
{kind=link}
{kind=link}
高温胁迫对Bt棉蕾中杀虫蛋白含量及氮代谢的影响
[衡丽1 , 李亚兵2 , 胡大鹏1 , 王桂霞1 , 吕春花1 , 张祥1 , 陈源1 , 陈德华1, *  ]
]
]
|
|


第一作者联系方式: E-mail: 917103441@qq.com
为明确高温气候下Bt棉抗虫性下降的原因, 以2个不同类型Bt棉品种为材料, 2011年于盛蕾期设计38℃高温胁迫3、5和7 d; 2012年设计38℃/25℃的昼夜变温胁迫4、7和10 d, 研究其对转Bt棉蕾的杀虫蛋白表达量影响及其生理机理。结果表明, 持续高温胁迫7 d内, 棉蕾中Bt杀虫蛋白含量呈下降趋势, 持续胁迫3 d内下降幅度最大, 与未胁迫相比, 泗抗3号和泗抗1号分别下降18.71%和26.54%; 昼夜变温下高温胁迫4 d时蕾中Bt杀虫蛋白表达量无显著变化, 当胁迫7 d以上时, Bt杀虫蛋白含量显著下降, 泗抗3号和泗抗1号分别比对照下降11.32%和14.18%。持续高温或昼夜变温高温持续胁迫下, 棉蕾中游离氨基酸含量和蛋白酶活性都显著增加, 可溶性蛋白含量和GPT活性都显著下降。相关分析表明, 游离氨基酸含量、蛋白酶活性与Bt杀虫蛋白含量呈极显著负相关; 可溶性蛋白含量和GPT活性与Bt杀虫蛋白含量呈极显著正相关。分析表明, 高温胁迫下蛋白质合成功能下降, 分解能力增强导致可溶蛋白含量, 包括Bt杀虫蛋白含量下降。常规种泗抗1号的抗虫性和氮代谢受高温的影响较大。
To explore the reason for low insect resistance under high temperature climate in Bt cotton production, we selected two Bt cotton cultivars as experimental material with treatments of 38℃ for three, five and seven days in 2011, and 38℃/25℃ at day /night for four, seven and ten days, respectively at the artificial climate chamber in 2012. The results showed that the Bt insecticidal protein contents declined within seven days under 38℃, with the largest reduction occurred within three days. Compared with control, the square Bt insecticidal protein contents decreased by 18.71% and 26.54% for cultivar Sikang 3 (SK-3) and Sikang 1 (SK-1), respectively. Under 38℃/25℃ at day/night, the square Bt insecticidal protein contents had no obvious reduction within four days, and significant reduced after seven days with the reduction of 11.32% for SK-3 and 14.18% for SK-1. Under the treatment conditions the soluble protein contents, glutamate pyruvate transaminase (GPT) activities reduced, but the free amino acid contents, protease activities increased. There existed significant negative correlation of insecticidal protein content with free amino acid contents, and protease activities; and significantly positive correlation of insecticidal protein content with soluble protein contents, and GPT activities under high temperature stresses. Therefore, the reduced synthesis and the enhanced degradation for protein in the square under the high temperature condition resulted in the decrease of soluble protein content, including Bt insecticidal protein content. The larger reductions of the square Bt insecticidal protein content and nitrogen metabolic strength were detected in cultivar SK-1 as confound with cultivar SK-3 under the high temperature treatments.
