




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
一个新的水稻 D1基因等位突变体的遗传鉴定与基因功能分析
[王翠红, 马建, 王帅, 田鹏, 岂长燕, 赵志超, 王久林, 王洁, 程治军, 张欣, 郭秀平, 雷财林*  ]
]
]
|
|


第一作者联系方式: E-mail: waangch@sina.com
株高是影响水稻产量的一个重要性状。本研究从水稻稻瘟病普感品种丽江新团黑谷(LTH)经甲基磺酸乙酯(EMS)诱变群体中分离出一个遗传稳定的小粒矮化突变体 LTH-m3。该突变体是赤霉素(GA)和油菜素内酯(BR)相关突变体, 它对外源GA (GA3)不敏感, 对外源BR (eBL)的敏感性较野生型显著降低。遗传分析、基因克隆和转基因互补实验确认, 该突变体是一个新的 d1基因等位突变体, 其 D1基因在第6个外显子与内含子接合处发生单碱基突变(G2522→A2522), 导致第6外显子被选择性剪切及Gα蛋白翻译提前终止, 从而造成 LTH-m3小粒矮化突变表型。进一步的研究表明, 该突变体 D1基因突变引起 SD1和 SLR1等基因表达的显著改变, 因而影响植株细胞内GA和BR反馈调节功能和信号传递。突变体 LTH-m3弥补了LTH植株过高、茎秆软和极易倒伏等缺陷, 可作为LTH的改良系在今后水稻稻瘟病研究中加以利用, 其功能突变基因的鉴定为深入研究水稻Gα蛋白的功能及激素信号途径提供了新的材料。
Plant height is one of important traits for rice yield. One genetically stable rice mutant, LTH-m3, was isolated from the cv. Lijiangxintuanheigu (LTH)-derived mutant population by mutagenesis using ethylmethane sulfonate (EMS). LTH-m3 was involved in the pathways of gibberellic acid (GA) and brassinosteroid (BR), and showed no sensitiveness to exogenous GA (GA3) and significantly reduced sensitiveness to exogenous BR (eBL) compared with the wild type. The genetic analysis, gene cloning and transgenic complementary test confirmed that LTH-m3 was a new d1-allelic mutant with small grain and dwarf phenotypes, and a single base was mutated (G2522→A2522) in the functional dwarf gene D1 at the conjunction site of its sixth exon and intron, which caused excision of the sixth exon in mRNA and premature termination of the D1encoded Gα protein, resulting in mutated phenotypes in the mutant. The further study showed that the D1 mutation caused obvious expression change of some dwarf genes such as SD1 and SLR1 in the mutant, and could affect the GA and BR pathways in their feedback regulations and signaling transductions in plant cells. The mutant overcome the defects of the universally blast-susceptible cv. LTH, such as too tall plant, soft stem and easy lodging, and could be utilized as an improved substitute of LTH in the future rice blast researches. The mutated D1 gene identified from the LTH-m3mutant may be useful for further study of Gα functions and signaling pathways of GA and BR.
