
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
基于CRISPR/Cas9技术的水稻千粒重基因 tgw6突变体的创建
[王加峰 , 郑才敏, 刘维, 罗文龙, 王慧, 陈志强
, 郑才敏, 刘维, 罗文龙, 王慧, 陈志强* , 郭涛* ]
, 郑才敏, 刘维, 罗文龙, 王慧, 陈志强, 郭涛]
|
|


* 通讯作者(Corresponding authors): 郭涛, E-mail: guoguot@scau.edu.cn, Tel: 020-38604903; 陈志强, E-mail:chenlin@scau.edu.cn , Tel:020-85283237
第一作者联系方式: E-mail:bcjfwang@gmail.com, Tel: 020-38604903
利用CRISPR/Cas9技术对调控水稻产量千粒重基因 TGW6定点编辑, 获得了一套有重要育种价值的 tgw6突变体。设计了分别由U3、U6a和U6b启动子驱动、长20 bp的guide RNA (gRNA)靶点以靶向编辑 TGW6基因的外显子, 首先将这3个靶点一起组装到pYLCRISPR/Cas9-MT(I)载体上, 然后利用农杆菌介导侵染水稻材料H447 (R819/玉针香//R819的BC3F6); 提取T0代转基因植株的基因组DNA并对编辑位点附近的DNA片段进行PCR检测及测序分析。结果表明, T0代材料中 tgw6的突变频率高达90%, 其中纯合缺失突变率约占51%。对T1代纯合缺失突变体的千粒重性状的调查分析结果表明, 部分 tgw6的缺失突变能显著提高千粒重(大于5%)。不同类型 tgw6突变体的成功创建不仅丰富了 tgw6的变异类型, 为水稻的高产稳产奠定了重要的材料基础, 还证实了CRISPR/Cas9技术在水稻基因工程育种中高效、易操作的特点。
A set of tgw6 ( Thousand-grain weight 6) mutants were constructed using CRISPR/Cas9 technology in this study. Three sites of 20 nt guide RNA (gRNA) targeted to the exon of TGW6were designed and transcribed from the U3, U6a, or U6b promoters, respectively. The three target sites of gRNA were then ligated to the vector pYLCRISPR/Cas9-MT(I) based on golden gate cloning strategy. The recombinant plasmid was transferred to a rice cultivar, H447 (R819/Yuzhenxiang//R819 BC3F6) by Agrobacterium-mediated transformation. Sequencing for the genomic DNA of TGW6locus in T0 rice showed the mutagenesis frequency for TGW6was more than 90%, including 51% of homozygous deletion mutations. Further analysis for the T1 mutants showed that almost all the homozygous deletion mutants improved the thousand-grain weight significantly (more than 5%). The successful tgw6 editing not only provided a series of tgw6mutants for high and stable yield of rice but also proved that CRISPR/Cas9 is a facile and powerful means of rice genetic engineering for scientific and agricultural applications, which has important theoretical and practical significance for rice breeding.
水稻(Oryza sativaL.)是世界上重要的粮食作物之一。由于人口的不断增长, 人们对粮食的需求也越来越大, 但水稻的单位面积产量一直没有太大提升, 而且水稻的种植面积也在持续下降, 严重威胁着我国的粮食安全。要解决这一问题, 必须借助新技术和新的遗传改良策略。粒重是水稻产量构成的三因子之一, 主要决定于籽粒胚乳中淀粉的合成与积累和籽粒的大小, 表现为典型的数量性状, 适当提高千粒重, 有利于产量的提高[1]。目前, 已经克隆的千粒重相关基因有qSW5/GW5[2, 3]、TGW6[4]、GS3[5]、GS5[6]、GW2[7]、GW8[8]、qGL3/qGL3-1/GL3.1[9, 10, 11]等。其中, TGW6 (Thousand-Grain Weight 6)是调控水稻千粒重性状的最重要的基因之一。TGW6基因编码吲哚乙酸-葡糖糖水解酶, 其功能缺失突变(313 bp)会引起胚乳中吲哚乙酸含量下降, 进而细胞数量增加、粒长和粒重增加, 使水稻抽穗前籽粒的碳水化合物积累, 使日本晴产量增加15%, 且不影响稻米品质[4]。
