




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
大豆盐胁迫相关 GmNAC基因的鉴定、表达及变异分析
[张彦威1, 2 , 张礼凤1 , 李伟1 , 王彩洁1 , 张军1 , 徐冉1, *  ]
]
]
|
|


NAC基因在植物的逆境胁迫中发挥着重要作用。本研究参照水稻和拟南芥的逆境相关 NAC基因, 采用生物信息学方法鉴定了大豆逆境相关 GmNAC基因, 利用荧光定量PCR技术分析了 GmNAC基因在耐盐差异的大豆品种根部、叶片的表达及其对NaCl胁迫的应答, 采用反转录PCR技术克隆了表达差异显著的 GmNAC基因。结果表明, 大豆 GmNAC基因家族包含175个基因, 其中11个基因所编码的GmNAC蛋白与水稻和拟南芥的逆境相关NAC蛋白位于同一进化分支, 这些蛋白具有高度保守的NAC结构域; 这11个 GmNAC基因在大豆根部的表达均高于在叶片, 而且在叶片和根部均受NaCl诱导, 部分基因在根部和叶片以及品种间表现出不同的表达规律; 在大豆品种齐黄34、徐豆10和汾豆95中, Glyma06g11970.1存在3个同义突变和1个非同义突变, Glyma06g16440.2存在1个同义突变。
NAC genes play an important role in plant stress tolerance. In this study, bioinformatic method was used to identify the stress related GmNAC gene in soybean. The expression of candidated GmNAC genes in root and leaf was analyzed in soybean with NaCl treatment by Real-time PCR. Reverse transcription PCR was performed to clone genes with significant difference in expression. The results showed that there were 175 genes in soybean GmNAC gene family. There were 11 GmNAC proteins with highly conserved NAC located on the same evolutionary branch with the stress related NAC proteins in rice and Arabidopsis. The expression of 11 GmNAC genes in soybean root was higher than that in leaf. The GmNAC genes were all induced by NaCl stress, but part of the GmNAC genes showed different expression levels between root and leaf in soybean varieties with