

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
[王亚梁* , 张玉屏* , 朱德峰*  , 向镜, 武辉, 陈惠哲, 张义凯]
, 向镜, 武辉, 陈惠哲, 张义凯]
, 向镜, 武辉, 陈惠哲, 张义凯]
|
|


第一作者联系方式: 王亚梁, E-mail: wangyaliang1992@hotmail.com, Tel: 18867962255; 张玉屏, E-mail: cnrrizyp@163.com, Tel: 15397092027
为明确水稻穗分化期高温胁迫对颖花退化和籽粒充实的影响, 选用耐热性品种黄华占和热敏感性品种丰两优6号进行人工气候箱盆栽试验, 于倒一叶心叶抽出时(花粉母细胞形成至减数分裂期)进行高温处理(10:00—15:00, 40℃) 1 d、3 d、5 d、7 d、9 d, 并以同时段适温处理(10:00—15:00, 32℃)为参考, 自然条件下生长的植株为对照, 分析不同持续天数高温对颖花退化、花器官发育和籽粒充实的影响。结果表明: (1)随着处理天数的增加, 高温加剧颖花退化, 与适温处理及CK相比, 高温处理9 d黄华占颖花退化率增加45.8%和62.9%, 丰两优6号颖花退化率增加81.7%和136.1%, 丰两优6号增加幅度大于黄华占。(2)高温处理1~3 d降低花粉活力, 5~9 d抑制花药充实。花药发育不良, 花粉形成受阻是导致结实率显著下降的原因, 两品种趋势一致。(3)高温处理缩短颖花长度, 降低籽粒充实性, 使千粒重显著下降, 热敏感性品种丰两优6号下降幅度大于耐热性品种黄华占, 籽粒充实在5 d以上高温处理下降显著。研究说明倒一叶生长时期高温胁迫影响穗生长发育, 生产中需加强相应栽培措施的调控。
Pot experiment was conducted to investigate the effects of heat stress on rice spikelets degeneration and grain filling at panicle initiation stage. A tolerant (Huanghuazhan) and a susceptible (Fengliangyou 6) cultivars were treated in phytotron with high temperature (10:00-15:00, 40℃) and normal temperature (10:00-15:00, 32℃) for one, three, five, seven, and nine days at last leaf emergence stage, with these grown under natural condition as control. The results indicated that the ratio of spikelet degeneration increased greatly with increasing heat-treating time. The ratio of spikelet degeneration of Huanghuazhan increased 45.8% and 62.9% after nine days under high temperature as compared with normal temperature treatment (control), and the ratio of Fengliangyou6 increased 81.7% and 136.1% in the same treatment, showing higher ratio of spikelet degeneration in Fengliangyou6 than Huanghuazhan. The pollen activity significantly decreased after one day under high temperature stress, and anther formation significantly inhibited after five days under high temperature stress. Bad anther formation and poor pollen activity under heat stress resulting in seed-setting rate significantly decreased. Spikelet length and grain filling of two cultivars decreased due to heat stress effect, resulting in a significant reduction of grain weight, with a larger reduction rate in Fengliangyou 6 than in Huanghuazhan. Grain filling reduced significantly after five days of high temperature treatment. According to the study, heat stress at last leaf emergence stage may sharply decrease rice yield, and corresponding cultivation measures should be strengthened in production.