CRISPR/Cas9系统是近几年发展的一种准确、便捷、高效率的生物基因组编辑方法。该系统仅需要短的gRNA和核酸酶(Cas9)就可以使特定的生物靶基因定点突变, 为生物定点编辑技术的发展注入了新的活力[12, 13, 14]。目前, CRISPR/Cas9系统不但在酵母、果蝇、鼠、人等生物中, 而且已成功在拟南芥、烟草、甜橙、水稻、小麦、高粱、玉米以及苔藓植物地钱等植物中实现了定点基因组编辑[15, 16, 17, 18, 19], 但对水稻育种中有重要价值的产量、品质、育性等关键基因的定向编辑研究鲜有报道, 更缺乏对相关突变体的育种价值评价。本研究利用CRISPR/Cas9技术定点编辑了调控水稻千粒重的TGW6基因, 获得了一套具有重要应用价值的水稻tgw6突变体新种质, 为进一步提高水稻产量奠定了重要的材料基础。
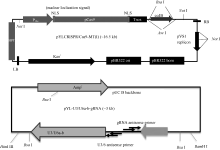
水稻材料H447为籼稻恢复系R819与优质米常规籼稻品种玉针香杂交后, 以R819为轮回亲本回交、自交得到的BC3F6代品系。该品系具有抗稻瘟病、株型紧凑、米质优等特点。Cas9载体pYLCRISPR/ Cas9-MT(I)及gRNA载体pYL-U3/U6a-b-gRNAs (图1)由华南农业大学刘耀光研究员馈赠。
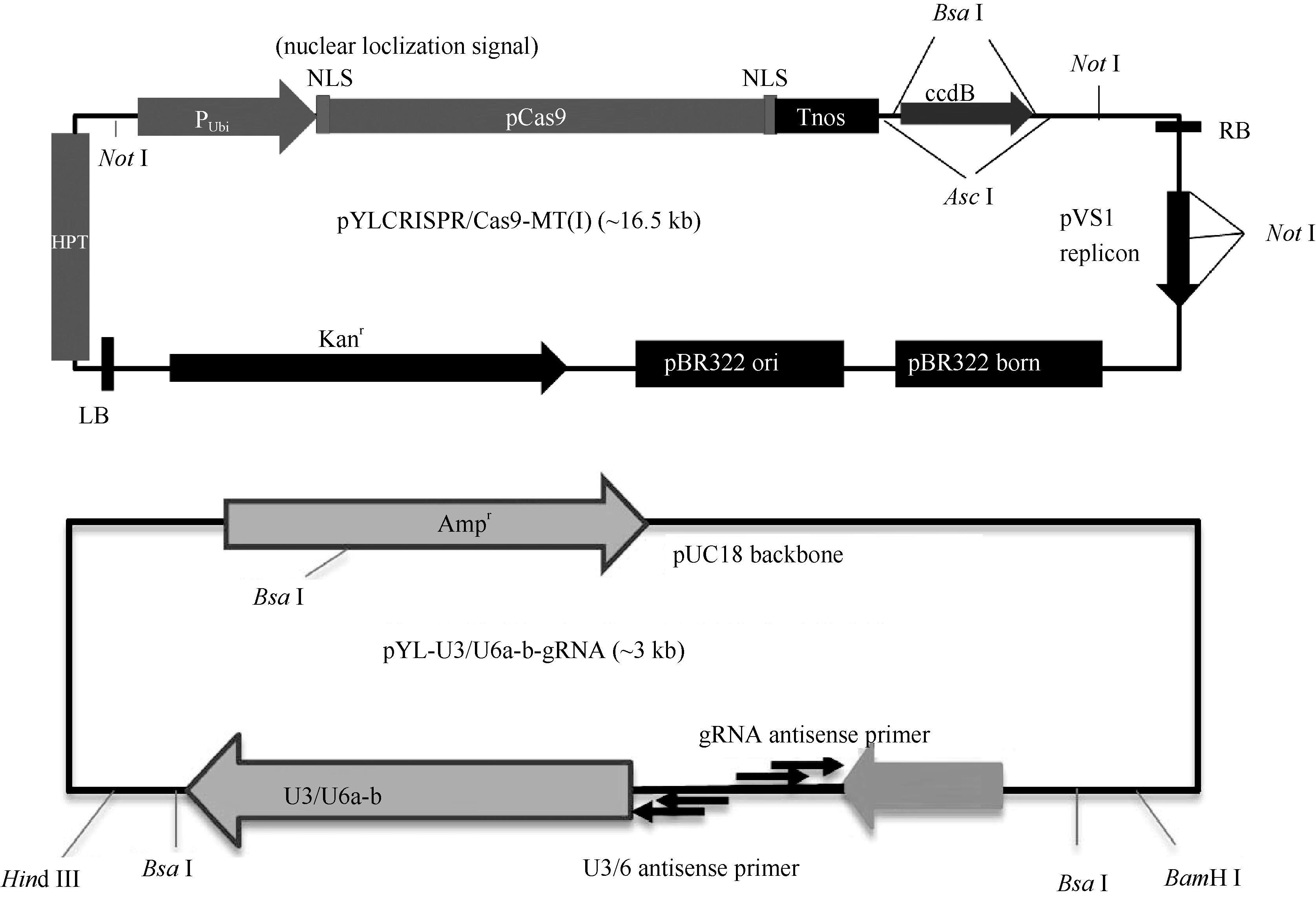 | 图1 YLCRISPR/Cas9-MT(I)及pYL-U3/U6a-b-gRNA质粒图Fig. 1 Maps of YLCRISPR/Cas9-MT(I) and pYL-U3/U6a-b-gRNA vectors |
根据调控水稻TGW6的外显子序列(GenBank登录号为AB513135.1), 设计TGW6的gRNA。按A/G(N) 20NGG序列设计20 nt的寡核苷酸gRNA核心序列, 在水稻基因组数据库中比对设计好的gRNA序列以排除非特异性的靶切位点。寡核苷酸序列见表1。同时设计CRISPR鉴定引物Cas9-TGW6testF: 5° -CAAC CAAACCAAAGCCTGC-3° 和Cas9-TGW6testR: 5° -C CAATGCCTCATCAACTTAC-3° 。所有寡核苷酸链引物均由Thermo Fisher Scientific公司合成。
| 表1 gRNA靶点以及寡核苷酸序列 Table 1 Target sites of the gRNA and the oligonucleotide sequences |
参照Ma等[18]的方法略作修改。(1)制备双链接头。分别取等量每个靶点的1对gRNA寡核苷酸链的上游引物与下游引物混合(终浓度1 µ mol L-1), 95℃处理30 s, 然后移至室温冷却完成退火; (2)酶切。