different salt tolerances. There were three synonymous mutations and one non-synonymous mutation on the CDS region of Glyma06g11970.1 and one synonymous mutation on the CDS region of Glyma06g16440.2 in Qihuang 34, Xudou 10, and Fendou 95.
高温、低温、干旱和盐碱等非生物逆境胁迫严重影响作物的生长, 造成作物大幅度减产[1]。NAC蛋白是一类植物特异的参与多种植物生理生化过程的转录因子[2], 广泛参与植物生长发育以及逆境应答。NAC(NAM, ATAF1-2, CUC2)最初起源于NAM (no apical meristem), ATAF1-2和CUC2 (cup-shaped cotyledon) [3]。NAC蛋白包含一个保守的N-端DNA结合域和一个多样化的转录调控域[4]。NAC家族成员能够激活植物的逆境应答基因, 提高植物对逆境的耐受性。AtNAC072和AtNAC019是拟南芥的ABA信号通路的关键基因, AtNAC072的表达受高盐、脱落酸(ABA)、生长素、乙烯、茉莉酸(JA)、脱水、损伤等多种因素诱导并参与植物的信号传导[5], AtNAC019的表达 受ABA、热、高盐、JA、干旱应答等因素诱导, 并调节植物生长发育[6, 7, 8]。AtNAC055是拟南芥JA信号通路的关键基因, 调控JA的生物合成, 并能对高盐、真菌等多种逆境应答[7, 8]。AtNAC102能够参与拟南芥的洪涝胁迫应答[9]。OsNAC002[10, 11, 12, 13]、OsNAC048[14, 15]和OsNAC068[16]参与水稻的耐旱、耐盐等多种逆境胁迫。
随着对大豆NAC基因家族研究的深入, 越来越多的大豆NAC类转录因子得到了鉴定。孟庆长等首次在大豆中克隆得到6个GmNAC基因[17, 18]; 韩巧玲等[19]利用NTT (核蛋白筛选系统)从大豆耐盐品种铁丰8号中克隆得到GmNAC2a基因, 该基因对干旱、高温、低温、高盐、ABA、乙烯等多种途径均有响应; 金杭霞等[20]发现GmNAC2参与逆境调控, GmNAC5参与大豆发育调控; 才华等[21]利用酵母单杂交的方法从野生大豆中克隆得到能与耐逆相关顺式元件MYB1AT特异结合的GsNAC20基因, 该基因能够响应高盐、干旱和低温胁迫, 并且在根和叶中具有不同的表达模式, 超量表达GsNAC20的拟南芥提高盐胁迫的敏感性; Tran等[22]在大豆中克隆了31个GmNAC基因, 发现9个NAC基因能同时响应干旱、高盐、冷、ABA等胁迫应答, 聚类分析发现9个GmNAC基因与前人报道的逆境相关NAC基因位于同一分支; Le等[23, 24]利用大豆基因组数据库预测得到152个GmNAC基因, 发现58个基因可能参与逆境胁迫应答, 经定量分析发现25个和6个基因在干旱胁迫时分别上调和下调2倍以上; 王洋等[25]利用大豆基因组数据库预测得到152个GmNAC基因, 并将其划分为10个亚家族; Hao等[26, 27]研究发现, GmNAC11、GmNAC20通过调节DREB1A等相关基因增强植物高盐胁迫耐性, 进一步研究表明GmNAC20既可以调控生长素信号相关基因促进植株侧根形成, 又可以激活DREB/CBF-COR途径增强植物的高盐胁迫和冷胁迫耐性。