20世纪70年代以来, 我国长江中下游稻区的水稻种植季节发生了重大转变, 双季稻面积大幅下降, 单季稻面积大幅上升。长江中下游稻区单季稻幼穗生长发育和开花结实集中在7月和8月份, 正值一年中气温最高时期[1, 2, 3]。全球气候变暖导致水稻生产季节高温胁迫天气(日最高温度高于35℃)频繁[4], 水稻开花期受高温胁迫天气影响最为严重, 胁迫导致花药开裂率降低, 散粉量下降, 柱头上花粉粒数量减少, 水稻受精结实受阻, 产量大幅下降[5, 6, 7, 8, 9]。因此, 目前普遍采用推迟水稻播种期来推迟开花时期, 避开高温与开花期相遇。然而, 开花期推迟后, 高温与穗分化期相遇。而且近年来高温胁迫天气的出现有提前的趋势, 2013年高温热害天气比往年提前了20多天, 且极端高温天气达40℃以上, 持续时间长达10~15 d[10]。播期的调整和高温天气的提前使长江中下游单季稻穗分化期受高温影响的机率增加。
2013年高温天气严重影响单季中稻的幼穗发育和开花结实, 颖花退化严重, 部分地区水稻绝收。有研究表明, 穗分化期高温会导致水稻花粉形成受阻[11], 代谢紊乱[12]。也有研究指出[13, 14], 高温会导致参与颖花形成、花粉发育的基因表达紊乱。目前对水稻穗分化期高温伤害的研究较少, 我们于2014年将穗分化期分成倒四叶、倒三叶、倒二叶、倒一叶4个生长时期, 以40℃高温处理7 d, 发现水稻倒一叶生长时期高温胁迫对穗部性状影响最为严重[15], 颖花退化幅度达50%以上, 结实率降低80%, 且千粒重也显著下降。倒一叶生长时期(花粉母细胞形成至减数分裂期)是水稻颖花形态生长、花器官发育最快的时期[16], 为进一步明确这一时期高温对穗发育的影响, 在2014年试验的基础上, 于2015年开展不同持续天数高温处理对水稻颖花退化和籽粒充实特性影响的研究, 以期为穗分化期高温伤害理论和穗分化期水稻品种耐热性评价提供参考依据。
选用耐热性籼稻品种黄华占(主茎总14片叶)和热敏感性籼稻品种丰两优6号(主茎总14片叶), 2个品种生育期基本一致。
2015年在中国水稻研究所试验场进行盆栽试验, 塑料种植盆规格为20 cm× 18 cm× 25 cm, 每盆装过筛土10 kg, 土壤pH 5.93、含有机质28.26 g kg-1、全氮1.50 g kg-1、碱解氮128.24 mg kg-1、全磷0.87 g kg-1、有效磷44.80 mg kg-1、全钾25.27 g kg-1、速效钾130 mg kg-1。于5月22日播种, 大田育秧, 6月17日选长势一致的秧苗移栽至塑料种植盆中, 每盆2穴, 每穴2苗, 共500盆。每盆施复合肥(N︰P2O5︰K2O=12︰24︰12) 4.00 g作基肥, 移栽后施尿素0.50 g作为分蘖肥, 穗分化开始时施复合肥2.25 g作为穗肥。水稻生长期间其他管理措施与高产栽培一致。
设置高温(HT)、适温(NT)及自然温度对照(CK) 3个处理。高温处理和适温处理均在人工气候箱中进行, 人工气候箱四周以超白钢化玻璃保持其光照与外界自然光照一致。根据盆栽场湿度测定, 设置人工气候箱湿度为75%。人工气候箱内部装有鼓风机, 保持CO2浓度与室外基本一致。自然温度处理(CK)在室外自然条件下进行。高温(HT)处理的温度为7:00— 10:00 36.5℃、10:00— 15:00 40℃、15:00— 17:00 36.5℃, 17:00— 次日7:00 33.0℃, 日平均温度为36.5℃。适温(NT)处理的温度为7:00— 10:00 28.5℃、10:00— 15:00 32℃、15:00— 17:00 28.5℃、17:00— 次日7:00 25℃, 日平均气温为28.5℃。自然温度对照(CK)日平均温度为28.4℃, 10:00— 15:00平均温度为32.9℃, 适温处理的温度设置与自然温度处理接近。
