
{kind=link}
水稻粒形基因 GW8的功能标记开发和单体型鉴定
[裔传灯*  , 王德荣, 蒋伟, 李玮, 成晓俊, 王颖, 周勇, 梁国华, 顾铭洪]
, 王德荣, 蒋伟, 李玮, 成晓俊, 王颖, 周勇, 梁国华, 顾铭洪]
, 王德荣, 蒋伟, 李玮, 成晓俊, 王颖, 周勇, 梁国华, 顾铭洪]
|
|


水稻粒形性状包含粒长、粒宽、粒厚和长宽比, 是一类影响水稻产量和品质的重要性状。基于基因 GW8序列分析, 在该基因启动子区10 bp的Indel和第3外显子A/C和T/G的2个变异位点分别开发了功能标记, 并将其用于294份水稻微核心种质和2007—2013年江苏省审定的65份粳稻品种的基因型鉴定。研究结果表明这3个变异位点的等位变异对主要的粒形性状有着显著或极显著的影响; 其在水稻微核心种质存在的8种单体型对水稻粒形性状有着极显著的影响, 其中分别有126、85和58个代表品种的Hap3、Hap6和Hap1是最主要的单体型, 单体型Hap7对应水稻籽粒的粒长最长、长宽比最大、千粒重最大; 而江苏省审定的粳稻品种中仅发现分别有63个和2个代表品种的单体型是Hap6和Hap2。这些研究结果为水稻产量和品质育种中充分利用基因 GW8的优异等位基因或单体型奠定了基础。
, WANG De-Rong, JIANG Wei, Li Wei, CHENG Xiao-Jun, WANG Ying, ZHOU Yong, LIANG Guo-Hua, GU Ming-Hong
Rice grain shape including grain length, grain width, grain thickness, and grain length-to-width ratio is a major character affecting rice yield and quality. On the basis of the alignment of gene GW8 genomic DNA sequence, three functional markers were developed on the 10-bp indel in promoter region and two missense polymorphism sites (A/C and T/G) in the exon3, respectively. Subsequently the markers were used to identify the genotypes of gene GW8 in the 294 accessions of rice mini-core collection and 65 japonica varieties certificated in Jiangsu province from 2007 to 2013. We found that the allelic variations of the three target loci had significant or very significant effects on most traits of grain shape. On the bais of the three variation sites of gene GW8, eight haplotypes (combinations) were found to have very significant effect on the four grain shape traits in the rice mini-core collection. The main haplotypes were Hap3, Hap6, and Hap1, contained in 126, 85, and 58 representative varieties, respectively; Hap7 has the most strongest effect on grain length, ratio of length to width and 1000-grain weight. While the only two haplotypes, Hap6 and Hap2, contained in 63 and 2 japonica varieties certificated in Jiangsu provice, respectively. The results paved a way to apply the useful allelic variations or haplotypes of gene GW8 in rice yield and quality breeding programs.