株高是水稻株型的重要组成因子, 直接影响水稻的抗倒伏能力和产量。20世纪60年代由sd1引领的水稻第一次“ 绿色革命” , 即通过半矮化育种以提高抗倒性而实现的[1]。近年来育种家相继提出水稻理想株型的概念, 适当的株高依然是其核心内容之一[2, 3, 4, 5]。目前我国乃至全球优良水稻矮秆资源匮乏, 生产上应用的品种普遍携带sd1[6, 7], 潜伏着遗传脆弱性的风险[8]。发掘新的矮化突变体, 深入研究水稻植株矮化基因及株高调控机制, 将有力推动实现可利用矮秆基因的多样化及水稻育种的新突破。
目前已报道80多个水稻矮化突变体, 它们被划分为半矮秆、多蘖矮秆、小粒矮秆、畸形矮秆等类型[9, 10](http://shigen.lab.nig.ac.jp/rice/oryzabase/genes)。已克隆近30个矮化基因, 它们大多参与赤霉素(gibberellin, GA)合成或信号传导途径[11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27], 也有部分参与油菜素内酯(brassinosteroid, BR)[19, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37]或独脚金内酯(strigolactone, SL)[38, 39, 40, 41, 42, 43, 44, 45, 46]途径。多种激素的参与使得水稻株高调控机制错综复杂。
水稻小粒矮化突变体通常与GA和BR有关[10, 28, 47]。d1是发现最早的水稻小粒矮化突变体, 其D1基因发生碱基缺失突变, 不能编码有功能的异源三聚体GTP结合蛋白α 亚基(Gα ), 导致突变体植株矮小, 叶型紧凑, 谷粒小而圆[47, 48, 49]。Gα 蛋白存在于多种真核生物中[50, 51, 52], 在哺乳动物中其功能缺失引起多种严重疾病[10, 51, 52]。在水稻中Gα 除参与GA和BR等激素应答外[29, 47], 还参与抗病反应[50, 53, 54]及光信号等多种途径[55]。d1突变体是研究Gα 蛋白功能及GA和BR信号途径的重要材料。
尽管目前已克隆多个植物株高基因, 但株高形成的调控网络仍不够清晰[4, 5]。因此, 有必要进一步鉴定不同的株高突变体、克隆更多的株高基因, 并深入研究植物矮化分子机制。本研究从水稻稻瘟病普感品种丽江新团黑谷(LTH)[56, 57]的EMS (ethylmethane sulfonate, 甲基磺酸乙酯)诱变群体中筛选到一个稳定的小粒矮化突变体LTH-m3, 通过图位克隆和功能基因互补验证确认该突变体是一个新的D1基因等位突变体, 其D1基因在第6外显子与内含子结合处的第2522位碱基由G突变为A, 导致第6外显子被剪切掉而无法翻译出有功能的Gα 亚基。水稻突变体LTH-m3及其功能基因的鉴定, 将为深入研究Gα 蛋白的功能及GA和BR信号途径提供有用材料。
突变体LTH-m3是水稻稻瘟病普感品种LTH经EMS诱变后分离出来的, 已经过多代自交, 性状稳定。将LTH-m3分别与野生型LTH及籼稻品种93-11杂交获得F1, 再经自交获得F2群体。LTH-m3与野生型杂交的F2群体用于矮化性状遗传分析, LTH-m3与93-11杂交的F2群体用于矮化基因定位。在中国农业科学院作物科学研究所北京昌平试验基地正季种植上述材料, 种植方式与管理同常规大田。
在成熟期, 针对突变体LTH-m3和野生型, 各选10株考查株高、分蘖数、粒长、粒宽等性状, 并以主茎为对象调查各节间长度、穗长、穗粒数等性状; 针对F2群体, 分别考查各单株及双亲的株高, 统计正常表型和矮化表型植株的数量。
将饱满的野生型和突变体种子经1%次氯酸钠溶液消毒和催芽处理后, 分别选取适量的萌发整齐一致的种子播于带孔的泡沫板上, 将泡沫板置盛有水的烧杯中, 放入30℃恒温箱暗处理。20 d后, 观察、记载中胚轴的伸长情况。