在水稻植株倒一叶心叶抽出时, 将待处理的盆栽水稻移入40℃及32℃人工气候箱, 分别处理1、3、5、7和9 d后移出到自然环境下生长, 每品种每处理20盆。处理期间保持盆栽土壤水层3~4 cm。黄华占处理时间为7月28日至8月5日, 丰两优6号处理时间为7月29日至8月6日。
1.4.1 生育进程记载 移栽后对所有主茎蘖挂牌, 每隔5 d记录叶龄, 以掌握水稻生长进程。
1.4.2 主茎穗颖花分化与退化及颖花大小 两品种不同胁迫时间处理均于主茎稻穗抽出约1/2时集中取样一次, 选取大小一致主茎穗。采用松岛省三痕迹法[17]考察每穗颖花数与退化数, 退化的颖花为具有白色的小颖花状薄膜痕迹, 部分可用肉眼观察计数, 部分颖花退化数痕迹较小在体视显微镜(OLYMPUS, SZX9)下观察计数, 记录逐个枝梗颖花分化与退化数。以8个穗为一个样本, 3次重复, 取平均值。每穗颖花分化数为每穗颖花数和每穗颖花退化数的总和。
颖花退化率(%) = (颖花退化数/颖花分化数)× 100
颖花大小利用万深考种仪(万深SC-G型)考察。
1.4.3 主茎穗花药形态大小 两品种不同胁迫时间处理于稻穗抽出约20%时集中取样一次, 选取大小一致主茎穗。从每穗上、中、下部位各取3个颖花在体视显微镜下(OLYMPUS, SZX9)观察花药形态, 照相。利用测微尺测量花药长度和宽度。3个穗为一个样本, 3次重复, 取平均值。
1.4.4 主茎穗花粉活力 取样方法时期同1.4.3。从每穗上、中、下部位各取3个颖花, 用1% I2-KI溶液染色并于生物显微镜下(OLYMPUS, BX61)选取3个视野统计可染率, 染色花粉即具有活力的花粉。3个穗为一个样本, 3次重复, 取平均值。
花粉活力(%) = (染色花粉粒数/花粉粒总数) × 100
1.4.5 主茎穗部性状 水稻成熟后, 选取每处理大小一致主茎穗, 测定每穗粒数、结实率及单穗重量, 利用万深考种仪(万深SC-G型)分别考察稻谷面积和稻谷粒重, 并计算稻谷比粒重(单位面积稻谷重)。每8个穗为一个样本, 3次重复, 取平均值。
稻谷比粒重(GRWA, mg mm-2) = 稻谷重(g) / 稻谷面积(mm2) × 1000
采用Microsoft Excel统计数据, 用SAS 9.2软件进行方差分析, Duncan’ s法(新复极差法, SSR)检验处理间差异。
倒一叶生长期水稻颖花分化已经完成, 高温主要对水稻颖花退化及花器官发育造成影响, 结果见表1。高温导致两品种每穗粒数显著下降, 并随胁迫时间延长下降幅度增大。高温处理9 d后黄华占每穗粒数比CK及适温处理下降31.6%和23.5%, 丰两优6号同比下降55.7%和47.6%。每穗粒数的下降主要由于颖花退化率的增加, 高温处理使黄华占和丰两优6号颖花退化率每天平均增加2.5%和3.6%, 其中高温5~7 d颖花退化率增加较快, 高温处理9 d黄华占和丰两优6号颖花退化率分别比CK增加55.6%和136.1%, 比适温处理增加45.7%和81.7%。从不同一次枝梗部位上的颖花退化分析可知(图1), 两品种均表现为颖花退化增加幅度下部要大于上部, 品种中部一次枝梗上的颖花退化表现出明显差异, 且热敏感性品种丰两优6号中部颖花退化率要大于耐热性品种黄华占。从结实率上看, 随着高温处理时间的延长呈显著下降趋势, 高温处理9 d黄华占比CK降低95.0%, 丰两优6号比CK降低97.1%, 热敏感性品种下降速度更快。适温处理与CK相比处理1~5 d颖花退化率无显著差异, 适温处理7~9 d颖花退化率较CK有所增加, 而适温处理与CK相比结实率无显著差异。说明倒一叶高温处理对颖花退化和结实率的影响是一个动态过程, 随着高温处理天数的增加, 颖花退化不断加剧, 耐热性品种受影响程度小于热敏感性品种。
