
{kind=link}
{kind=link}
大豆籽粒大小与形状性状的QTL定位
[陈强* , 闫龙* , 邓莹莹, 肖二宁, 刘兵强, 杨春燕*  , 张孟臣
, 张孟臣* ]
, 张孟臣]
|
|


第一作者联系方式: 陈强, E-mail: chenqiangwsm@163.com; 闫龙, E-mail: dragonyan1979@163.com
大豆籽粒大小和粒形性状不仅与产量和外观品质紧密相关, 还对机械化播种有着一定的影响。本研究采用大粒栽培品种冀豆12与小粒半野生地方品种黑豆(ZDD03651)杂交衍生的包含188个重组自交系的F6:8和F6:9群体为材料, 对粒长、粒宽、粒厚、长宽比、长厚比和宽厚比的遗传结构进行分析, 并分别以WinQTLCart 2.5、QTLNetwork 2.1和IciMapping 4.1 3种模型对以上性状的加性效应QTL, QE互作效应及上位性互作效应进行检测。6个性状的广义遗传率介于64.01%~79.57%, 遗传力较高, 且除粒厚外的其他性状受环境影响显著。共定位到加性效应QTL 38个, 单个QTL的贡献率介于2.21%~10.71%之间, 分布在12条染色体的17个标记区间内, 且12个染色体区段至少与2种性状相关。两种及以上模型同时检测到的QTL有24个, 3种模型均能检测到的QTL共8个, 分别为 qSL-17-1、qSL-18-1、qSW-6-1、qST-2-1、qST-6-1、qSLT-2-2、qSWT-2-1和 qSWT-20-1。检测到7对上位性互作QTL, 分别涉及粒长、粒宽、长宽比、长厚比和宽厚比, 互作效应贡献率介于0.78%~6.20%之间。QE互作效应贡献率均较低, 介于0.0005%~0.3900%之间。以多种模型同时检测结果准确性较高, 可为分子标记辅助育种工作提供可靠理论基础。
, ZHANG Meng-Chen
Seed size and shape not only relate to seed yield and quality, but also affect mechanical seeding in soybean ( Glycine maxL.). In this study, F6:8 and F6:9populations derived from Jidou 12 × Heidou were used to analyze the genetic character and detect quantitative trait loci (QTLs) for seed length, seed width, seed thickness, seed length-to-width ratio, seed length-to-thickness ratio, and seed width-to-thickness ratio. Softwares WinQTLCart 2.5, QTLNetwork 2.1 and IciMapping 4.1 were used to identify the additive, epistatic and environmentally interacted QTLs for seed size and shape related traits. As results, the heritability of the six traits varied from 64.01% to 79.57%. A total of 38 additive QTLs were identified to be located on 12 chromosomes, with the heritability varying from 2.21% to 10.71%. Eight of them ( qSL-17-1, qSL-18-1, qSW-6-1, qST-2-1, qST-6-1, qSLT-2-2, qSWT-2-1, and qSWT-20-1) were identified using three methods, simultaneously. In the meantime, seven pairs of additive × additive epistasis were detected and the heritability of epistasis pairs ranged from 0.78% to 6.20%. Additionally, the effects of QTL by environment interaction ranged from 0.0005% to 0.3900%. The QTLs identified using different mapping softwares in this study could provide a reliable theoretical basis for marker-assisted selection breeding.