随着转Bt基因棉在生产上应用, 棉铃虫等相关害虫危害和农药的用量得到一定的控制[1, 2]。但其抗虫性不稳定受到了人们的密切关注[3, 4]。陈松等[5]对转Bt抗虫棉32B不同生育期抗虫性测定表明整个苗期抗虫性均较强, 现蕾期有所下降, 生长中后期显著下降至最低。沈平等[6]研究表明, 转Bt基因棉不同品种的抗虫性表现为蕾期> 盛花期> 花铃期> 铃期。李汝忠等[7]研究表明, 全展功能叶中的Bt杀虫蛋白含量最高, 根、茎和叶柄中含量较低。这种时空差异性影响Bt棉不同生育期不同部位的抗虫性的表达[8, 9, 10]。此外, Bt棉抗虫性的表达同时还受环境的影响, 其中, 温度是其重要因素之一[11, 12, 13]。陈德华等[14]报道在结铃盛期给予37℃的高温胁迫, 叶片中Bt杀虫蛋白含量和抗虫性明显下降, 夏兰芹等[15]研究表明高温使Bt基因沉默时间提前, 杀虫蛋白表达降低时间相应提前, 导致杀虫蛋白含量急剧降低。其他研究也表明高温和低温都会影响Bt棉杀虫蛋白表达量[16, 17, 18, 19], 但上述研究主要是以叶片为研究对象。由于生殖器官是棉铃虫等相关害虫首选危害目标, 因此研究Bt棉生殖器官抗虫性受高温的影响具有重要意义。研究高温对蕾抗虫性的影响有利于建立蕾期合理防治棉铃虫等相关害虫的策略。因此, 本文以棉蕾为研究对象, 探索蕾期持续高温和昼夜变温对Bt棉蕾杀虫蛋白表达量及其氮代谢特征, 明确高温条件下棉蕾抗虫性的下降程度及其原因, 为Bt棉生产中对高温逆境下棉铃虫等相关害虫的防治提供预警决策依据。
2011— 2012年在扬州大学江苏省遗传栽培生理重点实验室进行, 以Bt转基因棉常规品种泗抗1号、杂交种泗抗3号为材料(分别以代号SK-1和SK-3表示)进行盆栽试验。供试土壤为沙壤土含有机质1.88%、水解氮134.7 mg kg-1、速效磷22.5 mg kg-1、速效钾81.3 mg kg-1。试验所用盆钵直径30 cm, 高27 cm, 每盆装土11 kg, 将取自大田的土壤自然风干、过筛去杂后装盆, 用水沉实。4月7日营养钵育苗, 5月20日移栽盆中, 定期浇水, 保持每天盆中土壤含水量接近田间持水量。肥料与其他管理措施同当地高产栽培要求一致。
2011年于盛蕾期以38℃临界高温分别胁迫3、5和7 d, 以温度32℃作为对照。胁迫过程中保持空气湿度70%。在人工气候箱达到预定温湿度后, 移入盆栽棉花样本15盆, 其棉株上已标记每个蕾的现蕾日期。样本移入后开始计时, 在到达设计的胁迫时间后及时取现蕾后15 d棉蕾。每个处理3个重复。取样后即以液氮冷冻样品, 并存放在-20℃冰箱用于Bt杀虫蛋白含量及氮代谢相关物质和酶的分析。
2012年在盛蕾期设计昼夜温度38℃/25℃的白天临界高温, 夜晚适宜温度的昼夜变温胁迫, 分别持续4、7和10 d。设置湿度为70%。每个处理3个重复, 样本量及现蕾标记与2011年相同。在人工气候室达到预期温湿度后移入棉花样本。同时设对照, 温度为32℃/25℃。胁迫至预定时间后, 取15日龄棉蕾, 液氮速冻, 同样存放在-20℃冰箱用于Bt杀虫蛋白含量及氮代谢相关物质和酶的分析。
1.3.1 Bt蛋白含量 参见何忠佩所编《农作物化学控制实验指导》[20], 应用酶联免疫法(ELISA)测定, 药盒由中国农业大学提供。
1.3.2 可溶性蛋白含量 应用G-250考马斯亮蓝比色法测定[21]。
1.3.3 游离氨基酸含量 应用抗坏血酸茚三酮染色法测定[21]。
1.3.4 谷氨酸丙酮酸转氨酶(GPT)活性 经Tris-HCl缓冲液研磨离心后得到提取液, 用赖氏比色法测定[22]。
1.3.5 蛋白酶活性 应用考马斯亮蓝法测定, 进而计算蛋白酶活性。
1.3.6 数据处理 运用Microsoft Excel 2003等软件统计分析数据, 用SigmaPlot 10.0绘制图表。