本研究在前期大豆品种耐盐性鉴定的基础上, 利用生物信息学手段挖掘大豆中逆境相关的GmNAC基因, 采用荧光定量PCR技术分析其在不同耐盐性大豆品种中的表达规律, 探索GmNAC基因对盐胁迫的应答, 克隆盐胁迫相关基因, 分析候选基因序列变异, 为抗逆转基因品种的培育提供基因材料及理论基础。
选用本课题组前期鉴定的耐盐品种徐豆10、齐黄34和盐敏感品种汾豆95。
从植物转录因子数据库PlantTFDB (http:// planttfdb.cbi.pku.edu.cn/)下载大豆NAC转录因子序列, 对去冗余后的蛋白序列根据Phytozome (http:// phytozome.org/)的功能注释并利用SMART (http:// smart.embl-heidelberg.de/)分析氨基酸结构域, 获得具有NAC保守结构域的候选基因; 查询文献获得拟南芥和水稻逆境胁迫相关的NAC蛋白质序列。
利用MEGA 5.05软件内置的Clustal W程序对获得的NAC蛋白进行序列比对; 根据多重序列比对结果, 使用MEGA 5.05软件采用最大似然法(maximum likelihood, ML)构建系统进化树。
选取与拟南芥和水稻的逆境胁迫相关NAC蛋白位于同一进化分支的大豆GmNAC蛋白, 查询文献或利用Primer 5设计引物进行荧光定量分析(表1)。将大豆种子播种于装有细沙的育苗盘中, 用1/4 MS培养液浇灌至大豆第一片三出复叶全展, TRIzol法提取大豆根部和叶片RNA, 利用Roche Lightcycler 480II进行荧光定量PCR, 利用2-Δ Δ Ct数据分析该基因在根叶中的表达情况; 将大豆种子播种于细沙的育苗盘中, 用1/4 MS培养液浇灌至大豆第1片三出复叶全展, 移至水培, 添加150 mmol L-1 NaCl的1/4 MS培养液处理, 1/4 MS培养液处理作为空白处理, 分别采集处理1、3、6和12 h的叶片和根提取RNA并定量分析表达情况。
| 表1 荧光定量引物序列 Table 1 Primer sequence for real-time PCR |
根据GmNAC的表达结果, 选取相关基因进行克隆。根据基因的5° UTR和3° UTR序列设计引物(表2), 分别以徐豆10、齐黄34和汾豆95的cDNA为模板进行基因克隆。利用DNAMAN对测序成功的基因进行序列比对, 分析变异位点。
| 表2 基因克隆引物序列 Table 2 Primer sequence for gene cloning |