在大豆中, 粒长、粒宽、粒厚等籽粒大小性状是衡量百粒重的重要指标, 与产量密切相关[1]。而大豆粒形性状直接关系大豆的品质和商品性, 既是机械化播种的重要限制因素, 又在一定程度上决定大豆在国际贸易中的商业价值[2]。籽粒大小和粒形性状为多基因控制的数量性状[3], 籽粒大小性状遗传力一般为59%~79%, 粒形性状的遗传力一般在19%~56%间[4]。通过QTL定位分析, 检测籽粒大小和粒形相关QTL位点, 对分子标记辅助选择的高产、高品质大豆育种具有重要的指导意义。
在水稻中籽粒大小和粒形的研究较为深入[5, 6, 7, 8, 9], 定位了较多QTL, 并克隆了控制粒长和粒重的基因GS3[5], 控制粒宽和粒重的基因GW2[6], 控制粒宽、长宽比和粒重的基因qSW5[7]等多个基因。大豆籽粒大小和粒形相关性状QTL定位的研究相对较少[10, 11, 12, 13]。大豆数据网站SoyBase (http://www.soybase.org/)中公布了粒长QTL 18个、粒宽QTL 16个、粒厚QTL14个。近期, 牛远等[14, 15]在初定位基础上, 采用Lasso法和复合区间作图法, 同时将O连锁群中Satt331- Satt592区段内多效性QTL分解后精细定位, 并对目标区段内候选基因进行预测。Xie等[16]对位于C2连锁群Satt640-Satt422区间内的粒形QTL精细定位分析, 分别检测到粒长、粒宽、粒厚及百粒重相关联的SNP标记, 并预测了结果区段内可能的效应基因。Hu等[17]在分离群体中检测到12个籽粒大小和粒形相关QTL, 在关联分析中检测到41个SNP标记与6个性状相关联。而随着研究的深入, QTL间的上位性互作及QE互作逐渐受到研究人员的重视[18, 19, 20]。Niu等[21]以257份大豆栽培品种为材料, 采用关联分析方法检测到59个主效QTL和31个QTL与环境互作效应。梁慧珍等[22]利用QTLNetwork 2.0软件检测到3对粒形性状上位性互作QTL。闫宁等[23]采用Bayes分层广义线性模型对粒形性状的主效QTL、QTL与环境互作效应、QQ上位性互作及QTL与细胞质互作效应分别进行检测。苏成付等[24]和李杰勤等[25]研究结果显示不同的定位程序适用的遗传模型范围不同, 建议使用多种作图法共同作图, 并优先标定共同发现的QTL。
目前, 大豆粒形相关性状上位性互作及QE互作研究相对较少, 且前人研究多采用单一模型检测QTL。本研究利用重组自交系群体为材料, 分别采用WinQTLCart 2.5[26]、QTLNetwork 2.1[27]和IciMapping 4.1[28, 29]软件对籽粒大小和粒形各性状的加性QTL, QTL间上位性互作及QE互作进行检测, 以得到可靠的QTL信息, 为进一步精细定位及辅助育种提供依据。
以冀豆12× 黑豆杂交构建的包含188个家系的F6:8和F6:9重组自交系为材料。冀豆12为河北省农林科学院粮油作物研究所育成的百粒重为22 g的大粒品种, 黑豆(ZDD03651)为陕西省半野生地方品种, 百粒重为7 g。两亲本粒长、粒宽等籽粒大小相关性状间存在较大差异。将该群体F6:8(2011年)和F6:9(2013年)重组自交系播种于石家庄田间, 采用3 m行长, 3行区, 随机区组试验设计, 行距50 cm, 株距10 cm。材料自然成熟后从每小区收取中间行中间10株。
自每家系籽粒中随机选取10粒, 采用数显游标卡尺测量粒长(seed length, SL)、粒宽(seed width, SW)、粒厚(seed thickness, ST), 长宽比(seed length- to-width, SLW) = 粒长/粒宽、长厚比(seed length-to- thickness, SLT) = 粒长/粒厚、宽厚比(seed width-to- thickness, SWT) = 粒宽/粒厚[21], 重复2次, 以平均数为观测值。