水稻产量是由单位面积株数、每株穗数、每穗实粒数和千粒重4个因素共同决定的, 其中千粒重主要受水稻籽粒的粒形影响。水稻的粒形性状包括粒长、粒宽、粒厚和长宽比[1]。目前多个水稻粒形相关基因, 如GW2[2]、GS3[3]、qGL3/GL3.1[4, 5]、qSW5/ GW5[6, 7]、GS5[8]、GS6[9]、GW7[10]、GW8[11]、SLG7[12]、TGW6[13]等克隆研究证实, 通过调节粒形性状可以增加千粒重, 从而提高水稻产量水平。同时GW8[11]和GW7[10]对稻米品质, 尤其是稻米的外观品质也有着重要的影响[14]。因此粒形性状调控机制的研究对水稻的产量育种和品质育种有着重要的参考价值。
GW8是水稻重要粒形基因之一。Wang等[11]研究发现基因GW8编码细胞增殖的正调控因子, 来自水稻品种华粳籼74的基因GW8启动子区10 bp序列(GAGCTGAGCT)的存在, 对于该基因在幼穗中的正常表达有着重要的作用, 从而增加了籽粒的粒宽, 进而提高了水稻产量; 而来自水稻品种Basmati 385的基因gw8启动子区10 bp序列的缺失, 导致该基因表达降低, 籽粒的宽度变小, 从而提高了稻米的外观品质。目前该基因其他的变异位点对水稻粒形性状有何效应还不清楚。
为了加快基因GW8有利等位变异在水稻育种中的应用, 本研究对已知的功能变异和未知功能的错义突变开发了相应的功能标记, 结合水稻微核心种质和近年来江苏审定的粳稻的基因型检测, 分析了这些变异位点和组合(单体型)对水稻粒形性状的影响, 以期为我国和江苏省的水稻产量和品质育种提供理论依据和快捷的选择手段。
选用294份水稻微核心种质和65份江苏省在2007— 2013年期间审定的粳稻品种[15]。水稻微核心种质具有丰富的遗传多样性, 其包括从1530份国外栽培稻品种中筛选出的96份微核心种质, 从4310份中国栽培稻品种中筛选出的198份微核心种质[16]。这些种质材料覆盖全球五大洲74个国家或研究机构[17], 来自国内外不同稻作区, 它们的感光性存在较大的差异。为确保能够正常抽穗, 2013年11月于海南陵水播种所有供试水稻品种并育苗, 2014年1月移栽大田, 田间管理同常规水稻品种。待水稻籽粒完全成熟后收种, 晒干后测量水稻籽粒相关性状。
参照《水稻种质资源描述规范和数据标准》的方法[18]收获并风干水稻种子, 挑选饱满成熟种子使用游标卡尺(精确到0.01 mm)测量粒长、粒宽和粒厚, 5次重复, 计算平均值。用电子天平测定1000粒成熟烘干种子的重量, 3次重复, 计算平均值。
从Rice Genome Annotation Project网站(http:// rice.plantbiology.msu.edu/)下载到GW8基因的基因组DNA序列(LOC_Os08g41940)。以LOC_Os08g 41940序列作为种子序列, 在NCBI网站核酸序列数据库中找到高度同源的3个基因组DNA序列(JX867117、JX867118和JX867119)和1个cDNA序列(AK109469)。借助BioEdit软件对上述序列比对分析。
利用Primer Premier 5.0软件对基因GW8启动子区一个10 bp的Indel序列变异和第3外显子的2个错义突变的点突变分别设计测序引物、dCAPs和CAPs标记。由生工生物工程(上海)股份有限公司合成引物和测序。
收集供试材料分蘖盛期新鲜幼嫩的叶片, 采用SDS法提取水稻基因组DNA。
PCR体系含50 ng μ L-1基因组DNA 2.0 μ L、2 µ mol L-1引物F和R各2.5 μ L、10× 缓冲液2.0 μ L、25 mmol L-1 MgCl2 2.0 μ L、2.5 mmol L-1 dNTP 1.6 μ L、5 U μ L-1Taq DNA聚合酶(TaKaRa code: R001C) 0.2 μ L或1.25 U μ L-1高保真DNA聚合酶(PrimeSTAR GXL DNA Polymerase, TaKaRa Code: DR050S) 1.0 μ L, 以灭菌双蒸水补足至20 μ L。在Eppendorf Master cycler proS PCR仪上进行扩增, 反应条件为: (1) 94℃, 预变性5 min; (2) 94℃, 30 s; 55~60℃, 30 s; 72℃, 1 min; 共35个循环; (3) 72℃再延伸10 min。反应产物经3.0%的琼脂糖凝胶电泳分离, 筛选出最清晰、单一且没有杂带的引物。
利用筛选的PCR引物扩增基因GW8的目标片段, 进一步用于酶切反应。酶切反应体系为10 μ L, 含PCR反应产物5 μ L、10× buffer 1 μ L、酶(10 U μ L-1) 0.25 μ L、ddH2O 3.75 μ L。混匀后置37℃恒温水浴锅酶切3~4 h, 酶切产物经3.0%琼脂糖凝胶电泳, EB染色, 紫外凝胶成相系统成像。
