




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
海岛棉转录因子基因 GbMYB60的克隆、表达及其抗逆性分析
[高巍* , 刘会利* , 田新权, 张慧, 宋洁, 杨勇, 宋纯鹏*  ]
]
]
|
|


第一作者联系方式: E-mail: gaowei021@163.com *同等贡献(Contributed equally to this work)
MYB转录因子是在真核生物细胞内广泛存在的一类转录因子蛋白, 在植物的生长发育、代谢调节、抗病抗逆、以及激素介导的信号路径等方面都发挥着重要的作用。本研究从海岛棉品种海7124中克隆得到一个MYB转录因子基因, 根据序列同源性和进化分析, 将其命名为 GbMYB60。该基因序列长990 bp, 编码一个36.9 kD的R2R3类MYB转录因子蛋白。GbMYB60蛋白被特异地定位于植物细胞核。 GbMYB60基因的表达水平较低, 在真叶中优势表达, 根系中表达量最低。该基因受甘露醇、NaCl、低温、高温等非生物逆境, 以及脱落酸、乙烯利、茉莉酸甲酯和水杨酸等植物激素的诱导上调表达。利用病毒诱导的基因沉默技术在海岛棉中干涉 GbMYB60发现, 降低 GbMYB60基因的表达水平, 使棉花幼苗对高盐胁迫的耐受性降低; 但 GbMYB60干涉的植株在甘露醇溶液的处理下与对照植株并无显著的抗性差异。
MYB transcription factors are conserved proteins in all eukaryotic cell, which play important roles in plant growth, development, metabolism, abiotic and biotic stress resistance, as well as phytohormone-mediated signal transduction. In this research, a MYB gene was isolated from the sea-island cotton cultivar Hai 7124. This gene was named GbMYB60 based on the sequence similarity search and phylogenetic analysis. The full-length of GbMYB60 coding sequence is 990 bp, and GbMYB60 encodes a 36.9 kD R2R3-type MYB protein, which is specifically located in nucleus of plant cell. GbMYB60 was preferentially expressed in leaf and induced by abiotic stresses (such as salt, mannitol, cold, and heat) and phytohormones (abscisic acid, ethephon, methyl jasmonate, and salicylic acid) treatments, but the general expression of GbMYB60 was low in all tissues. Salt and mannitol tolerances were analyzed in control and GbMYB60silenced-cotton generated by virus-induced gene silencing system, and the results showed that GbMYB60 positively regulated cotton tolerance to salt but not to mannitol.