从PlantTFDB下载大豆NAC转录因子序列247个, 去冗余后根据Phytozome的功能注释并利用SMART进行氨基酸结构域分析获得175条含有保守结构域的大豆GmNAC序列。通过文献检索(表3)分别获得拟南芥逆境相关NAC蛋白4条, 即AtNAC019、AtNAC055、AtNAC072和AtNAC102; 水稻逆境相关NAC蛋白3条, 即OsNAC002 (SNAC1)、OsNAC048 (OsNAC6)和OsNAC068 (OsNAC4)。将其与175条大豆GmNAC家族蛋白进行进化树分析(图1), 有11条大豆GmNAC蛋白与拟南芥和水稻逆境胁迫相关NAC蛋白处于同一分支, 分别为Glyma12g35000.1 (GmNAC092)、Glyma06g38410.1 (GmNAC043)、Glyma13g35550.1 (GmNAC101)、Glyma12g22880.1 (GmNAC085)、Glyma04g42800.1 (GmNAC022)、Glyma02g26480.1 (GmNAC011, GmNAC20)、Glyma14g24220.1 (GmNAC109)、Glyma06g16440.2、Glyma06g11970.1、Glyma04g 38560.1和Glyma05g32850.1, 其中7个基因GmNAC011、GmNAC022、GmNAC043、GmNAC085、GmNAC 092、GmNAC 101和GmN AC109受干旱胁迫诱导。
| 表3 水稻和拟南芥逆境相关NAC基因 Table 3 Stress related NAC genes in rice and Arabidopsis |
 | 图1 NAC蛋白系统进化树 ●: 大豆干旱胁迫诱导的GmNAC蛋白; ○: 水稻和拟南芥中逆境相关的NAC蛋白。Fig. 1 Phylogenetic tree of NAC proteins ●: GmNAC proteins induced by drought in soybean; ○: Stress related NAC proteins in rice and Arabidopsis. |
序列比对发现, 11个大豆NAC蛋白与拟南芥和水稻逆境相关的NAC蛋白结构域高度保守, 具有典型的NAC转录因子结构特征(图2)。AtNAC072、AtNAC019、AtNAC055、Glyma12g35000.1、Glyma 06g38410.1、Glyma13g35550.1、Glyma12g2288 0.1具有较高的同源率, AtNAC102、AtNAC002、Glyma04g42800.1、Glyma02g26480.1、Glyma14g242 20.1、Glyma06g16440.2、Glyma06g119 70.1、Glyma0 4g38560.1、Glyma05g32850.1具有较高的同源率, 这与基因的进化树分析的结果是一致的。
 | 图2 候选逆境相关NAC蛋白序列比对Fig. 2 Sequence alignment of candidate stress-related NAC proteins |
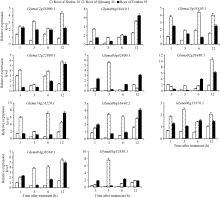
荧光定量分析GmNAC在大豆根部的表达规律(图3), 6个GmNAC基因Glyma12g35000.1、Glyma 06g38410.1、Glyma12g22880.1、Glyma06g16440.2、Glyma04g 38560.1、Glyma05g32850.1在耐盐品种徐豆10和齐黄34根部的表达量均高于盐敏感品种汾豆95; 2个GmNAC基因Glyma13g35550.1和Glyma14 g24220.1等在耐盐品种齐黄34根部的表达量高于盐敏感品种汾豆95; Glyma06g11970.1基因在耐盐品种徐豆10根部的表达量高于盐敏感品种汾豆95; Glyma04g42800.1、Glyma02g26480.1等2个GmNAC基因在3个品种根部的表达量差异不大。