| 表1 高温处理对水稻颖花退化和结实率的影响 Table 1 Effect of high temperature on rice spikelets degeneration and seed-setting rate |
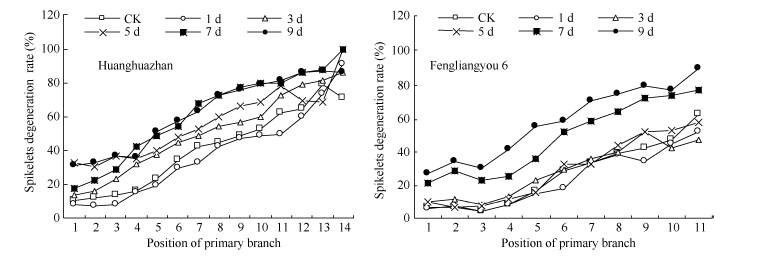 | 图1 高温处理对不同部位枝梗上的退化率的影响 对照为自然环境条件下的处理; 图中稻穗一次枝梗部位顺序依次从稻穗顶部到基部。Fig. 1 Effect of high temperature on degenerated spikelet ratio of different primary branches CK is the treatment under natural condition; Order of primary branches is from panicle top to bottom. |
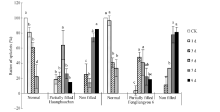
2.2.1 花药充实性 高温胁迫导致花药充实性变差(图2), 高温处理后花药多出现不充实或半充实现象, 为便于分析, 将颖花分成花药充实、花药部分充实和花药不充实3个级别。高温处理1 d, 花药受影响较小, 随着高温处理时间的增加, 花药部分充实和不充实的颖花所占比例逐渐增加, 高温处理5 d花药部分充实的颖花占较高比例, 而高温处理7 d和9 d后花药多出现不充实的透明状态, 无花粉形成, 高温处理9 d表现尤为明显(图3)。
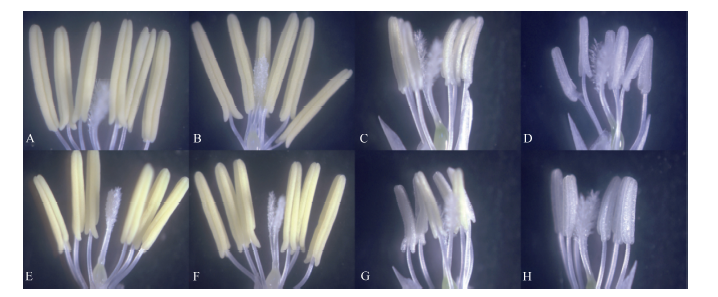 | 图2 高温处理花药性状的分类 A~D: 黄华占品种的花药; E~H: 丰两优6号的花药; A, E: 对照; B, F: 适温处理; C~D, G~H: 高温处理; A、B和E、F是正常的花药; C和G是部分充实花药; D和H是不充实的花药。Fig. 2 Classification of anther characters under high temperature A-D: Huanghuazhan’ s anther; E-H: Fengliangyou 6’ s anther; A and E: CK; B and F: normal temperature treatment; C-D and G-H: high temperature treatment; A-B and E-F: normal anther; C and G: partially filled anther; D and H: non-filled anther. |
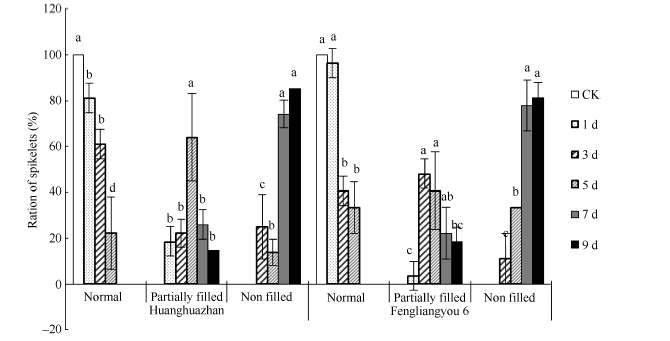 | 图3 高温处理后不同类型花药颖花比例 对照(CK)为自然环境条件下的处理; 不同小写字母表示处理间差异显著(P< 0.05)。Fig. 3 Ratio of spikelets with different anther characters under high temperature CK is the treatment under natural condition. Bars superscripted by different small letters are significantly different at the 0.05 probability level among treatments. |