将饱满的野生型和突变体种子经1%次氯酸钠溶液消毒和催芽处理后, 分别选取适量的萌发整齐一致的种子播于96孔PCR板(下端剪掉), 每孔1粒种子, 将PCR板置离心管盒中, 加入500 mL蒸馏水培养。待芽鞘生长至1 cm左右时, 换用1× 10-9、1× 10-8、1× 10-6、1× 10-5 µ mol L-1的赤霉素GA3溶液(500 mL)分别处理, 以等量蒸馏水处理为对照。每个处理20苗以上, 3次重复。在光照培养箱30℃及14 h光照/10 h黑暗条件下培养7 d后, 测量苗高和根长, 并以蒸馏水处理为对照, 计算不同浓度GA3处理后的苗高增长率。苗高增长率(%) = (GA3处理的苗高 - 对照的苗高)/对照的苗高 × 100。
将饱满的野生型和突变体种子经1%次氯酸钠溶液消毒和催芽处理后, 分别选取适量的萌发整齐一致的种子播于96孔PCR板(下端剪掉), 每孔1粒种子, PCR板置于离心管盒中, 加入500 mL蒸馏水, 在光照培养箱30℃及14 h光照/10 h黑暗条件下培养。待第3片叶开始显现后, 用移液枪将浓度为1 μ mol L-1油菜素内酯eBL (Sigma公司)的乙醇溶液(1 μ L)点滴到野生型和突变体的第3叶的叶枕和叶尖, 以等量乙醇点滴作对照。每个处理20苗以上, 3次重复。每天点滴2次, 每次间隔12 h。7 d后, 用Image J软件测量第3叶的叶夹角。
利用均匀分布于12条染色体的170对SSR (Single sequence repeat)或InDel (Insertion and deletion)标记对LTH-m3和93-11进行PCR扩增, 筛选和利用亲本多态性标记用于目标基因的连锁分析。根据Gramene网站(http://www.gramene.org/)公布的93-11和日本晴序列的差异, 利用DNASTAR软件设计、开发新的InDel标记用于目标基因的进一步定位。各标记及其相关信息列于表1, 所有引物均由北京博迈德生物科技公司合成。
将新鲜叶片用液氮速冻后粉碎至粉末状, 采用CTAB法提取基因组DNA[30]。将DNA溶解于适量ddH2O中, 作为PCR扩增反应的模板。10 μ L反应体系含10× PCR缓冲液1 μ L, dNTPs (10 mmol L-1) 0.2 μ L, 正、反引物(2 μ mol L-1)各0.5 μ L, DNA 2 μ L (20~50 ng), Taq DNA聚合酶(2 U μ L-1) 0.1 μ L, ddH2O 6.1 μ L。PCR程序为95℃预变性5 min; 94℃变性30 s, 55~60℃退火30 s, 72℃延伸30 s, 35个循环; 72℃再延伸5 min。PCR产物经8%聚丙烯酰胺凝胶电泳检测及0.1% AgNO3溶液银染显色。
选取均匀分布于各染色体上的亲本多态性分子标记, 对F2群体中表现极端突变表型的单株进行基因型鉴定, 筛选与突变表型连锁的标记, 确定目标基因的候选区段。利用扩大的F2群体中极端突变表型个体, 通过增加候选区段内分子标记密度, 对目标基因精细定位。
根据从Gramene数据库(http://www.gramene. org/)下载的D1全长基因组DNA序列, 设计D1基因扩增引物(表1), 采用宝生物工程(大连)有限公司生产的高保真酶PrimeSTAR HS DNA Polymerase with GC buffer分别扩增野生型和突变体的基因组DNA。将PCR产物送北京博迈德科技发展有限公司测序。利用DNA STAR软件的SeqMan程序比对突变体和野生型的测序结果, 确定突变位点。
| 表1 本研究供试引物的信息 Table 1 Primers used in this study |
所用植物转化载体为pCAMBIA1305.1。根据预测的候选基因序列, 以LTH为模板, 设计引物1305D1用以扩增D1基因组全长7640 bp的片段, 包括起始密码子ATG上游2000 bp, 编码区3640 bp和终止密码子TAG下游2000 bp。利用Clonetech的In-FusionTM Advantage PCR Cloning Kits把该片段同源重组至植物双元载体pCAMBIA1305.1的Kpn I位点, 构建转化载体。载体质粒经测序验证正确后, 利用热激法将其转入农秆菌菌株EHA105中, 挑选阳性克隆转化水稻成熟胚愈伤组织。获得T0代转基因植株后, 将转基因植株叶片分别浸入GUS染液和潮霉素(50 μ g mL-1)中检测, 经GUS染色24 h后变蓝且叶片在潮霉素中光照72 h后不发黄被视为阳性转基因植株。将阳性植株种植于转基因隔离圃观察和繁种。