 | 图3 GmNAC基因在根部的表达Fig. 3 Expression of GmNAC in root 1: Glyma12g35000.1; 2: Glyma06g38410.1; 3: Glyma13g35550.1; 4: Glyma12g22880.1; 5: Glyma04g42800.1; 6: Glyma02g26480.1; 7: Glyma14g24220.1; 8: Glyma06g16440.2; 9: Glyma06g11970.1; 10: Glyma04g38560.1; 11: Glyma05g32850.1. |
150 mmol L-1 NaCl水培处理徐豆10、齐黄34和汾豆95, 荧光定量分析GmNAC在其根部的表达量(图4)。GmNAC在根部的表达呈现品种间差异。在徐豆10根中, Glyma04g42800.1、Glyma06g164 40.2、Glyma06g11970.1、Glyma04g38560.1、Glyma 05g32850.1在NaCl处理1、3和6 h均下调表达, 但在12 h趋于稳定。其他6个基因在不同时刻小幅上调或下调表达, 未表现出明显规律。在齐黄34根中, 除Glyma04g42800.1外, GmNAC均不同程度的上调表达, 其中, Glyma06g16440.2、Glyma06g11970.1、Glyma05g32850.1在3 h达到上调高峰; Glyma 13g35550.1、Glyma14g24220.1在6 h达到上调高峰; Glyma12g35000.1、Glyma06g38410.1、Glyma12g22 880.1、Glyma02g264 80.1、Glym a04g38560.1在12 h达到上调高峰; 除3 h外, Glyma04g42800.1在其他取样时间均不同程度的下调表达。NaCl处理1 h和3 h时, GmNAC在汾豆95根中不同程度的上调或下调表达, 但在6 h和12 h时, 所有GmNAC基因均持续上调表达, 并在不同时间达到上调高峰。
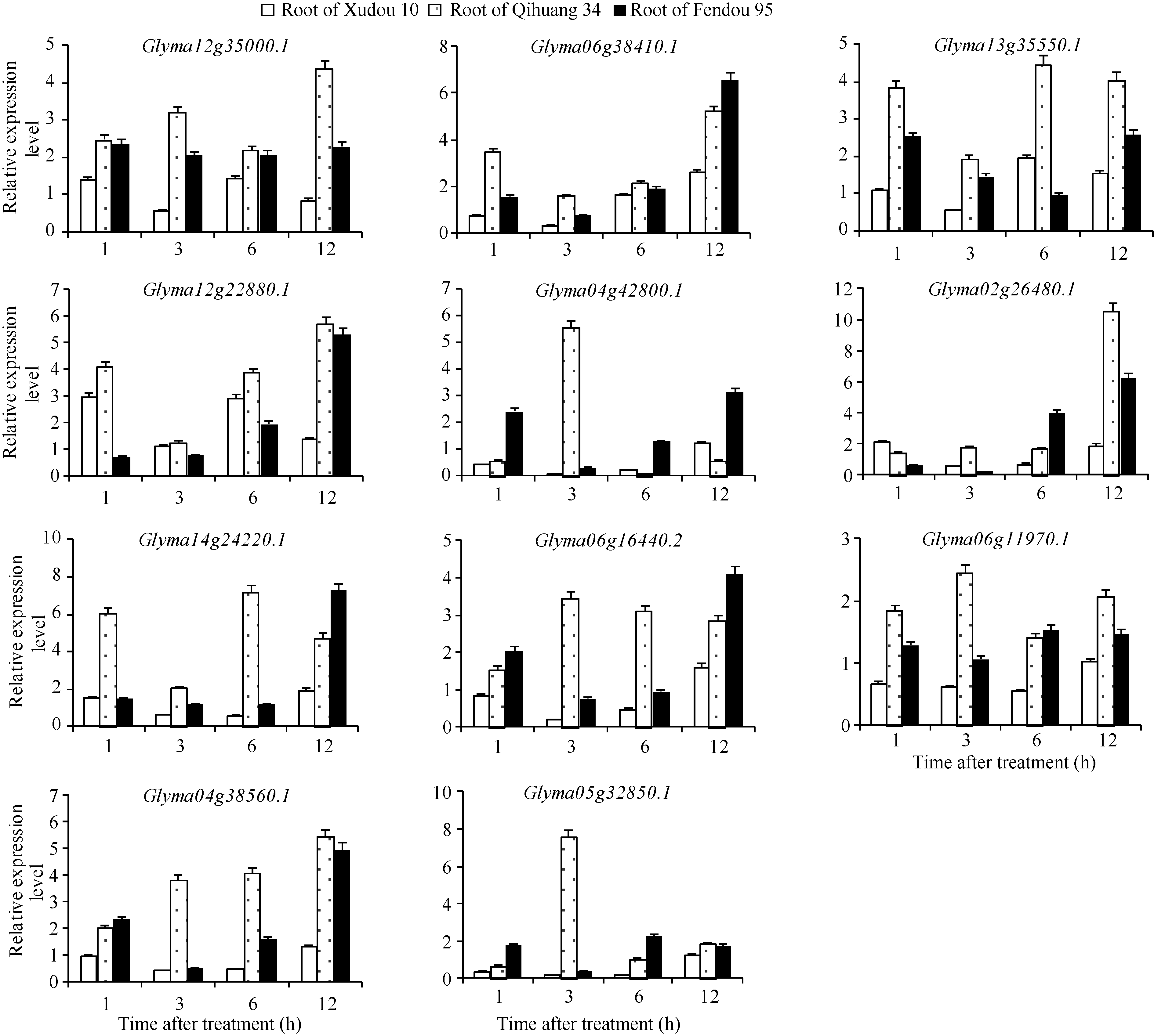 | 图4 NaCl处理后GmNAC基因的根部表达Fig. 4 Expression of GmNAC in root treated with NaCl |
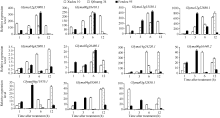
荧光定量分析GmNAC在大豆叶片中的表达规律(图5), 7个GmNAC基因Glyma12g22880.1、Glyma04g42800.1、Glyma14g24220.1、Glyma06g 16440.1、Glyma06g11970.1、Glyma04g38560.1、Glyma 05g32 850.1在耐盐品种徐豆10和齐黄34叶片的表达量均高于盐敏感品种汾豆95; Glyma02g26480.1基因在耐盐品种徐豆10叶片的表达量高于盐敏感品种汾豆95; 3个GmNAC基因Glyma12g35000.1、Glyma06g38410.1、Glyma13g355 50.1在耐盐品种徐豆10和齐黄34叶片的表达量均低于于盐敏感品种汾豆95。