用Microsoft Excel和SPSS软件分析和处理数据。
序列比对分析表明, 基因GW8的基因组DNA序列有52处变异, 其中46个变异发生在非编码区。除了Wang等[11]已报道位于启动子区(转录起始点上游23 bp处)的10 bp Indel变异与该基因功能密切相关外, 该基因的3个外显子中, 共有6个序列变异位点。以基因GW8的编码序列LOC_Os08g41940作为参照, 第1外显子第36 bp、236 bp和329 bp处分别有C/T同义突变、C/T错义突变和CGG的Indel变异, 其中后2个变异与启动子变异类型高度相关。第3外显子的第821 bp、1092 bp、1119 bp处分别有C/A错义突变、G/T错义突变和G/A同义突变。因此对基因GW8的启动子区的10 bp Indel变异和第3外显子的2个错义突变位点分别开发了测序引物(GW8-1)、dCAPs (GW8-2)和CAPs (GW8-3)标记(表1)。
| 表1 基于基因GW8启动子区和第3外显子DNA序列变异设计的PCR引物 Table 1 PCR primers based on the GW8 DNA sequence variations in promotor and exon3 |
利用分子标记GW8-1对包含基因GW8启动子区变异区域的151 bp的DNA序列进行高保真扩增, 将获得的PCR产物(图1-A)测序, 完成待测水稻品种基因GW8启动子变异类型的判定。
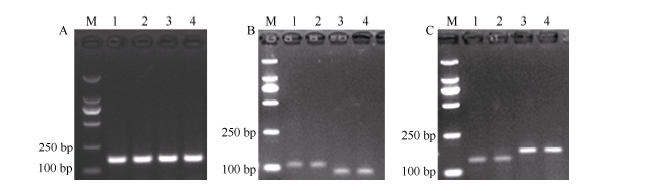 | 图1 基因GW8不同变异位点特异引物的PCR鉴定 A: 是引物GW8-1的扩增产物电泳图; B和C: 分别是引物GW8-2和GW8-3的扩增产物酶切电泳图。M: DL2000 DNA marker (TaKaRa), 泳道1~4: 分别为水稻品种Basmati、日本晴、9311和华粳籼74。Fig. 1 PCR identification of specific primers for corresponding mutation sites in geneGW8 A: PCR amplification products of primer GW8-1; B and C: enzyme-digested PCR amplification products of primer GW8-2(B) and GW8-3(C). M: DL2000 DNA marker (TaKaRa); lanes 1-4: Basmati, Nipponbare, 9311, and Huajingxian 74, respectively. |
对于基因GW8第3外显子的第821 bp处的C/A单碱基变异, 利用标记GW8-2在水稻品种中扩增出长度为108 bp的PCR产物, 经过限制性内切酶Hind II酶切后, 能够被切成89 bp条带的水稻材料基因型为A (如图1-B的泳道3和4); PCR产物仍为108 bp的水稻材料基因型为C (如图1-B的泳道1和2)。
对于基因GW8第3外显子的第1092 bp处的G/T单碱基变异, 利用标记GW8-3在水稻品种中扩增出长度为164 bp的PCR产物, 经过限制性内切酶Nco I酶切后, 能够被切成134 bp条带的水稻材料基因型为G (如图1-C的泳道1和2); PCR产物仍为164 bp的水稻材料基因型为T (如图1-C的泳道3和4)。
从表2可以看出, 基因GW8的启动子10 bp的Indel变异位点和第3外显子的G/T变异位点对粒长、粒宽、粒厚和籽粒长宽比都有极显著的影响; 第
3外显子的C/A变异位点对粒长和粒宽有极显著的影响, 对粒厚有显著影响; 但是这3个变异位点对千粒重都没有显著影响, 这表明基因GW8的3个等位变异对水稻籽粒的粒形性状有着重要的影响。
| 表2 基因GW8不同变异位点的粒形性状及其t测验 Table 2 Grain-related traits and their t-tests of different allels in gene GW8 |
为了进一步明确基因GW8这3个变异位点的不同组合(单体型)对水稻籽粒相关性状的效应, 依据这3个变异位点在水稻微核心种质中的不同组合将基因GW8分成了8个单体型(表4)。因为单体型8 (Hap8)只有一个样本, 无法参与方差分析, 所以利用基因GW8的其余7个单体型对水稻粒形性状进行了方差分析(表3)和多重比较(表4)。
| 表3 基因GW8不同单体型粒形性状的方差分析 Table 3 Analysis of variances (ANOVA) of different haplotypes in gene GW8 |