取pYL-U3-gRNA、pYL-U6a-gRNA及pYL-U6b-gRNA质粒各1 μ g, 制备20 μ L反应体系, 用5~10 UBsa I (NEB)酶切20 min, 70℃处理10 min使酶失活; (3)连接和PCR扩增: 分别将酶切过的pYL-U3-gRNA、pYL-U6a-gRNA及pYL-U6b-gRNA质粒与各自所对应双链接头连接, 然后分别利用引物B1’ /B2 (B1’ : 5° -TTCAGAGGTCTCTCTCGCACTGGAATCGGCA GCAAAGG-3° ; B2: 5° -AGCGTGGGTCTCGTCAGG GTCCATCCACTCCAAGCTC-3° )、B2’ /B3 (B2’ : 5° - TTCAGAGGTCTCTCTGACACTGGAATCGGCAGCAAAGG-3° ; B3: 5° -AGCGTGGTCTCGTCTTGGTC CATCCACTCCAAGCTC-3° )、B3’ /BL (B3’ : 5° -TTCA GAGGTCTCTAAGACACTGGAATCGGCAGCAAA GG-3° ; BL: 5° -AGCGTGGGTCTCGACCGGGTCCA TCCACTCCAAGCTC-3° )对gDNA (gRNA对应的DNA序列)进行扩增, 先95℃ 1 min, 然后按95℃ 15 s、60℃ 15 s和68℃ 30 s进行30个循环的反应; (4)产物纯化、酶切和连接。将所有靶点gDNA PCR产物回收混合后用20 U Bsa I在37℃酶切30 min, 然后75℃处理5 min; 将酶切片段纯化后与Bsa I酶切回收的pYLCRISPR/Cas9-MT(I)载体片段利用T4 DNA ligase (NEB) 20℃连接2 h; (5)转化及质粒测序。连接产物转化DH5α 感受态细胞后, 挑取单克隆接种培养, 抽提质粒后利用Asc I进行酶切鉴定, 并挑取酶切(Asc I)验证正确的克隆送Thermo Fisher Scientific公司测序。
参照Hiei等[20]的方法, 将构建好的三靶点pYLCRISPR/Cas9-tgw6-gRNA载体通过电击转化到农杆菌EHA105中, 将PCR检测呈阳性的克隆用于水稻愈伤组织转化。
采用CTAB法提取转基因水稻基因组DNA[21]。PCR扩增体系含2.5 μ L 10× buffer for KOD-Plus、0.5 μ L KOD plus聚合酶(5 U μ L-1)、1 μ L 25 mmol L-1 MgSO4、2.5 μ L 2 mmol L-1 dNTPs、各0.5 μ L引物(10 μ mol L-1, Cas9-TGW6testF与Cas9-TGW6testR)、0.5 μ L模板DNA, 以超纯水补至25 μ L。反应条件为94℃ 3 min; 32个循环的94℃ 30 s、55℃ 30 s和68℃ 60 s的反应, 最后68℃延伸5 min。扩增产物经1%琼脂糖凝胶电泳(电泳缓冲液1× TAE), BIORAD凝胶成像系统观察、照相, 并送Thermo Fisher Scientific公司测序, 每个样品测序2次。