150 mmol L-1 NaCl水培处理徐豆10、齐黄34和汾豆95, 荧光定量分析GmNAC在其叶片的表达量(图6)。几乎所有的GmNAC在大豆叶片中均高度上调表达。在徐豆10叶片中, Glyma12g35000.1、Glyma06g38410.1、Glyma13g35550.1、Glyma12g228 80.1、Glyma02g26480.1、Glyma14g24 220.1和Glyma06g16440.2上调表达, 并在6 h后达到高峰; Glyma04g42800.1在6 h达到上调高峰, 但在1 h和12 h均下调表达; Glyma06g11970.1、Glyma04g38 560.1和Glyma05g32850.1均在12 h达到上调高峰, 但在3 h下调表达。Glyma12g35000.1、Glyma06g38 410.1、Glyma13g35550.1和Glyma06g11970.1在齐黄34叶片中表达规律与徐豆10相同, 但表达倍数稍有不同; Glyma12g22880.1和Glyma14g24220.1在齐黄34叶片中上调表达, 并在12 h达到高峰; Glyma04g42800.1在1 h、3 h和6 h均下调, 但在12 h上调表达; Glyma02g26480.1在6 h达到上调高峰, 但在1 h下调表达; Glyma06g16440.2和Glyma06g11 970.1在12 h达到上调高峰, 但在3 h下调表达; Glyma04g38560.1在12 h达到上调高峰, 但在3 h和6 h均下调; Glyma05g32850.1在1 h达到上调高峰, 并在3 h开始下调之后持续上调表达。Glyma12g3 5000.1、Glyma06g38410.1、Glyma13g35550.1和Glyma02g26480.1在汾豆95叶片的表达规律和徐豆10相同, 但表达倍数稍有不同; Glyma04g42800.1在汾豆95中下调表达; Glyma12g22880.1、Glyma14g 24220.1、 Glyma06g16440.2、Glyma06g11970.1、Glyma04g385 60.1 和Gly ma05g 32850.1在汾豆95叶片中均上调表达并在3 h达到上调高峰。
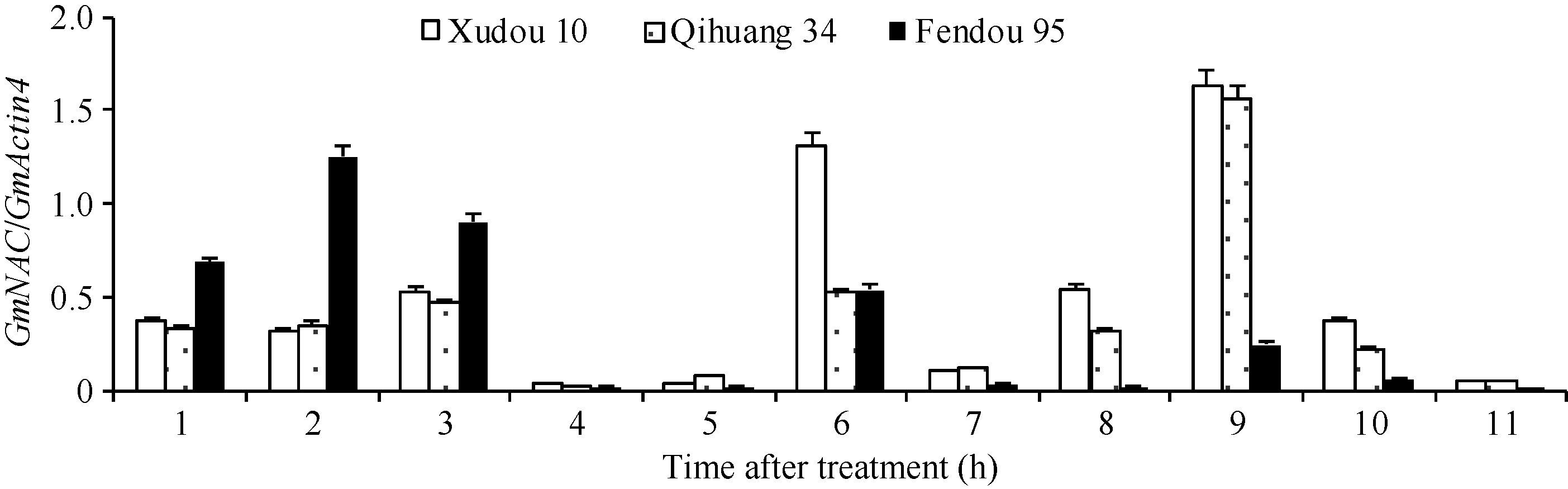 | 图5 GmNAC基因在叶片的表达Fig. 5 Expression of GmNAC in leaf 1: Glyma12g35000.1; 2: Glyma06g38410.1; 3: Glyma13g35550.1; 4: Glyma12g22880.1; 5: Glyma04g42800.1; 6: Glyma02g26480.1; 7: Glyma14g24220.1; 8: Glyma06g16440.2; 9: Glyma06g11970.1; 10: Glyma04g38560.1; 11: Glyma05g32850.1. |
