳稻品种的生态类型及分子标记聚类结果 Supplementary table 1 Ecological type and molecular cluster of major japonica varieties in Jiangsu province |
| 附表2 江苏主栽粳稻品种的食味品质特征 Supplementary table 2 Charateristics of taste quality of japonica rice varieties in Jiangsu province |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|