将PCR测序鉴定为突变的T0代转基因单株自交结实后得到T1代种子。T1代种子播种成苗后提取叶片基因组DNA进行PCR检测, 对经鉴定tgw6发生了纯合突变的个体进一步利用引物hptF: 5° -AAGCTG CATCATCGAAATTGC-3° 与hptR: 5° -AAGAATCTC GTGCTTTCAGCTTCG-3° 进行PCR检测以获得不含T-DNA成分的纯合tgw6突变株系。
采用随机区组设计种植不含T-DNA成分的纯合tgw6突变株系及H447, 2个重复, 各小区中每个株系种植6行, 每行6株。株行距为20 cm × 20 cm, 单本栽插。按大田常规栽培要求实施田间管理(水、肥、病虫害防治等)。收获成熟种子烘干后从中随机数取饱满、无病种子1000粒, 称重(g), 2个重复。保证重复间的差数与平均数之比< 5%。采用SPSS13.0计算平均值和标准误, 采用单因素方差分析(One-Way ANOVA)及Duncan法多重比较, 差异显著性水平为P≤ 0.05。
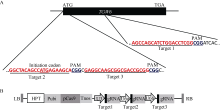
为利用CRISPR/Cas9技术获得tgw6的突变体, 根据CRISPR/Cas9技术的原理, 利用载体pYLCRISPR/Cas9-MT(I)及pYL-U3/U6a-b-gRNA构建基因编辑载体。设计了3个靶点以定点编辑千粒重基因TGW6 (表1), 分别从起始密码子附近开始设计这3个靶点(图2-A), 以保证可以彻底破坏TGW6的功能, 然后利用Golden gate的克隆方法[18]将这3个靶点gRNA组装到pYLCRISPR/Cas9-MT(I)载体上(图2-B)。
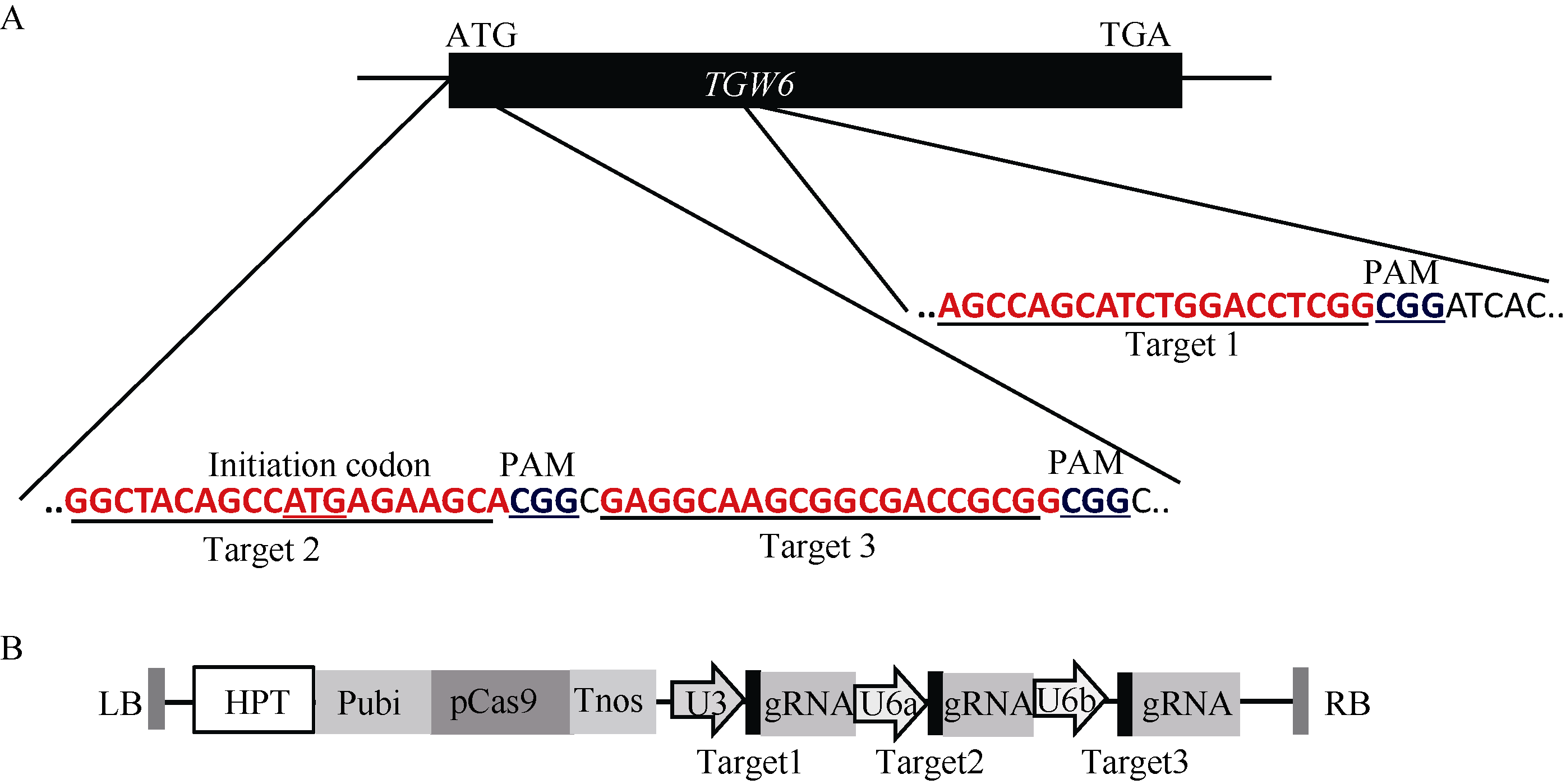 | 图2 gRNA靶位点及CRISPR/Cas9-gRNA的组装示意图 A: 3个靶点分别在TGW6基因内的位置; B: 3个靶点组装到pYLCRISPR/Cas9-MT(I)载体而成的T-DNA元件。Fig. 2 Target sites of the gRNA and cloning of gRNA cassette into the CRISPR/Cas9 vector A: positions of three targets in the TGW6 gene locus; B: T-DNA fragment assembled with the three targets and the pYLCRISPR/Cas9-MT(I) vector. |
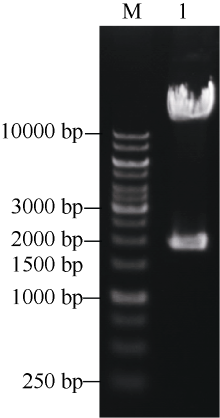
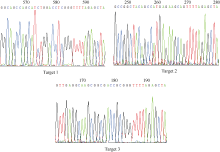