 | 图6 NaCl处理后GmNAC基因的叶片表达Fig. 6 Expression of GmNAC in leaf treated with NaCl |
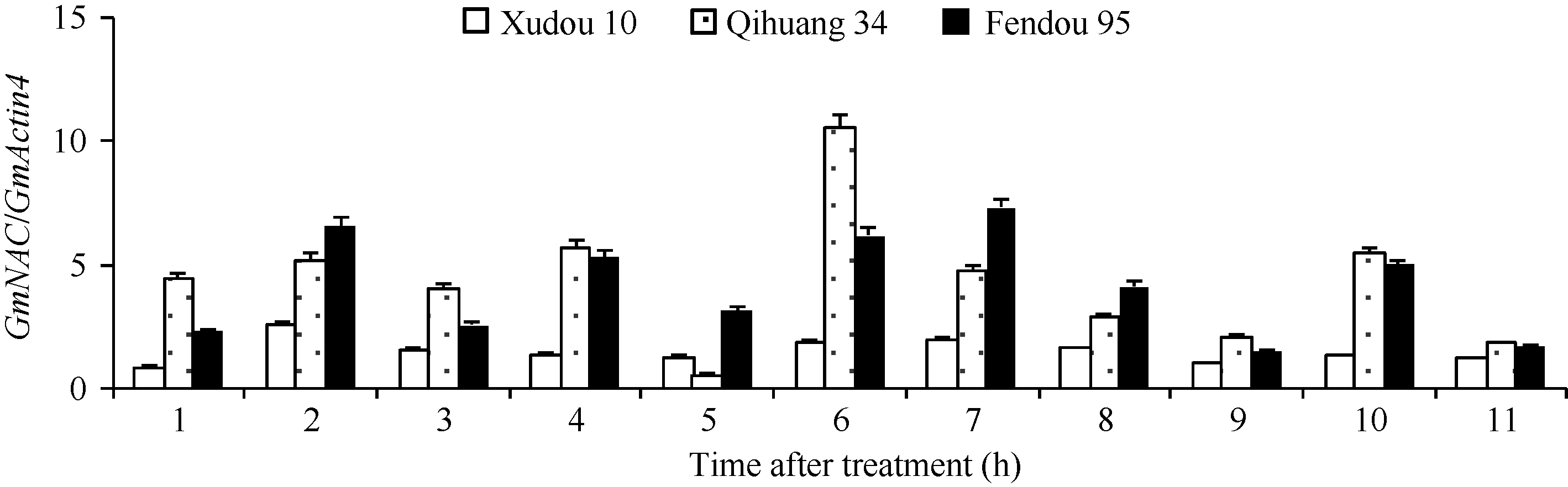
表达分析表明, 4个新发现的逆境候选GmNAC基因中, Glyma05g32850.1在大豆叶片和根部的表达量都很低, 虽然受NaCl胁迫诱导, 但在耐盐差异品种中未表现出特异的规律。选择其余3个表达差异显著的新基因(Glyma04g38560.1, Glyma06g11970.1和Glyma06g16440.2)进一步的克隆分析表明, Glyma04g38560.1在徐豆10、齐黄34、齐黄35中序列一致。Glyma06g11970.1 (图7-a)存在3个同义突变和1个非同义突变, 其中, 距起始密码子174 bp处发生同义突变, 突变类型为C/T; 距起始密码子444 bp处发生T/G的非同义突变, 导致所编码的氨基酸从天冬氨酸转变到谷氨酸, 但未引起蛋白质疏水性的变化; 距起始密码子591 bp处发生A/G的同义突变; 距起始密码子611 bp处发生G/C的同义突变。Glyma06g16440.2 (图7-b)在距起始密码子681 bp处存在1个同义突变, 突变类型为G/A。这些变异是否导致基因功能的变异还需进一步的功能分析。
 | 图7 GmNAC基因在大豆品种中的序列变异 a: Glyma06g11970.1在大豆品种中的序列变异; b: Glyma06g16440.2在大豆品种中的序列变异。Fig. 7 Sequence variations of GmNAC in soybean varieties a: Variations of Glyma06g11970.1 in soybean varieties; b: Variations of Glyma06g16440.2 in soybean varieties. |
高温、低温、干旱、盐碱等非生物逆境胁迫严重影响大豆的生长, 造成大豆的大幅度减产甚至绝产。NAC类转录因子在水稻、拟南芥等植物的生长发育、衰老、细胞分裂、逆境胁迫等途径中发挥着重要的作用。前人利用大豆基因组数据库获得152个GmNAC基因[23, 24, 25], 本研究利用植物转录因子数据库获得175个GmNAC基因, 这可能是基于的数据库不同造成的。李伟等[28]在分析22种NAC蛋白进行时发现非生物逆境相关的NAC蛋白聚成一类。You等[29]利用同源分析鉴定了18个可能参与非生物逆境应答的二穗短柄草NAC基因, 同时分析101个二穗短柄草的NAC基因在非生物逆境胁迫和逆境相关激素处理下的表达模式, 证实这18个基因可能在非生物逆境应答中发挥功能, 进一步证实基于进化树分析的功能预测在逆境相关NAC基因挖掘上的可行性。Tran等[22]在大豆中克隆了31个GmNAC基因, 发现9个NAC基因能同时响应干旱、高盐、冷、ABA等胁迫应答, 且与前人报道的逆境相关NAC基因聚成一类, 本研究经聚类分析发现11个大豆GmNAC基因与拟南芥和水稻中报道的逆境相关的NAC基因位于同一分支, 推测这11个NAC为大豆逆境相关基因。