2.2.2 花药大小 高温胁迫导致花药充实受阻, 花药形态变小, 结果如表2所示, 高温处理3 d对黄华占花药大小影响较小, 而使丰两优6号花药长度和宽度比CK降低12.4%和10.5%。高温处理5 d后黄华占和丰两优6号花药长度和宽度下降幅度增加, 高温处理9 d黄华占花药长度和宽度分别比CK降低了38.2%和30.2%, 丰两优6号花药长度和宽度比CK下降了24.8%和18.4%, 黄华占下降幅度要大于丰两优6号。
| 表2 高温处理对花药大小的影响 Table 2 Effect of high temperature on the size of anther (mm) |
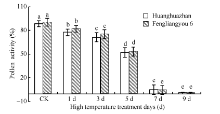
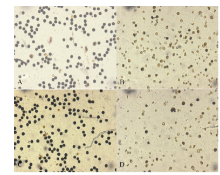
2.2.3 花粉活力 如图4所示, 随着高温处理天数花粉活力增加逐渐下降, 随着高温处理时间的增加, 花药充实受阻, 不充实的花药无花粉粒形成, 且充实性差的花药里花粉多为畸形(图5), 2个品种变化趋势一致。
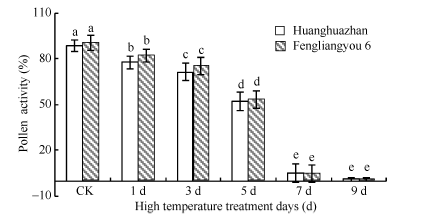 | 图4 高温处理对花粉活力的影响 对照为自然环境条件下的处理; 不同小写字母表示处理间差异显著(P< 0.05)。Fig. 4 Effect of high temperature on pollen viability CK is the treatment under natural condition; Bars superscripted by different small letters are significantly different at the 0.05 probability level among treatments. |
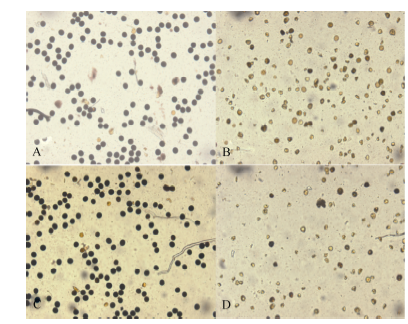 | 图5 不同花药的花粉性状比较 A: 黄华占正常花药的花粉; B: 黄华占充实性差花药的花粉; C: 丰两优6号正常花药的花粉; D: 丰两优6号充实性差花药的花粉。Fig. 5 Comparison of pollen character in different anthers A: pollen of normal anther in Huanghuazhan; B: pollen of partially filled anther in Huanghuazhan; C: pollen of normal anther in Fengliangyou 6; D: pollen of partially filled anther in Fengliangyou 6. |
2.3.1 颖花大小 颖花大小是决定千粒重的重要因子, 由表3可知, 颖花长度受高温影响十分敏感, 高温处理1 d可导致颖花长度显著下降, 且随着高温处理时间的延长, 颖花长度逐渐缩短, 高温处理9 d黄华占颖花长度比CK降低10.3%, 丰两优6号颖花长度比CK降低24.4%, 适温处理与CK相比有下降的趋势, 但下降幅度远远小于高温处理, 高温对丰两优6号颖花长度的影响大于对黄华占。颖花宽度受高温处理后有增加趋势, 但相对于颖花长度的变化高温对颖花宽度的影响要小得多。
| 表3 高温处理对颖花大小的影响 Table 3 Effect of high temperature on size of spikelets (mm) |