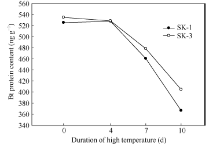
2.1.1 对Bt杀虫蛋白表达量影响 图1表明, 盛蕾期持续高温胁迫处理下, 2个类型品种棉蕾Bt蛋白含量变化动态一致, 在胁迫3 d内, 棉蕾Bt蛋白含量大幅下降, 胁迫3~7 d下降速度趋于平缓。以未胁迫时Bt蛋白含量为对照, SK-3在38℃高温胁迫3、5和7 d后, 蕾中Bt蛋白含量分别下降18.71%、25.36%和32.67%; SK-1分别下降26.54%、33.20%和42.15%。品种间相比, 常规种SK-1下降幅度较大。可见, 38℃高温持续胁迫引起棉蕾中Bt杀虫蛋白浓度下降, 且随胁迫期延长, 下降幅度加大, 但以胁迫3 d内抗虫性下降幅度最大。
 | 图1 持续高温胁迫下棉蕾的Bt杀虫蛋白表达量的变化Fig. 1 Effect of the duration of high temperature on square Bt protein contents in Bt cotton |
2.1.2 对氮代谢特征的影响 表1表明, 盛蕾期持续高温胁迫7 d内, 2个类型品种棉蕾游离氨基酸含量都随胁迫时间的延长而增加。38℃高温胁迫3 d内, 棉蕾游离氨基酸含量大幅上升, 3~7 d增加速度趋于平缓。与各自对照相比, SK-3在高温下胁迫3、5和7 d后, 蕾中Bt杀虫蛋白含量分别增加73.50%、96.39%和111.47%, SK-1分别增加74.67%、97.13% 和116.89%。常规种SK-1棉蕾游离氨基酸上升幅度更大。说明持续高温胁迫7 d引起棉蕾游离氨基酸含量大幅增加, 这显然与蛋白质分解有关。
| 表1 持续高温胁迫下棉蕾氮代谢主要物质含量和关键酶活性 Table 1 Main compound contents and key enzyme activities in nitrogen metabolism of square under high temperature stress |
盛蕾期持续高温胁迫7 d内, 2个类型品种棉蕾可溶性蛋白含量都随着胁迫期延长而下降(表1)。胁迫3 d内, 棉蕾可溶性蛋白含量下降幅度最大, 3~7 d下降速度趋于平缓。与各自对照相比, SK-3在38℃高温水平胁迫3、5和7 d, 蕾中可溶蛋白含量分别下降21.00%、28.43%和42.41%, SK-1分别下降31.35%、36.03%和45.91%, 常规种SK-1下降幅度更大。可见, 高温胁迫引起棉蕾可溶性蛋白含量下降。
表1还表明, 盛蕾期持续高温胁迫7 d内, 2个类型品种棉蕾GPT活性都随着高温胁迫期延长而下降, 尤其以高温胁迫3 d内下降幅度大。与各自对照相比, SK-3在38℃高温水平胁迫3、5和7 d, 蕾中GPT活性分别下降11.93%、15.42%和17.87%, SK-1分别下降15.80%、18.87%和20.80%。常规种SK-1中GPT活性受影响更大。说明高温胁迫影响棉蕾的氨基酸和蛋白质合成。
随着高温胁迫时间的延长, 2个类型品种棉蕾蛋白酶活性增加(表1)。胁迫3 d内, 棉蕾蛋白酶活性增加幅度最大。与各自对照相比, SK-3在38℃高温水平胁迫3、5和7 d, 蕾中蛋白酶活性分别增加41.60%、59.29%和73.37%, SK-1分别增加50.18%、69.35%和90.17%。常规种SK-1棉蕾蛋白酶活性上升幅度更大。可见, 持续高温胁迫引起棉蕾中蛋白质分解能力加强, 且持续时间越长, 分解能力也越强。
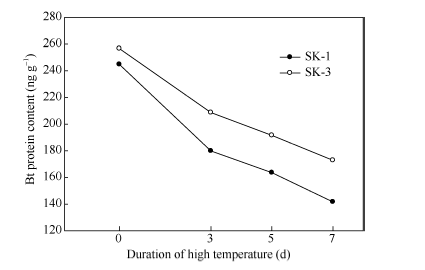