采用SPSS 17.0软件中相应程序对各性状进行方差、相关性和偏相关等分析。
广义遗传力h2= σ 2g/(σ 2g+σ 2ge/e+σ 2e/re) × 100%, 其中σ 2g表示基因型方差, σ 2ge表示基因型与环境互作的方差, σ 2e为误差项, e为环境数, re为重复次数[30, 31]。
在陈强[32]和雷雅坤等[33]分子遗传连锁图谱的基础上, 采用WinQTLCart 2.5[34]中的复合区间作图法(CIM)检测每个性状主效QTL位点, 选择模型6, 向前回归方法, 选择被检区间每侧10 cM的控制窗口, 进行1000次排列获得QTL显著的LOD阈值(P< 0.05)。利用QTLNetwork 2.1软件中基于混合线性模型的复合区间作图法(MCIM)分析QTL 之间的互作效应[35, 36], 将Permutation次数设置为1000次, 用于计算F值。以1 cM的步长对全基因组进行QTL扫描, 选择结果中P值显著的互作确定上位性互作及环境互作QTL。以IciMapping 4.1软件利用MET模块中ICIM-ADD和ICIM-EPI进行多环境联合分析, 以ICIM法进行上位性QTL定位, 常用LOD值为2.0, 由于本研究包含2个环境, 再结合定位结果, 故选取LOD值为4.0, 超过阈值就认为存在QTL[37]。同一性状的LOD峰值相距在5 cM以内者视为同一QTL。采取McCouch等[38]的方式命名QTL。
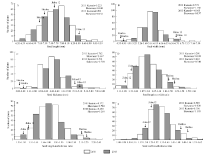
两年的表型数据分析结果(表1)显示, 冀豆12的粒长、粒宽和粒厚高于黑豆, 而黑豆的长宽比、长厚比和宽厚比高于冀豆12。群体中粒形相关6个性状均存在较大程度分离, 变异系数介于5.30%~ 8.71%之间, 且各性状均存在超双亲分离现象。各性状遗传力介于64.01%~79.57%之间。基因型与环境间的双因素方差分析结果显示, 粒长、粒宽和宽厚比受基因型、环境和基因型× 环境的影响均极显著(P< 0.01); 粒宽受基因型的影响极显著(P< 0.01); 长宽比受基因型和环境的影响均极显著(P< 0.01); 长厚比受基因型和环境的影响极显著(P< 0.01), 而基因与环境互作的影响显著(P< 0.05)。各性状的表型值均连续变异, 且为正态分布(图1)。
| 表1 亲本及群体籽粒大小与粒形性状在2011和2013年中的表现 Table 1 Characteristics of seed size and shape traits in RILs derived from the cross between Jidou 12 and Heidou in 2011 and 2013 |
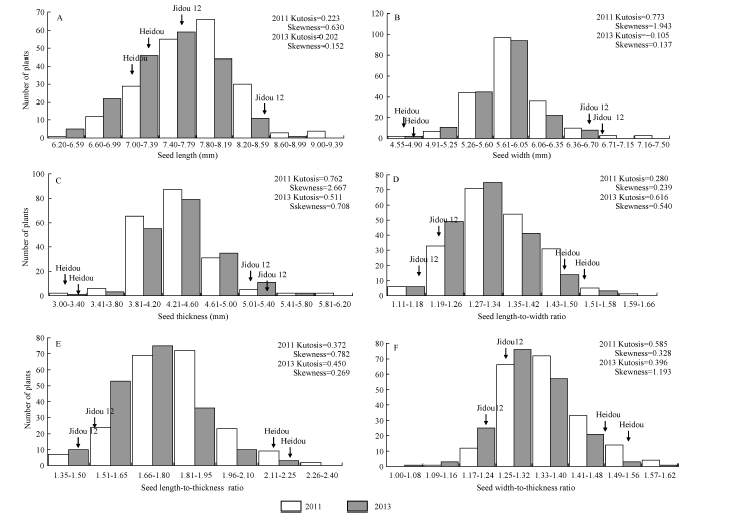 | 图1 群体粒形相关性状频率分布图 A: 群体粒长表型分布; B: 群体粒宽表型分布; C: 群体粒厚表型分布; D: 群体长宽比表型分布; E: 群体长厚比表型分布; F: 群体宽厚比表型分布Fig. 1 Frequency distribution of seed size traits of soybean populations in 2011 and 2013 A: frequency distribution of seed length; B: frequency distribution of seed width; C: frequency distribution of seed thickness; D: frequency distribution of seed length-to-width ratio; E: frequency distribution of seed length-to-thickness ratio; F: frequency distribution of seed width-to-thickness ratio. |