取生长1周左右的水稻幼苗经液氮研磨后, 用天根生化科技(北京)有限公司的RNAprep pure Plant Kit提取总RNA。用宝生物工程(大连)有限公司的PrimeScript 1st Strand cDNA Synthesis Kit合成第1链cDNA。用宝生物工程(大连)有限公司的SYBR Premix Ex Taq II试剂盒, 及Applied Bio systems公司的96孔PCR板在ABI PRISM 7500实时PCR仪上进行实时荧光定量PCR。实时荧光定量PCR以水稻的Ubiquitin (LOC_Os03g10170)基因作为内参, 反应程序为95℃下预变性30 s, 95℃下变性5 s, 60℃下34 s, 40个循环。设每个实验3个生物学重复, 采用2-Δ Δ CT算法[46]分析PCR结果。用于表达分析的基因及其特异性引物信息列于表1, 其中D10-Ex (D10)和D53-Ex (D53)分别为Zhou等[46]报道的引物D10和D53。所有引物均由北京博迈德生物科技公司合成。
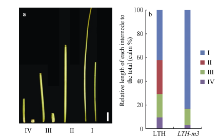
与野生型LTH相比, 突变体LTH-m3自出苗即表现矮化(图1-a, b), 叶片短宽且颜色深绿(图1-c), 生育期延迟5~6 d (表2), 成株期株高约为野生型的35.3%, 叶片上举, 株型紧凑(图1-e), 分蘖数和有效穗数显著增加(表2), 粒长和千粒重显著减少(图1-f, g, 表2)。据此看出, LTH-m3符合d1突变体的一些基本特征[47, 48, 49]。另外, 突变体的各个节间均较野生型显著缩短。其中, 第2节间基本缺失, 第1、第3和第4节间长度分别缩短至野生型的50.5%、17.8%和7.8% (图2-a)。按照Takeda的分类方法, LTH-m3属于“ dm” 类型的矮化突变体(图2-b)。
 | 图1 野生型与突变体LTH-m3的表型 a, b: 幼苗; c: 第4叶片; d: 暗培养下形态; e: 成熟期植株; f: 穗; g: 粒宽; h: 粒长. 图a~f的左侧为野生型LTH, 右侧为突变体LTH-m3。图g和h的上侧为LTH, 下侧为LTH-m3。白色箭头指向中胚轴。Fig. 1 Phenotypes of the wild type and the LTH-m3 mutant a, b: seedlings; c: the 4th leaf; d: plant under dark treatment; e: adult plant; f: panicle; g: grain width; h: grain length. From figure a to f, the left is wild type LTH and the right is mutant LTH-m3; in figures g and h, the upper is wild type LTH and the lower is LTH-m3. The arrows indicate the mesocotyl. |
| 表2 野生型和LTH-m3的主要农艺性状比较 Table 2 Comparison of major agronomic traits between wild type and LTH-m3 |