从表3可看出, 除千粒重外, 基因GW8的不同单体型对水稻籽粒的粒长、粒宽、粒厚和籽粒长宽比这4个粒形性状都有着极显著的影响。多重比较进一步表明基因GW8的不同单体型对这4个粒形性状也有着不同程度的影响(表4), 其中具有Hap7的水稻籽粒的粒长最长、长宽比最大、千粒重最大, 具有Hap5的水稻籽粒的粒宽最大、粒厚最大、长宽比最小。
| 表4 基因GW8不同单体型对水稻粒形性状的差异显著性分析 Table 4 Analysis of the difference of grain-related traits based on different haplotypes in gene GW8 |
从基因GW8的各单体型分布频率来看, 以Hap3对应的水稻品种最多, 占水稻微核心种质的42.86%; 其次为Hap6和Hap1, 分别占水稻微核心种质的28.91%和19.73%。这表明基因GW8的单体型在水稻种质中分布不是均匀的, 这可能与水稻育种家对水稻籽粒的粒形性状选择有关。
为了更好地指导育种家在水稻育种中对籽粒相关性状的选择, 对2007— 2013年江苏省审定的65份粳稻品种进行了基因GW8的3个目标变异位点的基因型测定。从表5可以看出, 近年来江苏省审定的粳稻品种中, 2个水稻品种利用基因GW8的Hap2单体型; 而其余63个水稻品种都为单体型Hap6, 具有该单体型的水稻粒形表现为粒长较短、粒宽较宽、粒厚较厚、长宽比较小的特点。这表明江苏省近年来育成水稻品种的粒形基因GW8遗传基础比较狭窄。
| 表5 近年来江苏省审定水稻品种基因GW8不同单体型的粒形性状 Table 5 Grain-related traits of gene GW8 different haplotypes in recently certified new rice varieties |
籽粒是水稻一生中最大的库, 是水稻光合产物重要的贮存器官。调节水稻籽粒的大小和形状(即粒形), 增加光合产物的积累, 与提高产量(尤其是产量的构成因子千粒重)有着直接的关系。Wang等[11]研究发现, 在水稻品种Basmati基因gw8的背景下, 无论基因GS3存在与否, 来源于水稻品种华粳籼74的基因GW8都可以通过增加粒宽提高千粒重。但是本研究发现这3个变异位点对水稻的粒长、粒宽和粒厚有着极显著或显著的影响, 而对千粒重的影响没有达到显著水平, 这可能是由于水稻的千粒重涉及多个基因位点, 或者基因GW8对千粒重的影响较小。
随着水稻分子生物学的进展, 虽然许多与水稻产量相关的重要功能粒形基因已被克隆[3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13], 但是这些基因还很少在水稻育种工作中得以利用, 主要受制于这些基因缺乏开发成本低廉且简便实用的基因功能标记, 且不明确这些基因有多少等位变异类型及各等位变异类型的效应如何等等。利用近等基因系和转基因的方法, Wang等[11]证实了基因GW8的启动子10 bp Indel变异对水稻粒形有重要的影响, 其研究中涉及的基因型主要为水稻品种华粳籼74的单体型Hap3和Basmati的单体型Hap6。此外, 我们还借助基因GW8第3外显子的2个错义突变的功能标记, 检测到水稻微核心种质的其他6个不同的单体型, 这些不同的单体型对粒形有着不同程度的影响。一般认为细长粒垩白粒率较低, 其稻米外观品质较好[19, 20]。本研究在基因GW8的所有单体型中, 单体型Hap7对应水稻籽粒的粒长最长、长宽比最大, 有利于改善稻米的外观品质; 同时其千粒重最高, 也有着提高水稻产量的潜力。这种单体型具有较高的水稻育种应用价值, 但是在当前育成的水稻品种中还没有得到利用。单体型Hap5对应的水稻籽粒具有宽、厚和长宽比小的特点, 有助于提高整精米率, 进而改善稻米的碾磨品质。江苏省近年来审定的粳稻品种主要利用基因GW8的单体型Hap6, 其籽粒短、宽、厚、长宽比小, 千粒重较小, 不利于提高水稻产量和改善稻米外观品质, 因此还需要进一步加强对粒形有利基因资源的开发和利用。
基于水稻基因GW8的启动子区的10 bp的Indel变异和第3外显子A/C和T/G两处错义突变, 分别开发了特异性的功能标记, 结合水稻微核心种质, 分析了这3个变异位点及其组合而成的8种单体型对水稻粒形的影响, 同时发现江苏省近年来审定的粳稻品种主要利用了其中的一种单体型。
致谢: 感谢华南农业大学张桂权教授提供的水稻品种Basmati 385种子, 中国农业大学李自超教授和张洪亮博士提供的水稻微核心种质, 江苏省农业科学研究院王军博士提供的2007— 2013年江苏省审定的粳稻品种。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|