偏相关分析结果显示(表2), 粒长与粒宽、长宽比间均显著正相关, 与粒厚极显著负相关, 与长厚比显著负相关, 与宽厚比在2011年相关性不显著, 2013年极显著负相关。粒宽与粒厚、宽厚比极显著正相关, 与长宽比极显著负相关; 与长厚比在2011年相关性不显著, 2013年极显著负相关。粒厚与长宽比极显著正相关, 与宽厚比极显著负相关, 与长宽比相关性不显著。长宽比与长厚比极显著正相关, 与宽厚比极显著负相关。长宽比与长厚比间为极显著正相关。粒长与宽厚比2011年简单相关分析为极显著正相关, 偏相关分析无显著相关性, 2013年简单相关无显著相关性, 偏相关分析为极显著负相关。性状间相关性不稳定, 表明该相关受其他性状的影响较大。
| 表2 两年中粒形性状间的简单相关和偏相关分析 Table 2 Simple and partial correlation coefficients for seed traits in soybean in two years |
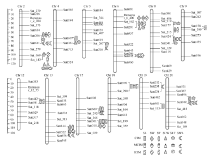
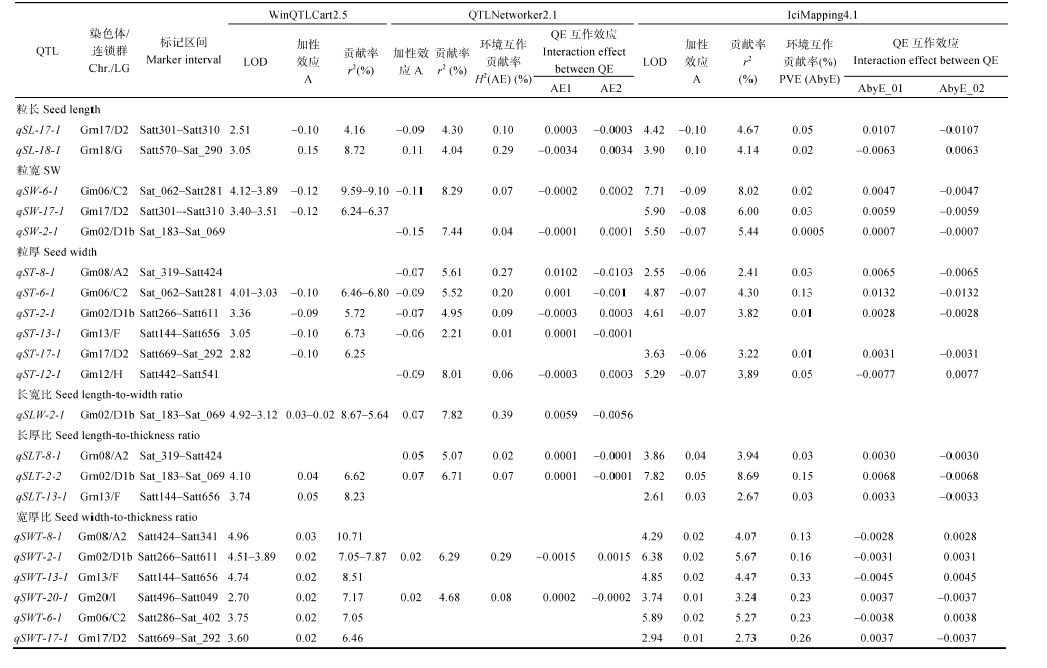
WinQTLCart 2.5和QTLNetwork 2.1进行1000次排列获得QTL显著的LOD阈值均为2.5。两年中6性状共检测到38个主效应QTL, 分布在12条染色体中(图2), 单个QTL的贡献率介于2.21%~10.71%之间。24个QTL能在2种及以上的方法中检测到, 其中8个QTL可被3种方法同时检测到(表3)。
 | 图2 检测到的主效QTL在连锁群中的分布Fig. 2 Distribution of main-effect QTLs for seed shape traits on linkage groups |
| 表3 不同方法同时检测到的大豆粒形性状加OTL及QE互作效应 Table 3 QTL for seed shape tratis in soybean |
检测到4个控制粒长的加性QTL, 贡献率范围为4.04%~8.76%。其中qSL-17-1和qSL-18-1可被3种方法同时检测到, 贡献率最大分别为4.67%和8.72%, 加性效应最大分别为-0.10 mm和0.15 mm, 增效基因分别来自于冀豆12和黑豆, 与环境互作的贡献率最大分别为0.10%和0.29%。检测到3个控制粒宽的加性QTL, 其中qSW-6-1在3种方法中均能被检测到, 贡献率最大为9.59%, 加性效应最大值为-0.12 mm, 与环境互作的贡献率最大值为0.02%; qSW-17-1和qSW-2-1分别被两种方法检测到, 贡献率最大分别为6.37%和7.44%, 加性效应值为-0.12 mm和-0.15 mm, 环境互作贡献率分别为0.03%和0.04%。检测到7个控制粒厚的加性QTL, 其中qST-6-1和qST-2-1在3种方法中可同时被检测到, 贡献率最大分别为6.80%和5.72%, 加性效应值最大分别为-0.10 mm和-0.09 mm, 环境互作贡献率最大值分别为0.20%和0.09%; qST-8-1、qST-12-1、qST-13-1和qST-17-1同时被2种方法检测到, 贡献率最大分别为5.61%、8.01%、6.73%和6.25%, 加性效应最大分别为-0.07、-0.09、-0.10和-0.10 mm, 与环境互作贡献率范围为0.01%~0.27%。