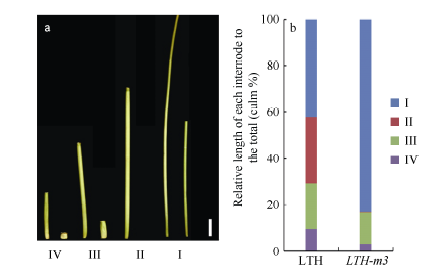 | 图2 野生型LTH和突变体LTH-m3各节间的长度及其伸长模式 a: LTH和LTH-m3各节间长度, 左侧为LTH, 右侧为LTH-m3, 标尺为5 cm。b: LTH和LTH-m3节间长度模式。I、II、III、IV分别表示穗下第1、第2、第3和第4节间。Fig. 2 Length and elongation model of the internodes in LTH and LTH-m3 a: internode length of LTH and LTH-m3. left: LTH; right: LTH-m3, the bar stands for 5 cm. b: internode elongation patterns of LTH and LTH-m3. I, II, III, and IV indicate internodes from the uppermost to the forth. |
在黑暗培养条件下, 正常水稻幼苗中胚轴会伸长, 而BR突变体的中胚轴不伸长, 这是BR突变体的特有反应[29, 40]。对LTH-m3及其野生型的幼苗黑暗培养处理, 发现野生型的中胚轴明显伸长而突变体的未伸长(图1-d), 这表明LTH-m3属于BR相关突变体。
用不同浓度赤霉素GA3处理7 d后, 发现突变体和野生型的苗高均有所增加, 但突变体未恢复到野生型的苗高(图3-a), 与对照相比, 处理后突变体的苗高增长率比野生型低(图3-b)。这些结果暗示着突变体LTH-m3是一个GA3钝感型矮化突变体。
 | 图3 GA3处理下突变体LTH-m3与野生型LTH苗高(a)及苗高增长率(b)的比较Fig. 3 Comparison of seedling height (a) and its relative increase (b) under treatments of GA3 between LTH-m3 mutant and wild type LTH |
以1 µ mol L-1油菜素内酯eBL的乙醇溶液持续处理7 d后发现, 与对照相比, 野生型的叶倾角极度增大(平均增加97.3º ± 4.0º ), 而突变体的叶倾角中度增大(平均增加31.8º ± 4.6º )(图4), 这意味着突变体LTH-m3对外源BR的敏感性较野生型明显降低。
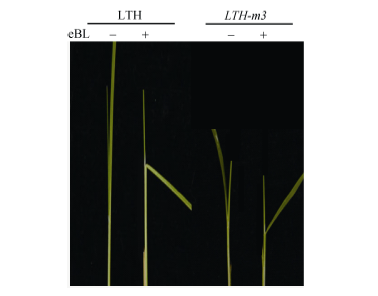 | 图4 eBL处理下LTH-m3与野生型LTH叶倾角的比较Fig. 4 Comparison of lamina joint inclination under treatment of eBL between LTH-m3 mutant and wild type LTH |
矮化突变体LTH-m3与野生型LTH杂交后的F1的株高均与LTH一致, 说明引起突变的基因为隐性基因, 在所调查的包含331个单株的F2群体中, 有258个单株表现野生型的表型(高秆正常粒), 73株表现突变体表型(矮化小粒), 未发现表现高秆小粒或矮化正常粒的单株, 且野生型单株与突变体单株分离比例符合3 : 1 (χ 20.05= 1.38 < χ 20.05= 3.84), 这表明LTH-m3的矮化和小粒性状均受单个隐性基因控制, 且该二性状间存在“ 一因多效” 或紧密连锁关系。
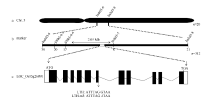
LTH-m3与籼稻品种93-11杂交后的F1株高与野生型LTH一致, 但F2群体分离复杂, 出现了高秆(类似LTH)、超高秆、半矮秆、矮秆(类似LTH-m3)、超矮等多种表型。为准确定位目标基因, 首先从该F2群体中筛选出20个表现突变体表型的单株, 利用覆盖12条染色体的亲本多态性标记进行基因型分析, 结果将目标基因定位到第5染色体上标记Indel5-6和Indel5-8之间(图5-a)。进一步, 利用该区间内多态性标记和312个表现突变表型的F2单株将目标基因锁定在标记SSR5-6-4和Indel5-7之间2.05 Mb的区域(图5-b)。在日本晴序列中, 该区域包含一个矮秆基因LOC_Os05g26890, 即编码一个GTP结合蛋白的α 亚基(Gα )的D1基因(http://www.gramene. org/), 其突变后引起的突变体表型与LTH-m3类似, 故优先将D1确定为目标基因的候选基因。