王洋等[25]将大豆NAC基因分为10个亚家族, 其中第6亚家族包含的11个基因与本研究推测的逆境相关候选基因相同。研究表明, 11个基因中, GmNAC011、GmNAC022、GmNAC043、GmNAC085、GmNAC092、GmNAC101和GmNAC109受干旱胁迫诱导[23, 24], 同时过表达GmNAC011的拟南芥能够提高盐胁迫和冷胁迫耐性[26, 27], 说明位于同一分支的11个候选基因可能均参与大豆逆境胁迫的应答。本研究同时发现, GmNAC11 (Glyma19g28476)等[26, 27]多个逆境诱导基因位于相邻分支, 可能暗示大豆中存在着更多的逆境相关NAC基因。Glyma12g13710.1等[23, 24]干旱诱导基因距离水稻和拟南芥的逆境相关NAC基因较远, 这些基因参与其他未知的逆境调控途径。
植物的逆境胁迫应答由多个复杂的网络通路共同调控, NAC受多种逆境、激素诱导。AtNAC019、AtNAC055、AtNAC072等基因参与高盐应答反应[5, 6, 7, 8], 其在大豆中的同源基因在本研究中均受NaCl诱导。大豆逆境相关候选GmNAC基因的表达在根部高于叶片, 这与前人的结果是一致的。NaCl胁迫下, 候选GmNAC在根部和叶片的表达模式不同, 这可能暗示大豆对盐胁迫应答存在组织特异性。Glyma12g35000.1、Glyma06g38410.1、Glyma13g35 550.1在耐盐品种齐黄34和徐豆10中表达量均低于在盐敏感品种汾豆95中, Glyma14g24220.1、Glyma06g16440.2、Glyma06g11970.1、Glyma04g3856 0.1和Glyma05g32850.1在耐盐品种齐黄34和徐豆10中表达量均高于在盐敏感品种汾豆95中, 这可能与大豆品种的耐盐性差异有关。Hao等[26, 27]研究表明, NaCl胁迫下GmNAC011在叶片中的表达量显著上升, 并在12 h达到表达高峰; 本研究表明, NaCl诱导下GmNAC011在叶片中的表达量显著上升, 但在6 h时达到表达高峰, 这可能与取样时间、大豆品种、NaCl处理浓度等多种因素有关, 本研究还发现, GmNAC011在大豆根部受NaCl的诱导程度弱于在叶片。NaCl胁迫下, 除Glyma12g22880.1外, 其他10个基因在叶片中的变化程度远大于在根部, 且同一个基因在根和叶片中表现出不同的表达规律, 这可能与基因参与的代谢通路有关。NaCl胁迫下, 数个基因在汾豆95的表达高峰比齐黄34和徐豆10中相对提前可能与品种的盐敏感程度有关。Glyma06g11970.1 和Glyma06g16440.2在齐黄34、徐豆10和汾豆95中存在序列差异, 这可能与品种的耐盐性有关, 这些变异位点的作用还需要进一步的功能分析。
获得了3个盐胁迫相关的GmNAC基因(Glyma04g38560.1、Glyma06g11970.1和Glyma0 6g16440.2), 其中Glyma06g11970.1和Glyma0 6g16440.2在徐豆10、齐黄34和汾豆95中存在SNP位点。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|