检测到3个控制长宽比的加性QTL, 其中qSLW-2-1在两种方法中同时被检测到, 贡献率最大为8.67%, 加性效应最大分别为0.07, 与环境互作贡献率为0.39%。共检测到9个控制长厚比的加性QTL, 其中qSLT-2-2在3种方法中同时被检测到, 贡献率最大为8.69%, 加性效应最大为0.07, 与环境互作贡献率为0.15%。qSLT-8-1和qSLT-13-1分别在两种方法中检测到, 贡献率最大分别为5.07%和8.23%。共检测到8个控制宽厚比的加性QTL, qSWT-2-1和qSWT-20-1可同时在3种方法中被检测到, 贡献率最大分别为7.87%和7.17%, 加性效应均为0.02, 与环境互作贡献率分别为0.29%和0.23%; qSWT-6-1、qSWT- 8-1、qSWT-13-1和qSWT-17-1可分别被两种方法检测到, 贡献率分别为7.05%、10.71%、8.51%和6.46%。
共检测到7对上位性互作QTL, 分别涉及粒长、粒宽、长宽比、长厚比和宽厚比(表4)。粒长上位性互作QTL发生在非主效QTL间, 贡献率为6.20%; 粒宽和长宽比上位性互作均发生在主效QTL与非主效QTL间, 贡献率分别为4.58%和4.06%, 效应值分别为-0.10 mm和-0.02, 表明该互作为亲本型大于重组型; 长厚比中检测到2对发生在主效QTL间的上位性互作, 为qSLT-13-1分别与qSLT-6-1和qSLT-2-2发生的互作, 贡献率分别为1.41%和0.94%, 效应值均为0.02。宽厚比上位性QTL为qSWT-20-1分别与qSWT-8-1和qSWT-13-3互作, 效应值均为0.01, 但效应相反。
| 表4 粒形性状上位互作效应QTL Table 4 Epistatie effects QTL for seed shape traits |
在QTL定位研究中, 前人建议使用多种作图法, 并优先标定共同发现的QTL [24, 25]。本研究采用的3种作图方法, 共检测到籽粒大小和粒形相关性状的加性效应QTL 38个, 其中3种方法同时检测到的QTL有8个, 分别是粒长QTLqSL-17-1和 qSL-18-1, 粒宽QTLqSW-6-1, 粒厚QTLqST-2-1和qST-6-1, 长厚比QTLqSLT-2-2, 宽厚比QTLqSWT-2-1和qSWT-20-1。参照Song等[39]的大豆公共图谱标记信息与前人研究结果比较发现, 以上8个位点中有5个与前人报道基本一致。本研究检测到的位于第18染色体的粒长QTL, 标记区间为Satt570-Sat_290, 与Moongkanna等[10]检测到粒长QTL区间Satt303-Sat_ 315部分重叠; 位于第17染色体的粒长QTL标记区间为Satt301-Satt310, 与Yu等[18]检测到粒长QTL区间Satt266-Sat_354部分重叠。本研究检测到的位于第6染色体的粒宽、粒厚QTL标记区间为Sat_062-Satt28, 与Yu等[18]检测到粒宽QTL区间Satt640-Satt422重叠, 且Xie等[16]在该区段内对粒长、粒宽和粒厚精细定位分析, 并分别检测到关联标记, Salas等[11]在该区段内同样检测到粒宽和粒厚QTL。本研究检测到的位于第20染色体的宽厚比QTL标记区间为Satt496-Satt049, 与Yu等[18]检测到粒宽QTL区间Satt419-Satt354部分重叠。该结果表明多种方法同时检测到的位点有着较高的准确性。而对于粒厚QTLqST-2-1、长厚比QTLqSLW-2-2、宽厚比QTLqSWT-2-1则未见报道。