2.2.1 对Bt杀虫蛋白表达量的影响 图2表明, 与对照相比, 2个品种在昼夜变温条件下持续高温胁迫4 d时, 蕾的Bt杀虫蛋白表达量变化较小, 但胁迫至7 d时, Bt杀虫蛋白表达量均明显下降。其中, 常规种SK-1胁迫至4、7和10 d时, Bt杀虫蛋白表达量比对照分别下降0.72%、14.18%和31.50%; 杂交种SK-3分别下降0.61%、11.32%和25.21%。说明, 无论是常规种还是杂交种, 变温条件下4 d的高温胁迫, Bt杀虫蛋白表达量基本不受影响, 但持续至7 d以上时, Bt杀虫蛋白表达量显著下降, 且随着胁迫持续时间的延长, 下降的速率加快, 幅度也越大。品种间则表现为常规种的下降速度和下降幅度稍高于杂交种, 说明杂交种在昼夜变温条件下高温胁迫使蕾的抗虫性下降幅度小。
 | 图2 昼夜变温下高温对棉蕾中Bt杀虫蛋白表达量的影响Fig. 2 Effect of the high temperature under variable temperature at day/night on square Bt protein content in Bt cotton |
2.2.2 对氮代谢特征的影响 由表2可知, 与对照相比, 2个不同类型品种在昼夜变温下高温胁迫4d时, 棉蕾中GPT活性下降较小, 胁迫至7 d时, GPT活性均明显下降, 且随着胁迫时间的延长, 下降速率和幅度均增加。其中, SK-1胁迫至4、7和10 d时, GPT活性比对照分别下降3.16%、13.76%和29.90%; SK-3分别下降2.01%、10.72%和22.94%。说明, 无论是常规种还是杂交种, 昼夜变温条件下高温胁迫7 d以上时引起棉蕾GPT活性的快速下降。常规种的下降速度和下降幅度稍高于杂交种。
随昼夜变温条件下高温胁迫时间延长, 棉蕾中的游离氨基酸含量增加, SK-1在胁迫4、7和10 d时, 比对照分别增加0.79%、23.60%和49.55%; SK-3比对照分别增加0.89%、15.58%和39.54%。可见, 无论是常规种还是杂交种, 7 d以上的昼夜变温条件下高温胁迫会造成棉蕾中游离氨基酸含量快速增加, 且常规种上升幅度较大。
表2还表明, 与对照相比, 2个不同类型品种在昼夜变温条件下高温胁迫4 d时, 蕾中可溶性蛋白含量无显著变化, 但胁迫至7 d时, 可溶性蛋白含量均显著下降, 且随胁迫时间的延长, 下降幅度増大。常规种SK-1在胁迫4、7和10 d条件下, 可溶性蛋白含量分别下降0.65%、14.19%和31.64%; 杂交种SK-3分别下降0.33%、11.91%和28.85%。说明无论是常规种还是杂交种, 棉蕾中可溶性蛋白含量出现明显下降是在变温胁迫7 d以上, 且常规种的下降幅度高于杂交种。
与对照相比, 2个不同类型品种在昼夜变温条件下高温胁迫至4 d时, 棉蕾中蛋白酶活性变化较小, 胁迫至7 d时, 蛋白酶活性均明显增加, 随着胁迫时间的延长, 增长幅度变大(表2)。SK-1在胁迫至4、7和10 d时, 蕾蛋白酶活性分别增加1.81%、57.12%和111.11%; SK-3分别增加1.29%、30.79%和83.86%。说明昼高夜低变温胁迫至7 d及以上, 导致棉蕾中的蛋白酶活性快速上升, 且随胁迫时间的延长, 上升幅度和速率加大。常规种的上升速率和幅度强于杂交种。
| 表2 昼夜变温下高温对棉蕾氮代谢主要物质含量及关键酶活性的影响 Table 2 Main compound contents and key enzyme activities in nitrogen metabolism of cotton square under the variable high temperature at day/night |
表3表明, 无论是持续高温胁迫还是昼高夜低变温胁迫, 2个品种棉蕾中游离氨基酸含量、蛋白酶活性与Bt杀虫蛋白含量都存在极显著的负相关; 可溶性蛋白含量、GPT活性都与Bt杀虫蛋白含量呈极显著正相关, 进一步说明在高温胁迫下, 棉蕾GPT活性的下降, 蛋白酶活性的增加导致游离氨基酸含量增加, 可溶性蛋白含量下降, 进而引起Bt杀虫蛋白浓度下降。