MYB转录因子是在真核生物细胞内广泛存在的一类转录因子蛋白, 通过调节下游信号传导基因和功能基因的转录水平来实现对细胞进程的调控[1]。植物的MYB家族成员可以根据其N端MYB保守结构域的氨基酸构成被分成4类(1R、R2R3、3R和4R), 其中R2R3是植物特有的, 也是数量最多的。在拟南芥中, 目前报道的MYB转录因子有200个, 其中R2R3-MYB的数量为190个[2]。研究表明R2R3-MYB参与了植物生长发育、代谢、激素信号传导、抗逆、抗病等重要的生物学过程[1, 2, 3]。例如, 拟南芥AtMYB2在干旱胁迫下表达量上升, 通过激活脱落酸(abscisic acid, ABA)介导的信号途径增强植物的抗旱性。AtMYB2还影响了拟南芥腋芽发育和衰老过程[4, 5]。AtMYB60和AtMYB96也参与了拟南芥的抗逆反应[6, 7, 8]。AtMYB60是一个耐旱的负调控因子, 在气孔中特异表达, 表达水平被干旱胁迫抑制。atmyb60突变体气孔开度小于野生型, 因此突变体植株在缺水环境中水分散失速率明显下降, 耐旱性显著提高。而超表达MYB60.1和MYB60.2的拟南芥对干旱更加敏感[6, 8]。AtMYB96同时参与了拟南芥的抗旱和抗病过程, 促进干旱胁迫下表皮蜡质合成、水杨酸(salicylic acid, SA)和ABA介导的抗病信号传递[7, 9]。除了拟南芥, 其他物种如水稻、玉米中也发现了大量的R2R3类MYB成员[10, 11], 但关于棉花中R2R3-MYB转录因子深入的功能研究还较少。
随着棉花基因组测序的完成[12, 13], 棉花中预测有200多个MYB转录因子, 但大部分成员还没有功能报道, 只有少数成员被证明参与了棉花重要农艺性状如纤维品质、抗性的调控[14, 15, 16, 17, 18, 19]。例如, 海岛棉GbMYB5基因正调控植物对干旱的适应性反应, 干涉GbMYB5的表达降低了植株在干旱胁迫下脯氨酸和抗氧化酶的含量, 细胞内的丙二醛含量增加, 进而使得干涉植株在缺水环境下的死亡率上升。GbMYB5可能通过参与ABA信号路径和影响抗旱相关基因NCED3、RD22和RD26的表达来调节棉花的耐旱性[19]。近几年, 一些研究证实MYB基因参与了棉纤维发育过程。研究者分别在陆地棉中克隆到与拟南芥表皮毛发育相关基因GL1同源的GhMYB109、在纤维起始时期优势表达的GhMYB25和GhMYB25- like, 这些R2R3-MYB基因在纤维原始细胞(突起于胚珠表皮细胞)起始和纤维伸长过程中起重要作用[14, 15, 16, 17]。
棉花遗传转化的方法中, 应用最广泛的是农杆菌介导的稳定遗传转化[20]。然而, 此方法转化效率较低、周期长, 限制了棉花MYB基因功能的研究。病毒诱导的基因沉默(Virus-induced gene silencing, VIGS)技术在棉花中的成功运用能够部分弥补稳定转化在基因功能分析中的限制[21]。VIGS周期短、转化效率高。其中烟草脆裂病毒TRV (tobacco rattle virus)类的VIGS载体适用于棉花幼苗期的基因功能分析[21, 22], 应用此方法可以在棉花中有效、快速地开展MYB转录因子的功能研究, 为后续稳定转化棉花并深入解析其分子功能提供理论基础。本研究克隆了海岛棉中的MYB转录因子基因GbMYB60, 分析了GbMYB60的表达模式、蛋白定位及GbMYB60干涉棉花植株对非生物胁迫的响应。为GbMYB60后续的功能研究提供依据, 为棉花抗逆育种提供理论基础。
1.1.1 植物材料的种植和处理 海岛棉(Gossypium barbadense L.)品种海7124种子由南京农业大学提供, 并在海南三亚棉花繁殖基地繁种。本氏烟(Nicotiana benthamiana)由本实验室保存。棉花及烟草种子播种于营养土, 并于植物光照培箱中生长, 温度保持25℃, 光照周期为16 h光照, 8 h黑暗。