将构建的三靶点pYLCRISPR/Cas9-tgw6-gRNA载体利用Asc I酶切, 如图3所示, 3个靶点gDNA序列能被同时切出(1.8 kb左右), 表明三靶点gRNA元件顺利插入到pYLCRISPR/Cas9-MT(I)载体上。为进一步确定靶位点的准确性, 利用引物U3s (5° -GCATG GATCTTGGAGGAATCAGA-3° )、U6as (5° -GGCTA TCGAGATGCCATACA-3° )及U6bs (5° -AGAGAAGC CTAGTGTGCTCT-3° )分别对各个靶点(Target 1~3)测序分析。结果表明(图4), 通过Bsa I位点组装的3个靶点序列都与所设计靶点序列(表1)一致, 因此所构建的pYLCRISPR/Cas9-tgw6-gRNA载体适宜于下一步农杆菌介导的水稻遗传转化。
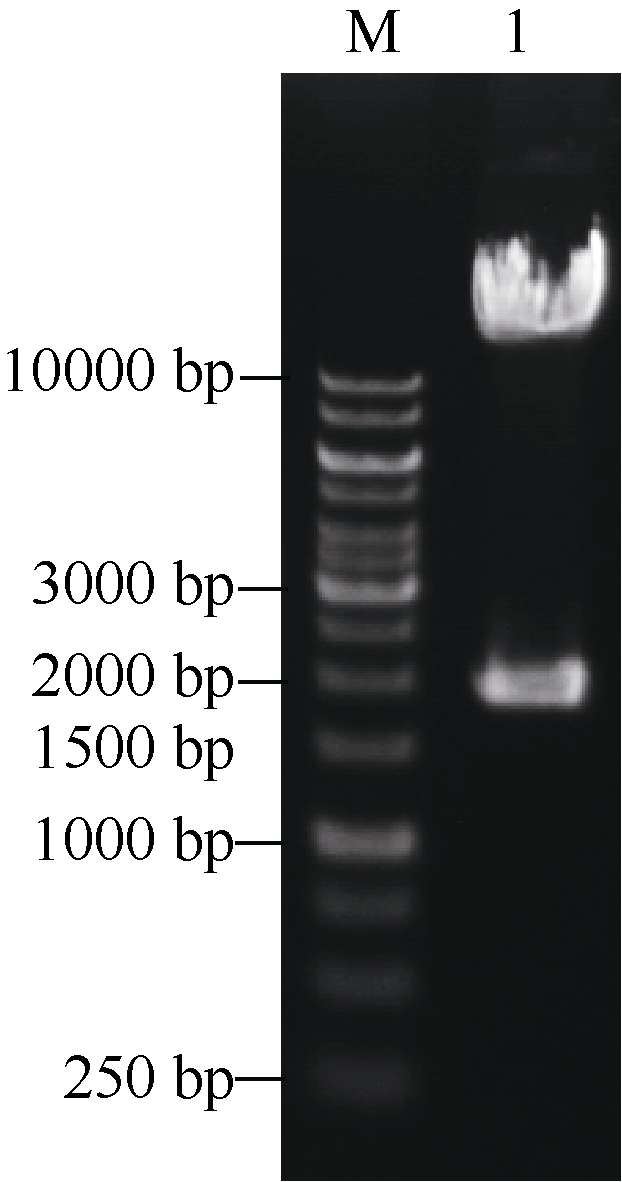 | 图3 AscI酶切鉴定pYLCRISPR/Cas9-tgw6-gRNA载体Fig. 3 Identification of the pYLCRISPR/Cas9-tgw6-gRNA plasmid digested with Asc I M: 1 kb DNA ladder marker; 1: pYLCRISPR/Cas9-tgw6-gRNA. |
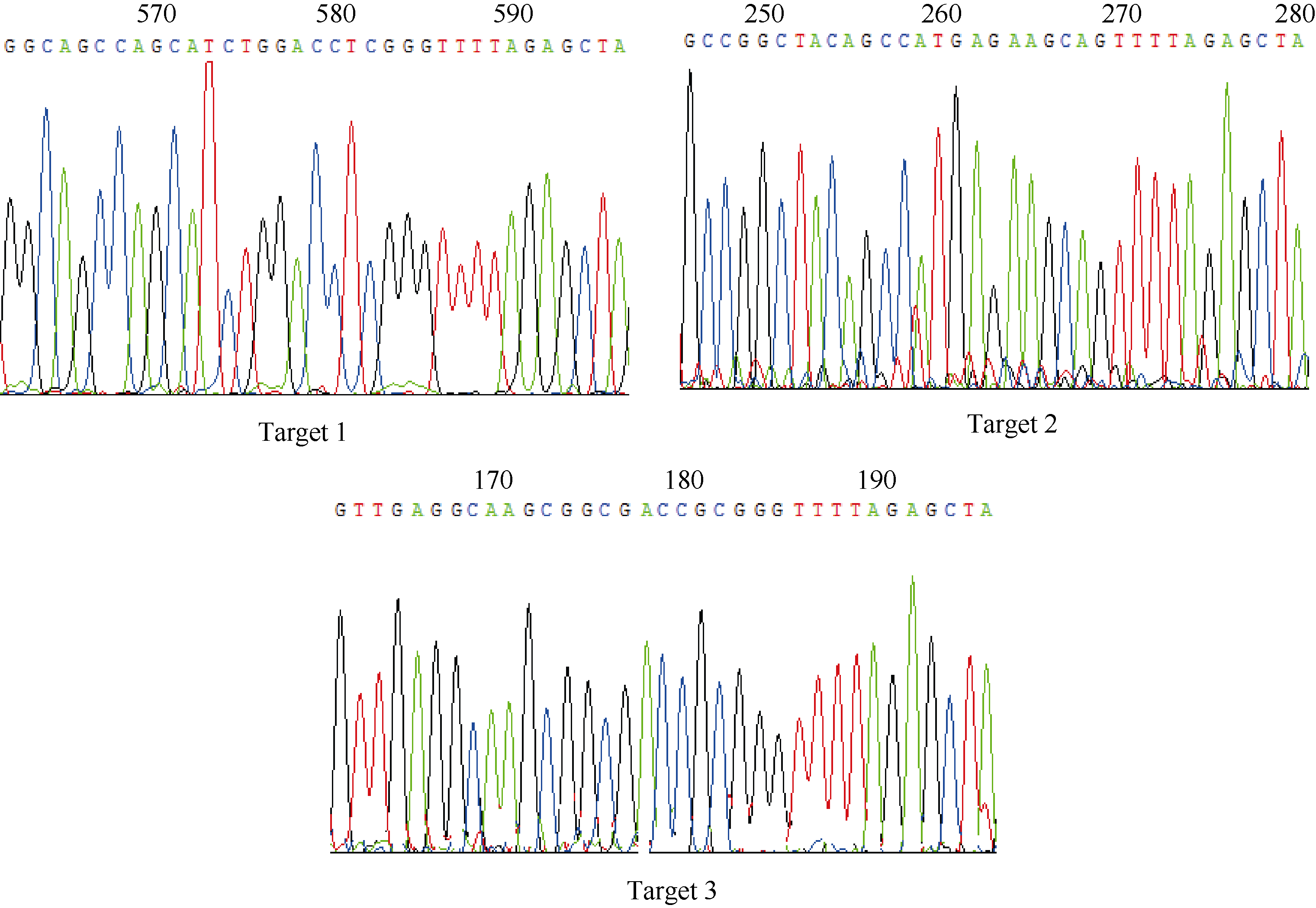 | 图4 3个靶点序列的测序结果Fig. 4 Sequencing results for the three target sequences |
采用CTAB法提取T0代再生水稻植株的叶片, 对潮霉素基因检测为阳性的植株利用引物Cas9- TGW6testF及Cas9-TGW6testR对tgw6编辑位点上下游200 bp左右区域DNA片段进行PCR扩增。电泳结果(图5-A)表明, 扩增产物中多为tgw6片段缺失纯合体, 也有部分双等位杂合突变体存在。进一步对扩增产物的测序分析表明(图5-B), tgw6突变频率高达90%, 其中51%为片段缺失纯合体(缺失片段长度均在100 bp以上), 39.5%是双等位杂合突变体。
 | 图5 tgw6突变体的PCR检测及其与野生型序列比对分析 A: T0代(1~22)水稻tgw6编辑位点附近DNA片段的PCR检测结果, 野生型(WT)为H447, 水稻扩增长度为953 bp; B: 对应于A的PCR产物的测得序列与野生型(WT)的序列比对结果。“ Frequency” 指A电泳图对应的突变体中同一类型编辑位点突变个体出现的频率。Fig. 5 PCR identification and sequence alignment of tgw6 mutants compared to the WT line A: PCR identification results for the DNA fragments near the edited locus of thetgw6 T0 mutants (1-22) and WT line (H447); B: sequence alignment of tgw6 mutants compared to the WT line. “ Frequency” refers to the frequency of the same edited mutation among the corresponding mutants in the electrophoresis map of A. |