| 表3 高温胁迫下蕾中Bt杀虫蛋白含量与氮代谢主要物质含量及关键酶活性的相关系数 Table 3 Correlation coefficients between square Bt protein content and main compound contents and key enzyme activities under high temperature stresses |
温度是引起Bt棉杀虫蛋白表达量变化的重要环境因素, 尤其在棉花生长季节内的极端高温[23, 16]。夏兰芹等[15]、张祥等[24]和周桂生等[25]研究均表明高温会引起棉花Bt杀虫蛋白表达量的下降, 张坤等[26]对转Bt基因棉花种子高温处理(55~130℃), 并用双抗夹心酶免疫检测方法(ELISA)研究其中的外源Bt蛋白的动态变化过程。结果表明棉花种子中Bt蛋白耐高温能力较强, 种子中的Bt蛋白在10 min内含量显著下降, 其后降解缓慢; 130℃以上的高温处理条件下, 种子中的外源Bt蛋白能显著降解。陈德华等[14, 27]研究认为, 37℃高温已引起Bt棉叶片杀虫蛋白的降解。本研究表明, 持续高温胁迫7 d内显著影响蕾的杀虫蛋白表达量。2个不同类型的Bt棉品种在盛蕾期38℃下持续胁迫7 d内棉蕾的杀虫蛋白浓度一直呈下降趋势, 尤其在3日内下降幅度更大。昼夜变温下高温胁迫则表明, 胁迫7 d以上时棉蕾Bt杀虫蛋白含量显著下降。2个品种间以品种常规种的下降幅度较大。以上结果表明, 持续高温和昼夜变温下高温引起棉蕾Bt杀虫蛋白含量下降的时间不同。在变温条件下, 需要更长的时间才会引起棉蕾Bt杀虫蛋白含量的下降, 这可能与其在夜间解除高温胁迫, 棉蕾恢复蛋白合成包括Bt杀虫蛋白合成有关[28]。由于棉花的生长常常遭受昼夜变温条件下高温气候影响[29], 因此需密切注意连续7 d以上高温气候情况下Bt棉抗虫性变化和棉铃虫等相关害虫的发生情况, 制定更有效的防控措施, 减轻虫害损失。此外, 常规种棉蕾的Bt杀虫蛋白含量受高温影响比杂交种大, 可能与杂交种对高温逆境具有更强的抵抗能力有关。
张桂玲等[30]实验表明短期盐胁迫的Bt杀虫蛋白含量与游离氨基酸含量呈极显著负相关, 与可溶性蛋白含量呈极显著正相关。王永慧等[31]得出盛铃期湿度胁迫叶片Bt杀虫蛋白含量的下降与Bt杀虫蛋白合成能力下降、分解能力增强相关。陈德华等[32, 33]通过对不同类型Bt棉品种生育过程中毒蛋白含量变化的研究表明GPT活性、可溶性蛋白含量与Bt杀虫蛋白含量密切正相关, 认为氮代谢活性影响Bt杀虫蛋白的表达。我们的研究同样表明, 持续高温胁迫和昼夜变温下高温引起可溶性蛋白含量、GPT活性显著下降, 游离氨基酸含量、蛋白酶活性明显增加。且两种胁迫条件下, 棉蕾中可溶性蛋白含量、GPT活性与Bt杀虫蛋白含量均呈极显著正相关。游离氨基酸含量、蛋白酶活性与Bt杀虫蛋白含量呈极显著负相关。说明高温胁迫下Bt棉蕾中蛋白质的分解能力增强, 合成能力减弱, 引起可溶性蛋白下降, 从而导致杀虫蛋白含量下降。因此, 高温逆境下调节Bt棉氮代谢强度, 特别是促进生殖器官中与蛋白质合成有关的酶活性的增强对于稳定和提高Bt棉抗虫性具有重要意义。
持续高温和昼夜变温下高温胁迫引起Bt棉蕾中杀虫蛋白表达量下降, 38℃高温持续胁迫3日内下降幅度最大; 38℃/25℃昼夜变温下高温胁迫持续7 d以上时, 棉蕾Bt杀虫蛋白含量显著下降; 高温胁迫下棉蕾Bt杀虫蛋白含量的变化与氮代谢密切相关。高温胁迫下引起游离氨基酸含量和蛋白酶活性升高, 可溶性蛋白含量和GPT活性下降。因此在棉花生长季节需及时注意高温气候对Bt棉蕾抗虫性的影响及程度, 防止棉铃虫等相关害虫对棉花的危害。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|