选择二叶一心时期的棉花植株, 收集根、茎、子叶和真叶的鲜样, 用于组织表达模式分析。选择长势一致的棉花幼苗, 于非生物逆境胁迫和植物激素处理后3 h取棉花叶片用于逆境和激素响应分析。盐和甘露醇处理是用300 mmol L-1的NaCl溶液和甘露醇溶液浇灌幼苗根系; 低温和高温处理是将幼苗放置到4℃和37℃的生长箱中; 伤口处理是用剪刀对待取样的幼苗叶片进行机械损伤处理; 植物激素处理是用100 μ mol L-1 ABA、200 μ mol L-1乙烯利(ethephon, ETH)、100 μ mol L-1 H2O2、100 µ mol L-1茉莉酸甲酯(methyl jasmonate, MeJA)和1 mmol L-1 SA分别对植株地上部分喷施, 水处理为平行对照。
1.1.2 载体和菌株 棉花VIGS实验载体pTRV1、pTRV2由荷兰瓦赫宁根大学Bart P. H. J. Thomma教授惠赠。Gateway系列载体pDONOR221和pK7FWGF2, 0分别从美国Invitrogen公司和比利时VIB研究所购得; TA克隆载体pGEM-T购自Promega公司。实验中所用大肠杆菌(Escherichia coli)菌株DH5α 、根癌农杆菌(Agrobacterium tumefaciens)菌株GV3101均由本实验室保存。
1.1.3 试验试剂 构建VIGS载体所用限制性内切酶购于NEB公司; Gateway载体构建所需要的BP酶、LR酶购于美国Invitrogen公司。质粒提取试剂盒、PCR产物回收试剂盒和DNA聚合酶购于北京天根生物公司; DNA测序和引物序列合成由生工生物工程(上海)股份有限公司完成; 植物激素购于索莱宝生物公司; RNA提取试剂盒购于北京艾德莱生物公司; 反转录试剂盒购于日本TOYOBO公司; qRT-PCR用的SYBR Green购于南京诺唯赞生物公司。
采用EASYspin Plus植物RNA快速提取试剂盒, 按说明书提取棉花总RNA。取1 μ g总RNA, 用ReverTra Ace qPCR RT试剂盒进行反转录。首先将总RNA在65℃变性5 min, 并立即于冰上冷却。反转录体系含5倍RT缓冲液2 μ L、RT Mix 0.5 μ L、引物0.5 μ L、变性RNA 1 μ g, 用无核酸污染的ddH2O补足到10 μ L。反转录程序为37℃ 15 min, 98℃ 5 min。反转录完成后稀释50倍用于RT-PCR和qRT-PCR。
在海岛棉基因组数据库(http://cotton.cropdb.org/ cotton/download/data.php)中搜索获得GbMYB60基因的CDS序列, 并据此设计基因特异引物M60O-F/R (表1), 以海7124幼苗叶片cDNA为模板, 用pfu DNA聚合酶进行高保真扩增。PCR反应体系含10倍pfu缓冲液5 μ L、dNTPs (10 mmol L-1) 1 μ L、M60O-F/R (10 μ mol L-1)各0.5 μ L、pfu DNA聚合酶1 μ L、用ddH2O补足至50 μ L。PCR反应程序为95℃ 5 min; 95℃ 30 s, 59℃ 30 s, 72℃ 2 min, 30个循环; 72℃ 10 min。将目标基因的PCR产物连接到TA克隆载体pGEM-T上, 并转化大肠杆菌DH5α 。比对测序后的片段与转录组中获得的片段, 序列一致者为同一基因。使用软件Primer Premier 5.0设计引物。
| 表1 本文中所涉及的引物 Table 1 Primers used in this study |
从NCBI上下载拟南芥、葡萄和棉花R2R3类MYB转录因子的氨基酸序列, 用MEGA6软件对GbMYB60及R2R3类MYB家族的其他成员进行系统进化分析, 分析方法为近邻结合法(Neighbor-Joining), 设置1000次BootStraps进行评估。多序列比对所使用的软件为DNAMAN。