2.3.2 千粒重及比粒重 由表4可知, 高温处理导致黄华占和丰两优6号千粒重平均比CK处理降低11.6%及23.1%, 比适温处理降低13.2%和23.0%, 且下降幅度随着高温处理天数的增加显著增加, 高温处理9 d黄华占千粒重比CK及适温处理分别降低16.5%和18.6%, 丰两优6号同比下降42.9%和43.1%。颖花大小和籽粒充实性是影响千粒重的2个主要因素, 随着高温处理天数的增加, 水稻比粒重呈下降趋势, 品种间变化存在差异, 高温处理1 d和3 d比粒重受高温影响较小, 高温处理5~9 d比粒重显著降低, 高温处理9 d黄华占比粒重比CK降低25.0%, 丰两优6号比粒重比CK降低18.4%。CK处理与适温处理相比差异不明显。
| 表4 高温处理对比粒重和千粒重的影响 Table 4 Effect of high temperature on ratio of grain weight to area and 1000-grain weight |
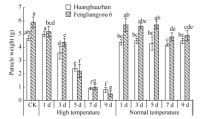
高温导致单穗重显著下降(图6), 高温处理黄华占单穗重平均值分别比CK和适温处理低46.0%和42.0%, 丰两优6号则降低51.9%和50.2%。随着高温处理时间的增加, 2个品种单穗重逐渐下降, 高温处理3 d单穗重开始显著下降, 日均降幅达5.1% (黄华占)和6.1% (丰两优6号), 高温处理9 d黄华占和丰两优6号的单穗重只有CK的17.0%和8.5%。由此说明, 随着高温持续时间的增加, 耐热性品种所受影响要小于热敏感性品种。相关分析表明(表5), 穗部性状和高温处理天数呈极显著负相关, 说明倒一叶生长期高温处理持续时间的增加会加剧对水稻穗发育的伤害。
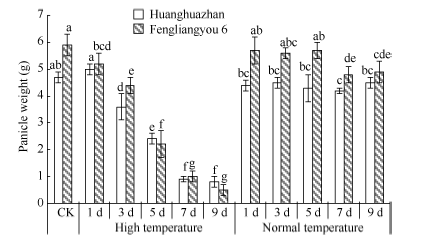 | 图6 高温处理对单穗重的影响 对照(CK)为自然环境条件下的处理; 不同小写字母表示处理间差异显著(P< 0.05)。Fig. 6 Effect of high temperature on rice panicle weight CK is the treatment under natural condition; Bars superscripted by different small letters are significantly difference at the 0.05 probability level among treatments. |
| 表5 高温处理穗部性状的相关性分析 Table 5 Correlation coefficients of panicle traits under high temperature |
方差分析(表6)表明, 穗粒数在品种间变异总差异并不明显, 但在处理天数、品种与处理天数交互间变异差异达极显著水平。结实率、千粒重及穗重在品种间、处理天数、品种和处理天数交互的变异均达到显著或极显著水平。说明高温处理品种间穗粒数变化幅度相一致, 品种间的穗粒数差异随着处理天数的增加逐渐表现出来, 2个品种间的差异在结实率和千粒重上表现最为显著。
| 表6 穗部性状总变异方差分析F值 Table 6 F-values in ANOVA analysis of panicle traits in different treatments |