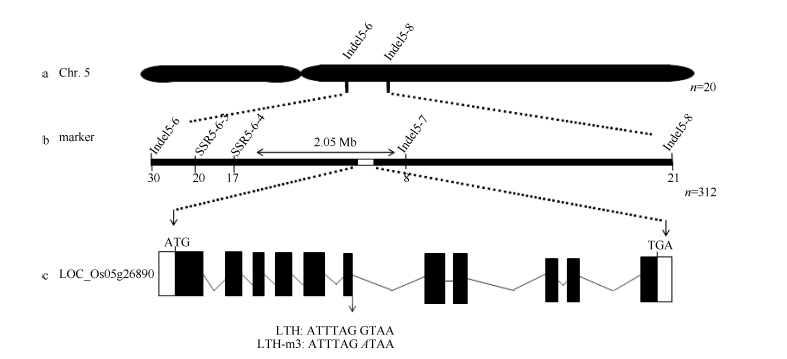 | 图5 突变体LTH-m3矮化基因的定位 a, b: 分别利用20个和312个突变表型的F2单株定位的基因位置。c: D1基因结构示意图, 白色框代表5′ 和3′ 非翻译区, 黑色框代表外显子区, 横线代表内含子区, 箭头指向突变体中D1的突变位点, 位于第6个内含子与外显子剪接位点处。Fig. 5 Mapping of the dwarf gene in mutant LTH-m3 a, b: gene mapping using 20 and 312 mutant-like F2 plants, respectively. c: the structure of D1, the white, black boxes represent 5′ and 3′ end un-translated regions and exons, and the horizontal lines represent introns. The arrow indicates the mutation position in the 6th intron of D1 in LTH-m3. |
在日本晴中, D1基因全长3640 bp, 包含11个外显子和10个内含子(图5-c), 其cDNA全长1032 bp, 编码的Gα 亚基(D1蛋白)包含343个氨基酸(http://www.gramene.org/)。利用引物D1-F/R分别扩增野生型LTH和突变体LTH-m3中D1基因的启动子区和编码区, 经引物D1-1和D1-2测序、拼接及序列比对发现, 在突变体中, D1基因在启动子区的序列与野生型的完全一致, 而其编码区ATG下游第2522个碱基由野生型的G突变为A, 该位点位于第6个外显子与内含子剪接位点处(图3-c)。对突变体和野生型中D1基因的cDNA克隆, 用引物D1-6测序及序列比对发现, 突变体中第2522碱基G→ A的改变直接引起了剪切方式的变化, 导致突变体中D1基因第6个外显子被剪切掉(图6-a), 所编码的Gα 亚基缺少了3个效应因子结合域(effector binding region)及受体结合域(receptor binding region)(图6-b)。因此, 推断D1基因的功能缺失是造成LTH-m3突变体表型的直接原因。
采用农秆菌介导方法, 将包含野生型中D1基因全长7640 bp的互补载体转化到突变体LTH-m3中, 所有T0和T1代阳性转基因植株能恢复到野生型的正常株高(图7-a), 其T1代种子能恢复到野生型的籽粒大小(图7-b), T1代阳性株的幼苗也能恢复到野生型的正常苗高(图7-c)。这些结果证明D1基因的突变是导致突变体矮化和小粒表型的直接原因。
 | 图7 D1转基因互补表型 a: 抽穗期野生型、突变体及转基因阳性株T0代植株表型; b: 成熟期野生型、突变体及转基因阳性株T1代种子表型; c: 苗期野生型、突变体及转基因阳性株T1代植株表型。Fig. 7 Phenotypes complemented by transforming D1into the mutant a: phenotypes of the wild type, mutant and T0 transformant plants at heading stage; b: phenotypes of the wild type, mutant and T1 transformant seeds at maturing stage; c: phenotypes of the wild type, mutant and T0 transformant seedlings. |
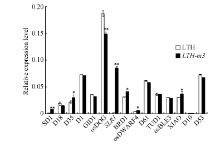