用Gateway系统构建融合表达载体。经BP、LR两步反应将目标片段分别连接到中间载体pDONOR221和目标载体pK7FWG2, 0上, 获得融合表达载体p35S-GbMYB60::GFP, 并通过电击转化导入农杆菌菌株GV3101中。BP和LR反应的操作步骤见BP Clonase Enzyme Mix和LR Clonase EnzymeMix试剂说明书。
根据GbMYB60的CDS序列设计引物M60V-F/R (表1), 构建VIGS载体, 具体操作方法参考Gao等[22]。将构建的VIGS载体pTRV-MYB60和空载体对照pTRV2通过电击转化导入农杆菌GV3101中, 保存阳性克隆备用。
将含有p35S-GbMYB60::GFP质粒的农杆菌从叶背面注射到幼嫩的本氏烟叶片中, 含p35S-GFP的农杆菌为阳性对照。叶片避光处理48 h后, 用激光共聚焦显微镜(Bio-Rad Radiance 2100, 美国)观察绿色荧光蛋白的表达。
RT-PCR反应引物为M60RT-F/R (扩增32个循环)和UBI7-F/R (扩增26个循环)。每次VIGS注射后, 都应通过RT-PCR对干涉棉花植株进行基因表达量检测, RT-PCR实验至少重复4次以上。用SYBR Green染料法, 在ABI7500 Fast (Applied Biosystems, 美国)上进行qRT-PCR扩增。反应使用的酶为AceQ qPCR SYBR Green Master Mix, 基因特异引物为M60Q-F/R, UBI7-F/R为内参基因扩增引物(表1)。GbMYB60干涉效率检测qRT-PCR的实验重复设置与RT-PCR的实验重复一样。组织表达及逆境表达部分GbMYB60基因的qRT-PCR实验进行3次生物学重复, 每次重复4个技术重复。目标基因在模板中的相对变化量计算方法为2-∆ Ct, 其中∆ Ct = (CT, Targe-CT, Actin)Time x[23]。
在子叶平展时期选择长势一致的棉花幼苗进行VIGS注射, 具体方法参照Gao等[21]。15 d后, 分别取TRV:00和TRV:MYB60的第2片真叶提取总RNA, 检测GbMYB60的沉默效果。选择相同大小、发育一致的真叶统计离体叶片失水率, 每30 min测定一次叶片鲜重变化。叶盘处理方法参考Long等[24], 同样取发育一致的棉花真叶, 用打孔器对叶片相同部位打孔取叶盘, 挑选一致的叶盘放于培养皿中。在培养皿中加入0、100、250和400 mmol L-1甘露醇或NaCl溶液, 处理6 d之后照相。选取长势一致的TRV:00和TRV:MYB60植株进行NaCl和甘露醇溶液浇灌处理, 处理浓度为400 mmol L-1。处理之后每天观察植株的萎蔫反应, 并统计萎蔫叶片和失绿叶片数量。
2.1GbMYB60基因的克隆及序列分析
根据本实验室研究棉花响应非生物胁迫的RNA-Seq分析结果, 获得一条在盐胁迫处理后表达丰度上升的EST片段。在海7124中扩增到一个CDS
(coding sequence)长度为990 bp的基因, 该基因包含与前期分析发现的EST片段完全一致的区段, 编码一个长度为329 AA的蛋白(图1)。在线分析软件(http://web.expasy.org/compute_pi/)预测显示该蛋白的分子量(Mw)为36.9 kD, 等电点(pI)为5.67。
 | 图1 GbMYB60与同源蛋白的序列比对 完全一致的氨基酸残基序列用黑色背景标注, 保守的R2和R3结构域用方框标注。Fig. 1 Multiple sequence alignments of GbMYB60 and homologous proteins Identical amino acid residues are shaded in black, conserved domains R2 and R3 are boxed. |
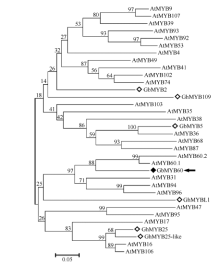