目前有关高温导致颖花退化的研究较少, 本试验结果表明水稻倒一叶生长时期(花粉母细胞形成至减数分裂期)高温时间延长加剧颖花退化, 导致白穗。正常情况下, 水稻颖花退化主要集中在稻穗下部, 引起颖花退化的首要原因是下部颖花物质供应不足[18, 19], 导致细胞凋亡[20], 本试验也发现颖花退化与穗部干物质积累减小有关。随着高温处理时间的延长, 稻穗中上部颖花退化率也明显增加, 颖花生长发育受到抑制, 未退化颖花变小, 除了物质供应的原因外, 可能还与内源激素比例失调[21, 22], 活性氧含量增加[8], 颖花细胞遭到破坏有关。
高温下结实率下降的原因是花器官受到伤害, 水稻倒一叶生长时期是花器官生长发育的重要时期, 这一时期高温处理对花药的影响是一个动态的过程, 高温处理时间影响花药充实和花粉发育具有累积效应。邓运等[11]研究指出, 花器官发育早期高温处理使生殖细胞排列紊乱, 导致花器官发育不良, Prasa等[23]和Mohamme等[24]指出高温会降低花粉量, 本研究与之结果类似, 本试验从倒一叶心叶长出时开始, 高温处理时期早, 覆盖了花药的发育前期, 高温处理5~9 d, 花药发育异常, 不充实花药无花粉粒形成, 部分充实的花药内花粉多为畸形。开花期高温也是通过影响花药开裂和柱头上花粉量导致结实率下降[5, 6, 7, 8, 9], 曹云英[12]等发现减数分裂期高温处理也降低了花药开裂能力, Matsui等[25]研究表明花药开裂受阻的原因是高温降低花粉粒的膨胀能力, 导致花粉囊鞘开裂不良, 本试验高温处理后花粉形成受到了影响, 生理特性变差势必影响花粉吸水膨胀, 进而影响花药开裂, 花药中花粉数量的减少也影响柱头上的花粉量, 这是本试验结实率大幅下降的原因。雌雄蕊发育是同时进行的, 形态观察表明, 高温可能对柱头分枝数有影响。高温对花器官生长发育及裂药特性影响的机制还需进一步研究。
千粒重取决于颖花大小和籽粒饱满度, 后者跟干物质供应有关[26, 27]。前人研究表明[28]比粒重能够很好衡量籽粒饱满度。高温处理1~3 d, 两品种比粒重受高温影响均不大, 虽然结实率下降导致源库比上升, 但颖花大小决定了库容量, 千粒重主要受颖花大小的影响。郑建初等[29]研究表明, 花期高温处理使水稻千粒重上升, 本试验与之差异在于处理时期不一致, 花期高温处理并不会对颖花形成造成影响。倒一叶5~9 d的高温处理下, 热敏感性品种颖花大小和比粒重下降幅度均大于耐热性品种, 其结实率下降幅度也大于耐热性品种, 这跟物质运转发生改变有关。笔者在这个时期高温处理10 d的结果表明[30], 抽穗期黄华占穗干重占主茎干重的8.5%, 而丰两优6号只占6.0%, 按照源库理论, 丰两优6号的籽粒充实度应该大于黄华占, 但高温胁迫后, 高节间分蘖出现, 成为水稻新的营养生长中心, 主茎抽穗期丰两优6号高节间分蘖干重占到整个主茎的22.2%, 而黄华占只占20.2%, 丰两优6号高节间分蘖生长要快于黄华占, 这些分蘖到后期能够抽穗, 试验观察到高温抑制水稻主茎穗下颈节生长, 降低水稻抽穗程度, 这也说明水稻主茎物质下降, 物质运输向高节间分蘖偏移。上海植物生理研究所人工气候室研究指出[31], 花期高温处理也引起高节间分枝, 使千粒重下降, 推测高节间分蘖的出现导致籽粒灌浆倒退。高节间分蘖的生长机理可能跟再生稻的生长机理一致[32], 如果生产上高温导致大面积绝产, 经过适宜的调控使高节间分蘖抽穗结实, 可以弥补一些产量损失。
水稻倒一叶生长时期持续5 d以上高温天气对颖花发育和花器官形成的影响极其严重, 7~9 d甚至导致绝产, 生产上要密切关注水稻倒一叶生长时期的环境条件, 结合高温预警及时加强田间管理。如遇高温, 及时深水灌溉, 降低田间温度, 可以减少产量损失[33]。水杨酸的喷施对减小水稻颖花和花器官的伤害也有积极的作用[34]。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|