为进一步获得无T-DNA插入元件的tgw6突变体, 将T1代tgw6缺失纯合突变的种子播种成苗, 苗期利用引物hphF与hphR对潮霉素基因进行PCR检测。结果表明, T1代转基因苗中潮霉素基因的分离符合孟德尔定律(数据未列出), 图6中列出了对部分tgw6突变体潮霉素基因PCR检测的结果, 以进一步对这些材料进行千粒重性状的分析。对T2代种子千粒重调查结果表明(表2), 所分析的5种类型的纯合缺失突变都显著提高了水稻的千粒重(大于5%)。
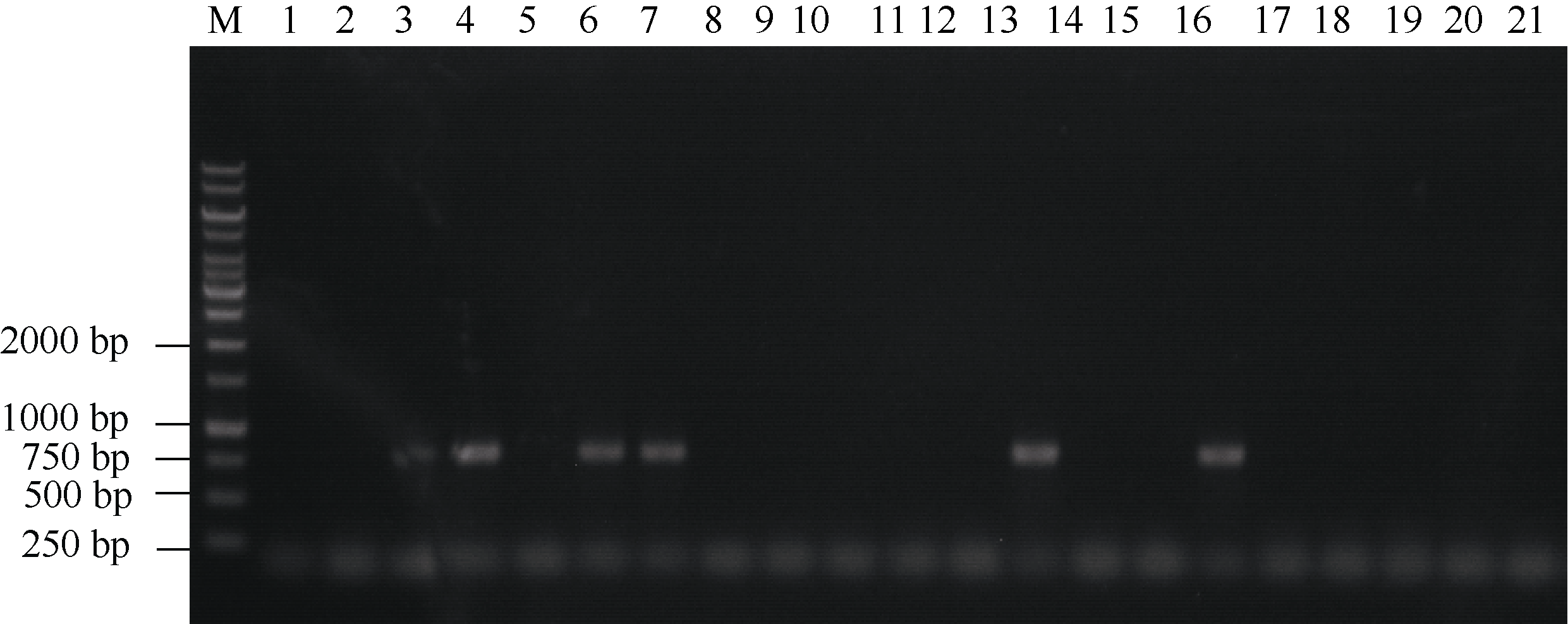 | 图6 部分tgw6突变体潮霉素基因的PCR检测 1: WT; 2~5: 缺失103 bp的突变体; 6~9: 缺失123 bp的突变体; 10~13: 缺失141 bp的突变体; 14~17: 缺失144 bp的突变体; 18~21: 缺失148 bp的突变体。Fig. 6 PCR identification for hph gene of parts of tgw6 mutants 1: WT; 2-5: the 103-bp deletion-mutants; 6-9: the 123-bp deletion-mutants; 10-13: the 141-bp deletion-mutants; 14-17: the 144-bp deletion-mutants; 18-21: the 148-bp deletion-mutants. |
| 表2 不同类型tgw6缺失突变体的千粒重测定结果 Table 2 Thousand grain weight of different types of homozygous deletion mutants of tgw6 |
CRISPR/Cas9是一种新兴的基因定点编辑技术, 具有较高的特异性和编辑效率。相对于锌指核酸酶(zinc-finger nucleases, ZFNs)[22, 23, 24]及TALE核酸酶(transcription activator like effector nucleases, TALENs)[25, 26, 27]2种基因编辑技术, CRISPR/Cas9技术构建简便灵活, 仅需要一个gRNA及一个核酸酶(Cas9)即可实现对靶基因DNA序列的剪切。目前的研究主要集中于CRISPR/Cas9技术的开发及特定基因敲除。