在NCBI网站用protein blast (http://blast.ncbi. nlm.nih.gov/Blast.cgi)分析蛋白的保守结构域, 发现该蛋白在靠近N端的部位有2个DNA结合区域, 可能为一个转录因子蛋白。这2个DNA结合区域的序列内部存在MYB类转录因子的典型特征。用tBlastx分析的结果显示, 该基因编码的蛋白与拟南芥AtMYB60.1 (GI:15223917)、葡萄VvMYB60 (GI:526117436)的相似性较高, 分别为52.6%和55.4%, 因此该基因被命名为GbMYB60。氨基酸序列比对分析表明, GbMYB60、AtMYB60.1、VvMYB60、GbMYB5 (GI:333101822)、GhMYB25 (GI:38456066)、GhMYB25-like (GI:326631704)和GhMYB109 (GI:38091115)在N端的120 AA比较保守, 且都含有并且仅有R2和R3两个MYB类DNA结合区域, 该区段为MYB转录因子蛋白的保守区段。C端序列的差异性相对较大, 说明C端为该蛋白的特异区段(图1)。对GbMYB60和部分拟南芥R2R3类MYB成员的氨基酸序列采用邻接法(N-J)构建进化树, 发现GbMYB60与棉花中已报道的GbMYB2、GbMYB5、GhMYB25、GhMYB25-like、GhMYB109和GbMYBL1遗传距离较远, 与AtMYB60.1、AtMYB60.2的遗传距离最近(图2)。
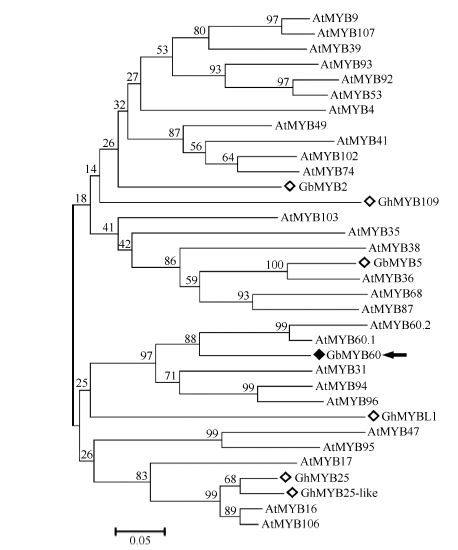 | 图2 GbMYB60与拟南芥R2R3-MYB转录因子的进化树分析Fig. 2 Phylogenetic tree of GbMYB60 and R2R3-MYB transcription factors in Arabidopsis |
利用在线软件ProtComp 9.0 (http://linux1.soft
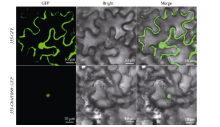
berry.com/berry.phtml)预测GbMYB60定位在细胞核。并构建C端融合GFP的融合表达载体p35S- GbMYB60::GFP转化烟草, 48 h之后用激光共聚焦显微镜观察叶片表皮细胞。结果显示, GFP单独存在时, 能在表皮细胞的细胞膜、细胞质、细胞核观察到荧光的存在; 而GFP融合GbMYB60蛋白共同表达时, 只能在细胞核里检测到绿色荧光(图3)。说明GbMYB60蛋白定位在细胞核, 符合亚细胞定位预
测的结果和转录因子蛋白通常存在的定位情况。
 | 图3 GbMYB60蛋白的亚细胞定位Fig. 3 Subcellular localization of GbMYB60 protein |
利用在线工具PlantCARE (http://bioinformatics. psb.ugent.be/webtools/plantcare/html/)对GbMYB60转录起始位点上游2 kb的DNA序列预测发现, 在GbMYB60的启动子区存在多种与生长发育、代谢调控、抗病、抗逆和激素响应相关的顺式作用元件(表2)。其中包括与干旱相关的MBS、响应高温胁迫的HSE、逆境胁迫和防卫反应相关的TC-rich repeats、真菌激发子诱导相关的Box-W1、启动基因在地上部分特异表达的as-2-box、ABA响应元件ABRE和乙烯响应元件ERE等。说明GbMYB60在转录水平受多种因素影响, 在植物细胞中可能参与激素调控和非激素介导的胁迫信号传导和响应过程。
| 表2 预测的GbMYB60启动子区段部分顺式元件 Table 2 Part of the predicated cis-elements in GbMYB60 promoter region |
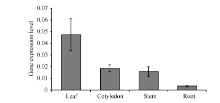