为了解突变体LTH-m3中D1基因突变对GA、BR和SL相关矮化基因表达模式的影响, 我们选择3个参与GA合成的基因(SD1、D18、D35)、4个参与GA信号传导的基因(D1、GID1、OsDOG、SLR1)、2个参与BR合成的基因(BRD1、OsDWARF4)、4个与BR信号传导有关的基因(D61、TUD1、OsBLE3、XIAO)及分别参与SL合成和信号传导的基因D10和D53等, 并对这些基因进行了实时荧光定量PCR分析。与野生型相比, 在LTH-m3中有7个GA和BR相关基因的表达发生了显著改变(图8), 其中SD1、D35、SLR1、BRD1、OsDWARF4、XIAO的表达量分别上调6.4、0.4、64.4、0.3、0.6和0.2倍, 而OsDOG的表达量下调0.2倍。这些结果表明, D1基因突变可导致LTH-m3植株细胞内的GA和BR信号传导受阻, GA和BR合成反馈调节功能也受到影响, 这可能进一步影响到受GA和BR调控的下游细胞分化和伸长调控因子的功能及植株生长发育过程, 最终导致突变体异常表型的发生。
 | 图8 野生型LTH和突变体LTH-m3中部分矮化相关基因的表达Fig. 8 Expression of some dwarf genes in the mutant LTH-m3 and wild type LTH |
云南地方粳稻品种丽江新团黑谷(LTH)不携带任何主效稻瘟病抗性基因, 对世界各稻区的稻瘟病菌表现普遍感病, 是开展稻瘟病菌致病性和水稻抗病性遗传等研究的理想材料[56, 57], 其缺点是植株过高且茎秆软, 极易倒伏。为了降低LTH株高, 提高其抗倒伏能力, 我们用EMS诱变处理LTH获得了部分矮化突变体。本文报道了一个小粒矮化突变体LTH-m3, 属“ dm” 类型的矮化突变体, 在苗期和成株期的高度分别约为野生型的1/2和1/3, 且其节间相对粗壮(图1-a, b, e, 图2, 表1), 完全克服了野生型苗期徒长和成株期植株过高且茎秆软和易倒伏等缺点。另外, 本实验室资料显示LTH-m3对供试的300多个国内外稻瘟病菌株无一例外表现高度感病, 表明其对稻瘟病菌的普感特性未因D1基因突变而受到影响。因此, LTH-m3可被视作LTH的改良系在今后的水稻稻瘟病研究中加以利用。进一步的研究证实, LTH-m3在黑暗条件下中胚轴不伸长(图1-d), 属于油菜素内酯(BR)相关突变体; 其对外源赤霉素GA3不敏感, 对外源BR的敏感性也较野生型明显降低(图3和图4); 其小粒和矮化性状均受隐性基因d1控制, D1基因的功能缺失是导致突变体表型的直接原因(图5, 图6, 图7)。这些结果为有效利用该矮化突变体或矮化基因开展株型改良及株高形成机理研究奠定了理论和物质基础。
目前已报道10多个d1突变体[7, 10, 58], 其中d1-4为弱等位突变体, 表现中粒半矮化表型, 其余的均为强等位突变体, 表现小粒矮化表型。d1-4表型是D1蛋白在Gα -GTP(A)结构域单个氨基酸突变所致, 而其他强等位突变体几乎都是Gα 蛋白氨基酸缺失突变或翻译提前终止所造成的。不同等位突变体由于D1基因突变位点不同可能出现一些在表型上不易察觉的差异, 例如d1-1 [Gα -GTP(G)后区段因2个碱基缺失而翻译提前终止]和d1-5 [Gα -GTP(A)后区段因单个碱基替换而翻译提前终止]在正常生长条件下表现相似的表型, 但在暗条件下表现前者节间不伸长而后者与野生型相同的节间伸长[47]。再如d1-4和d1-8 [Gα -EBR(3)结构域中单个氨基酸缺失], 前者在质膜上有Gα 累积而后者却无Gα 累积[58]。不同的d1等位基因的发现和研究不仅有助于了解Gα 构象及其与效应因子的相互作用, 而且可为系统深入研究株高形成的调控机理及激素信号传导过程提供新的信息。
LTH-m3是一个新的d1基因等位突变体, 其D1基因在第6个外显子与内含子剪接位点处[Gα -GTP (A)至Gα -GTP(C)区段的中间]发生一个碱基突变(G2522→ A2522), 导致第6个外显子被选择性剪切及终止密码子产生, 进而导致Gα 亚基翻译提前终止(图6); 但实时荧光定量PCR结果显示, 第6个外显子的选择性剪切并未影响突变体中D1基因的转录表达水平(图8)。LTH-m3这种特殊的突变方式是否会引起一些有别于其他d1突变体的非表型的差异, 尚待进一步考查和研究。鉴于选择性剪切对调控植物基因转录后表达及植物体形成和正常运转至关重要[59], 且水稻基因剪切位点处“ G→ A” 碱基突变鲜见报道[60], 突变体LTH-m3抑或对水稻基因选择性剪切研究具有参考价值。