本研究中利用多靶点gRNAs对水稻的TGW6基因进行一次性定点编辑, 获得了一系列不同类型的tgw6突变体, T0代tgw6的突变频率高达90%, 其中, tgw6纯合缺失突变比例约占51%, 表明多靶点的定点编辑利于在T0代获得纯合缺失突变的类型。对T1代的幼苗所含潮霉素基因的PCR检测结果与大多农杆菌介导的水稻遗传转化一致, 潮霉素基因的分离也符合孟德尔定律, 因此通过转基因T0代植株自交便可以在T1代分离出无转基因成分的tgw6纯合缺失突变株系。虽然Xu等[19]指出, T1代转基因植株的突变将具有不确定性, 到T2代以后该突变才能稳定遗传, 这主要是由基因编辑时选用的靶点过少(通常为1个)造成的, 单靶点很难造成大片段的缺失, 多是碱基的替换或者插入等突变, 使得T1代转基因植株靶基因的编辑位点还可能存在gRNA靶点识别的序列, 导致发生再次编辑, 从而使得突变具有不确定性; 相反, 利用2个以上靶点进行的基因定点编辑可以提高纯合缺失突变的频率, 在T0代就可能获得可以稳定遗传给T1代的突变。虽然单靶点的基因编辑也会出现纯合突变的变异类型, 但是突变率大多在20%以下[19], 而且难以利用PCR鉴定出来, 一般需要借助对变异位点附近的DNA片段测序才能鉴定出来。另一方面, 由于gRNA识别序列较短(20 bp左右), 靶点多造成脱靶的可能性越大[28], 水稻等植物可以通过回交等手段来降低脱靶的影响。在利用CRISPR/Cas9技术进行水稻等植物相关性状突变体的创建过程中, 需要综合考虑合理选择转化受体及靶点位置和数量, 如本研究中对突变体千粒重性状分析发现, 突变体的千粒重都比对照提高了5%以上, 但与已知的tgw6突变体(313 bp缺失)会使日本晴产量增加15%[9]的结果不太一致, 推测可能是材料背景差异或者突变位点的差异引起的。
随着人类对粮食的高产、高品质需求的不断提高, 仅对基因进行定向改良是远远不够的, 如何对特定育种材料进行多基因的一次性导入成为亟待解决的问题。随着研究的不断深入, 人们已经发现利用CRISPR/Cas9剪切形成的缺口可促进外源基因片段高效重组到基因组中, 这一发现已先后在细菌、酵母、老鼠、果蝇、斑马鱼、线虫上取得了成功[29, 30, 31, 32, 33, 34, 35]。由于植物的特殊性, 使得依赖于CRISPR/Cas9的基因导入技术发展相对较慢, 即使如此, 有研究小组已经利用双生病毒载体系统在烟草、番茄中取得了成功[36, 37]。相信在1~2年内该技术也会在水稻中被成功应用, 这一技术的发展将会对水稻品种抗性、品质等改良起到重大推动作用, 具有广阔的应用前景。
获得了类型较为丰富(多碱基缺失、插入等)的tgw6突变体, 为进一步应用于水稻的高产、稳产育种奠定了重要材料基础。为快速创建稻米品质(fgr、Chalk)、雄性不育(tms5)等生产上有重要应用价值的水稻优异新种质提供了重要参考, 并有望为水稻种质资源创新提供安全、高效的新途径, 具有重要的理论和实践意义。
致谢: 感谢华南农业大学刘耀光研究员提供的pYLCRISPR/Cas9-MT(I)及gRNA载体(pYL-U3/U6a- b-gRNAs)。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|