利用qRT-PCR技术检测GbMYB60的表达显示, GbMYB60在真叶中表达量最高, 子叶和茎中表达量次之, 在根系中表达量最低, 约为真叶中的1/20。说明GbMYB60在棉花幼苗的不同组织中转录水平呈现较大差异, 即GbMYB60的表达存在组织特异性, 在地上部分优势表达。此外, GbMYB60在棉花中的表达水平较低, 子叶和茎中的GbMYB60表达量仅为内参基因GbUB7的1/50 (图4)。
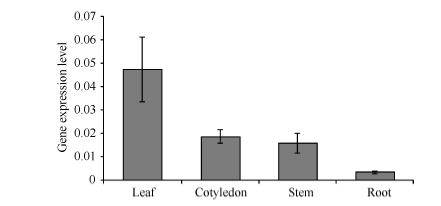 | 图4 GbMYB60在棉花幼苗真叶、子叶、茎和根中的组织表达模式Fig. 4 Tissue-specific expression of GbMYB60 in leaf, cotyledon, stem, and root of cotton seedling |
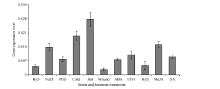
为了进一步验证启动子顺式元件预测的结果, 对真叶进行不同逆境和植物生长调节剂处理。结果显示, 在盐、甘露醇、低温和高温4种非生物胁迫以及ABA、ETH、MeJA和SA四种激素处理下, GbMYB60的表达量都有提高, 说明GbMYB60在转录水平受到这些因素的诱导。但是对真叶机械损伤和喷施H2O2时, GbMYB60的表达量维持在与对照相近的水平(图5)。
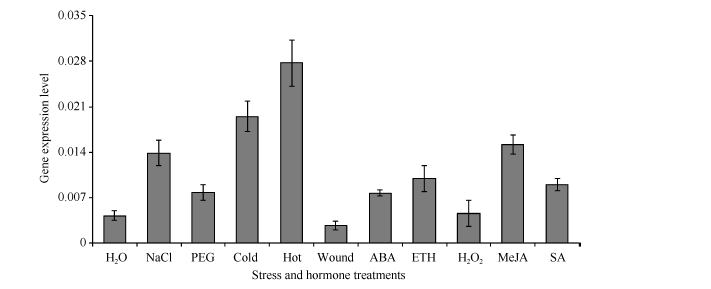 | 图5 GbMYB60基因在非生物逆境和激素诱导下的表达变化Fig. 5 GbMYB60 expression changes under abiotic stress and phytohormone treatments |
2.4.1 VIGS干涉GbMYB60棉花植株的检测 海7214在注射含VIGS载体的农杆菌2周时, GbMYB60沉默的棉花幼苗TRV:MYB60地上部分的外部特征, 包括株高、叶片大小等, 与空载体转化的对照植株TRV:00无明显区别(图6-A)。用RT-PCR和qRT-PCR检测植株中GbMYB60基因的干涉效率, 发现TRV:MYB60中GbMYB60的表达量仅为对照TRV:00的30%, 为同条件下内参基因GbUB7转录水平的1% (图6-B)。说明利用VIGS技术, 能显著降低海7124中GbMYB60基因的表达水平, 干涉GbMYB60不影响海7124棉花幼苗的生长发育。
 | 图6 利用VIGS技术沉默棉花幼苗中的GbMYB60基因 A: 空载体转化的对照材料TRV:00和GbMYB60干涉材料TRV:MYB60; B: qRT-PCR和RT-PCR检测棉花叶片中GbMYB60的干涉效率(* P< 0.01, t检验)。Fig. 6 Silencing of the GbMYB60 gene in cotton seedlingsusing VIGS A: Representative seedlings of TRV:00 (empty control) and TRV:MYB60 (GbMYB60- silencing); B: qRT-PCR and RT-PCR of GbMYB60 expression level in cotton leaf (* P< 0.01, t-test). |
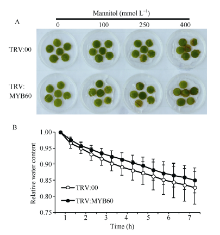
2.4.2 干涉GbMYB60不影响棉花对甘露醇的耐受性 图7-A所示, 选择TRV:00和TRV:MYB60同一发育时期的叶片, 取叶盘浸泡在不同浓度的甘露醇溶液中, 观察叶盘的变化。处理6 d后, 水处理的对照组中叶盘没有明显变化, 100、250和400 mmol L-1甘露醇处理的叶盘边缘均出现不同程度的失绿和褐化, 但TRV:00和TRV:MYB60之间的失绿、褐化程度没有差异。结果显示, TRV:00和TRV:MYB60的离体叶片在自然条件下水分散失的速率在数据统计上没有显著区别(图7-B); 浇灌400 mmol L-1甘露醇处理12 d后, TRV:00和TRV:MYB60植株均出现下端叶片黄化、萎蔫的现象, 但上部的叶片仍然保持绿色, 并未受甘露醇影响, 且TRV:00和TRV:MYB60黄化、萎蔫叶片的数目和萎蔫程度也未见差异(图8)。说明在海7124中干涉GbMYB60基因并未影响棉花对高浓度甘露醇溶液的耐受性和离体叶片的失水速率。