本研究用2种方法检测到的QTL有14个, 仅用1种方法检测到的QTL有14个。其中在第4染色体中的粒长QTLqSL-4-1标记区间为Satt607-Satt361, Niu等[21]检测到的粒长相关联的标记AW277661位于该区段内; 在第8染色体中粒厚QTLqST-8-2标记区间Satt424-Satt341内同样含有Niu等[21]检测到的粒厚相关联的标记AW132402; 在第5染色体长厚比和宽厚比的QTLqSLT-5-1和qSWT-5-1标记区间Sat_267-Satt200, 与Salas等[11]检测到的粒形QTL标记区间A975_1~K636_2部分重叠。
本研究中检测到的位于在第2染色体Satt266- satt611区间、第2染色体Sat_069-Sat_183区间、第12染色体Satt422-Satt541区间、第13染色体Satt144-Satt656区间和第17染色体Satt669-Sat_292区间, 共检测到13个QTL, 均未见前人报道。
本研究检测到的38个QTL集中分布在17个标记区间内, 其中13个标记区段存在同时与多个性状相关的现象。前人研究表明, 作物相关性状QTL的集中分布是一种普遍现象, 有不少QTL可能具有多效性。Mansur等[40]认为与性状间的高度遗传力和遗传相关一致, 可能是一个上位基因作用的结果, 也可能是控制这些性状的基因紧密排列所致。在水稻中, 杜景红等[41]应用剩余杂合体衍生群体, 在初定位基础上, 把水稻第6染色体短臂上控制水稻谷壳硅含量的qHUS-6分解为qHUS6-1和qHUS6-2, 并进一步将qHUS6-2分解为一个效应较大的qHUS6-2a和效应较小的qHUS6-2b。在大豆中, 牛远等[14, 15]通过构建剩余杂合系和重组自交系, 将O染色体Satt331-Satt592 区间剖分为与标记S21-S22和O23~O19/O19~O21关联的2个多效性QTL。Xie等[16]对第6染色体Satt640-Satt422间的粒形性状效应分析, 分别检测到粒长、粒宽和粒厚相关联的SNP标记。梁慧珍等[22]对粒形性状进行QTL定位分析, 同样发现同一区段同时与多个性状相关的现象, 通过对不同性状间增效基因的来源对区段的多效性进行了初步推测。
本研究第2染色体Sat_069-Sat_183区段和第6染色体Sat_062-Satt281区段均同时与粒宽、粒厚、长宽比和长厚比相关, 第4染色体Satt607-Satt361区段同时与粒长、长宽比和长厚比相关, 第13染色体Satt144-Satt656区段同时与粒厚、长厚比和宽厚比相关。以上4个区段内的QTL, 对于籽粒大小性状(粒长、粒宽或粒厚)增效基因来自冀豆12, 而对于粒形性状(长宽比、长厚比或宽厚比)则相反, 推测其可能为多个QTL的聚合。位于第17染色体Satt301-Satt310区段同时与粒长和粒宽相关, 且增效基因均来自冀豆12, 推测其可能为多效性QTL。具体是一个主效基因还是由多个QTL 紧密连锁的聚合体还需进一步研究。
共检测到籽粒大小和粒形相关性状中加性效应QTL 38个, 以3种方法同时检测到的位点8个, 分别为qSL-17-1、qSL-18-1、qSW-6-1、qST-2-1、qST-6-1、qSLT-2-2、qSWT-2-1和qSWT-20-1, 可靠性较高, 其中qST-2-1、qSLT-2-2和qSWT-2-1为首次报道。环境互作效应贡献率均较低, 而上位性互作位点7对, 效应相对较高, 其中对粒形性状影响显著, 不能忽略。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|