 | 图6 突变体中D1基因的CDS序列和D1蛋白结构域突变位点 a: 野生型和突变体中D1基因CDS序列比对, 红色下画线部分为剪切掉的第6外显子。b: 突变体中D1蛋白结构域示意图, 第6外显子的缺失导致翻译提前终止, 箭头是翻译终止位置。Fig. 6 CDS sequence of D1 gene and the mutation site of D1 domain in the mutant a: alignment of CDS sequences of D1 in the mutant and wild type, in which the red underline indicates the cut 6th exon. b: a schematic diagram of the mutation in D1 protein, and the arrow indicates the mutation position of preterm stop codon in the mutant. |
水稻D1基因参与GA和BR双信号途径[27, 49]。在突变体LTH-m3中, GA合成相关基因SD1和D35, GA信号传导相关基因SLR1, BR合成相关基因BRD1、OsDWARF4及BR信号传导相关基因XIAO等的表达显著上调, 而另一GA信号传导相关基因OsDOG的表达显著下调, 特别是编码GA生物合成关键酶— GA20氧化酶的SD1和编码与拟南芥RGA/GAI同源的DELLA蛋白的SLR1基因的超高表达(图8), 表明D1基因的突变确实影响了植株细胞内GA和BR的信号传递及其合成的反馈调节功能。在GA信号途径中, D1是GA一个受体蛋白和信号传导的正调控因子[28, 61]; SLR1编码的DELLA蛋白是GA信号传导的负调控因子, 参与GA动态平衡的反馈调节[61]。SLR1位于D1的下游, 对D1具有上位性[61]。在LTH-m3突变体中, SLR1和SD1基因之所以超高上调表达, 推测可能的原因是D1基因功能的缺失导致GA不能被有效运输到细胞内, 造成胞内GA浓度低, 进而刺激其负调控因子SLR1和其上游的合成基因SD1的转录和翻译。在BR信号途径中, D1被认为影响BR信号级联(cascade)而不是BRI1介导的识别和转导的信号分子[47], 或与TUD1直接互作参与BR信号传递[34]; BRD1编码BR生物合成的一个关键酶, 即BR-6氧化酶[29]; OsDWARF4编码一个调节BR合成速率的P450蛋白CYP90B1, 其表达受反馈调节[41]; XIAO编码一个LRR激酶, 调控BR信号传递和动态平衡[36]。在突变体LTH-m3中, 这3个基因的表达上调可能与BR反馈调节功能受到影响有关。植物株高的形成是一个非常复杂的过程, 到目前为止, 有关植物矮化的机理尚未阐述透彻, Gα 蛋白参与调控的GA和BR信号传导的具体过程也很不清晰[7]。因此, 有必要发掘和鉴定更多的矮化突变基因来研究D1基因上下游之间的调控关系, 为揭示Gα 的生物学功能及GA和BR信号传导过程提供依据。
水稻突变体LTH-m3是一个新的d1基因等位突变体。D1基因第6个外显子与内含子接合处发生单碱基突变(G2522→ A2522), 引起第6 外显子被选择性剪切及Gα 蛋白翻译提前终止, 这是造成LTH-m3小粒矮化表型的直接原因。该突变体中D1基因突变引起SD1和SLR1等基因表达显著改变, 进而影响植株细胞内GA和BR合成的反馈调节功能和信号传递。突变体LTH-m3弥补了LTH植株过高、茎秆软和极易倒伏的缺点, 在今后水稻稻瘟病研究中可用作LTH的改良系而加以利用。LTH-m3功能突变基因的确认, 为深入探讨水稻Gα 蛋白的功能及信号途径提供了新材料。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|