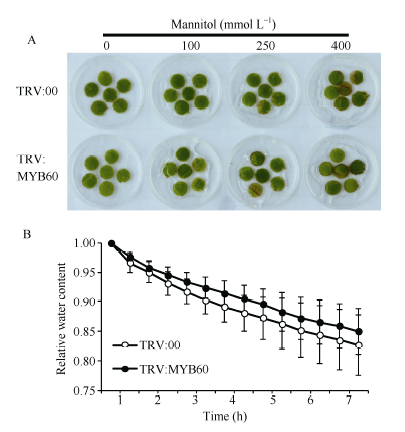 | 图7 TRV:00和TRV:MYB60叶片对甘露醇耐受性的分析 A: 0、100、250、400 mmol L-1甘露醇溶液处理TRV:00和TRV:MYB60叶盘; B: 离体棉花叶片的失水率测定。Fig. 7 Mannitol tolerance analysis of TRV:00 and TRV:MYB60 leaves A: Leaf disks of TRV:00 and TRV:MYB60 in 0, 100, 250, 400 mmol L-1 mannitol solutions; B: Measurement of the dehydrating rate of detached cotton leaf. |
 | 图8 经400 mmol L-1甘露醇溶液处理后TRV:00和TRV:MYB60棉花幼苗Fig. 8 TRV:00 and TRV:MYB60 seedlings treated with 400 mmol L-1 mannitol solution |
2.4.3 GbMYB60干涉植株对盐胁迫的敏感性增加
GbMYB60在盐处理下表达量显著上调, 为了分析GbMYB60是否影响棉花对盐胁迫的耐受性, 分别对TRV:00和TRV:MYB60材料的叶盘和整株植株用NaCl溶液处理。如图9所示, 处理6 d后, 除水处理的对照组外, 100、250和400 mmol L-1 NaCl溶液处理的叶盘均出现严重的失绿和褐化, 说明盐溶液对TRV:00和TRV:MYB60都造成了胁迫。100、250 mmol L-1两种浓度处理下TRV:MYB60材料受胁迫的程度略高于TRV:00, TRV:00在100 mmol L-1的NaCl溶液中有部分叶盘维持绿色, 而TRV:MYB60的叶盘都已经褐化。浇灌400 mmol L-1NaCl溶液12 d后, 棉花幼苗出现典型的盐胁迫表型, 下部叶片萎蔫现象严重(图10-A)。其中TRV:MYB60植株的萎蔫叶片占总叶片的48%, TRV:00仅为22% (图10-B)。说明在海7124中干涉GbMYB60基因增加了棉花幼苗对盐胁迫的敏感性。
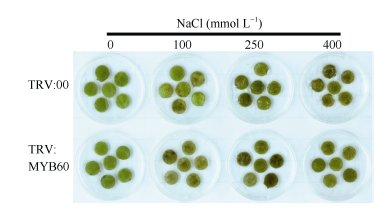 | 图9 TRV:00和TRV:MYB60叶片的耐盐性分析Fig. 9 Salt tolerance analysis of TRV:00 and TRV:MYB60 leaves |
 | 图10 TRV:00和TRV:MYB60幼苗的耐盐性分析 A: 经400 mmol L-1 NaCl溶液处理的TRV:00和TRV:MYB60棉花幼苗; B: 盐处理后植株存活叶片比率统计。Fig. 10 Salt tolerance analysis of TRV:00和TRV:MYB60 seedlings A: TRV:00 and TRV:MYB60 seedlings treated with 400 mmol L-1 NaCl solution; B: calculation of cotton leaves remaining green after salt treatment. |
MYB家族是植物中最大的转录因子家族之一, 通过N端保守的MYB核酸结合位点与启动子区特异位点相结合, 实现对下游基因的转录调控[1, 2, 3], 从而参与植物的生长发育、信号传递和对逆境协迫的响应[25, 26]。相对于拟南芥、水稻等模式植物, 棉花中MYB转录因子基因功能以及调控机制的研究主要集中在纤维发育上[14, 15, 16, 17], 抗逆相关棉花MYB转录因子的研究还很少, 克隆该家族基因并解析其功能对棉花抗逆分子机制的研究具有重要的意义。本研究从海岛棉中克隆到一个转录因子GbMYB60。通过序列比对分析发现GbMYB60在N端包含2个连续的R2、R3类MYB结构, 确定该基因为R2R3-MYB, 这2个结构的氨基酸序列与其同源蛋白AtMYB60和VvMYB60高度相似, 但在120 AA后的C端其序列长度和氨基酸组成都出现了很大的差异。MYB蛋白的C端为负责蛋白活性调节的特异区段, 决定了不同MYB成员之间的功能差异[2]。GbMYB60虽然在序列上与AtMYB60和VvMYB60同源度高, 但这3个蛋白C端活性调节区的差异说明GbMYB60在棉花中的功能与这2个蛋白已报道的功能可能不同, 参与的逆境响应和信号响应也可能存在差异。
本研究预测GbMYB60的启动子区域存在多种响应逆境胁迫和激素诱导的顺式元件。其中包括ABA响应以及干旱信号应答相关的元件ABRE和MBS[27]。qRT-PCR的结果也与预测结果一致, 在盐、甘露醇、低温、高温、ABA、ETH、MeJA、SA处理下, GbMYB60转录水平都出现了不同程度的上升, 说明GbMYB60可能参与了多种棉花抵抗非生物胁迫的响应和激素调控的信号传递过程。R2R3类MYB在植物MYB家族中数量众多, 功能各异。目前已有的大量研究工作证明R2R3-MYB参与植物初生及次生代谢、细胞分化、发育、抗病和抗逆等过程[1, 2, 3]。且同一个MYB转录因子在不同的环境中具有不同的功能, 如AtMYB60被报道响应病原菌的侵染; 在干旱胁迫下调控气孔运动; 异源表达AtMYB60影响了生菜中的花青素合成[6, 8, 28]。这些都预示着GbMYB60在棉花的逆境响应甚至生长发育中都可能行使着一定的功能。
本研究分析了GbMYB60在棉花抵御非生物胁迫中的功能, 通过对GbMYB60干涉棉花幼苗进行甘露醇和盐处理的表型实验分析, 发现海岛棉中GbMYB60的表达抑制对棉花幼苗对高浓度甘露醇溶液的耐受性、以及离体叶片的失水速率没有影响, 而对盐胁迫的敏感性增强。拟南芥的AtMYB60在叶片气孔细胞中特异表达, 而且其表达受到ABA的抑制[6, 8]。拟南芥突变体atmyb60对干旱胁迫的抗性增强, 说明AtMYB60是一个干旱耐受性的负调控因子[8], 对盐胁迫的影响目前未见报道。二倍体植物拟南芥的基因组为120 Mb, 包含11 600万个碱基对, 可能编码大约26 000个基因[29]; 海岛棉是一种异源四倍体, 基因组大小为2.5 Gb, 海岛棉基因组非常复杂, 重复序列多, 在异源四倍体中分离到的基因多数为多拷贝基因, 因此生物学过程中的基因调控网络也更复杂[12, 13]。在棉花中抑制GbMYB60的表达与同源基因AtMYB60的结果不同, 可能是由GbMYB60和AtMYB60氨基酸序列C端特异的活性调控区段, 或者是物种之间的差异造成的。高等植物为了保证其生长发育的正常运行, 进化出几个基因同时控制同一性状的现象, 即功能冗余。如拟南芥中MPK3和MPK6共同调控拟南芥抗病途径的Priming效应, 同时突变MPK3和MPK6才能降低拟南芥诱导抗病性的产生[30]。鉴于GbMYB60受逆境胁迫和抗逆相关激素的诱导, 在棉花中也可能存在与GbMYB60功能冗余的基因, 共同调控棉花的抗逆性, 但是这个推论还需要在棉花中更深入的研究来验证。研究GbMYB60基因在棉花抗逆中的功能和作用机制, 可能为棉花抗性育种提供理论基础和候选基因。
从海岛棉中克隆到一个MYB转录因子基因GbMYB60, 该基因在真叶中优势表达并受NaCl、低温、高温等非生物逆境, 以及ABA、ETH、MeJA和SA等激素的诱导上调。GbMYB60蛋白定位于植物细胞核。该基因不影响植株对甘露醇的耐受性, 但正调控对盐胁迫的耐性。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|