

{kind=link}
{kind=link}
{kind=link}
{kind=link}
花生栽培种与野生种( Arachis oteroi)人工杂交双二倍体的创制和鉴定
[李丽娜1, 2, *  , 杜培
, 杜培2, * , 付留洋2, 3 , 刘华2 , 徐静2 , 秦利2 , 严玫2 , 韩锁义2 , 黄冰艳2 , 董文召2 , 汤丰收2 , 张新友2, * ]
, 杜培]
|
|


第一作者联系方式: E-mail:lilina198283@163.com, Tel: 0371-65718247
花生野生种是改良花生栽培种的重要基因资源。为了利用野生花生的抗性基因, 本研究利用花生栽培品种豫花9331与二倍体野生种 A. oteroi人工杂交, 借助胚拯救和染色体秋水仙素加倍, 创制一个双二倍体杂种AmE-4, 并利用荧光原位杂交和分子标记技术准确鉴定了该双二倍体。观察结果表明, AmE-4的叶片与豫花9331存在显著差异, 而主茎高、侧枝长和总分枝数等性状与豫花9331差异不显著。AmE-4开花期较豫花9331推迟60 d, 结实性与荚果发育状况较差, 不利于AmE-4的育种利用。同时, 开发了57个追踪AmE-4中 A. oteroi染色体的显性或共显性SSR标记, 为创制和鉴定花生栽培种 A. oteroi易位系或渐渗系奠定分子基础。
, DU Pei
Wild Arachis species are important genetic resources. To introgress resistant genes of Arachis species, we developed a new amphidiploid AmE-4 through man-made cross between cultivated peanut variety Yuhua 9331 and a diploid Arachis species A. oteroi, with the assistance of following embryo rescue and chromosome doubling by colchicine treatment. AmE-4 was identified and characterized by fluorescence in situ hybridization (FISH) and SSR molecular marker. Morphological observation revealed significant differences in leaves between amphidiploid AmE-4 and Yuhua 9331, while the agronomic traits such as main stem height, length of first lateral branch and number of branches showed less difference between them. The date of first flower appearance in AmE-4 delayed sixty days compared with that in Yuhua 9331, and its pods setting and development were also poor, which would hinder its further utilization. In addition, 57 dominant or co-dominant SSR molecular markers were developed and could be used to identify translocation or introgression lines with A. oteroi chromosome fragment in future studies.
栽培种花生(Arachis hypogaea L., 2n = 4x = AABB)是世界主要的油料、经济作物, 也是人类所需的重要植物蛋白来源。然而长期的栽培和人工驯化导致花生栽培品种遗传多样性降低, 严重影响花生的育种效率。花生野生种具有栽培种不具备的优良性状。国际半干旱热带地区作物研究所(ICRISAT)、美国、中国等的研究表明, 野生花生中存在丰富的叶斑病、锈病、线虫病、病毒病等抗源和高含油量等优质性状, 而且对叶斑病和锈病的抗性机制与栽培种花生有所不同, 是改良花生栽培种的重要基因资源[1, 2, 3, 4]。
花生栽培种和多数野生种杂交成功率很低, 较难用于育种。栽培种除与花生组(Section Arachis)内野生种杂交表现出一定的亲和性外, 由于激素代谢的失衡导致种间杂种胚胎的败育, 与其他近缘野生种杂交大多表现不亲和、弱亲和或杂种胚败育(夭亡)[5, 6]。目前克服这些问题和利用野生花生种有: 野生同源四倍体途径(二倍体野生种经人工染色体加倍形成)[7]、野生异源四倍体途径(二倍体野生种之间杂交并经染色体加倍形成的双二倍体)[8]和三倍体-六倍体途径(栽培种与二倍体野生种杂交并经染色体加倍形成异源六倍体)等[9]。其中三倍体-六倍体途径是应用最多的方法, 该方法通过栽培种与野生二倍体种杂交与必要的胚拯救等技术, 获得育性低甚至完全不育的种间杂种F1(2n = 3x)[10], 经过自然加倍或秋水仙碱处理加倍, 获得六倍体花生(2n = 6x, 即双二倍体), 直接将野生种的整套染色质导入到双二倍体中。双二倍体花生染色体遗传相对稳定, 更容易与栽培亲本回交获得后代, 并通过染色体交换将外源染色体易位或渐渗到栽培种中。目前, 利用这种途径创制的聚合了优良农艺性状的栽、野种间杂交衍生系已被报道[11, 12], 如利用栽培品种白沙1016与二倍体野生种A. diogoi杂交育成的远杂9102, 具有高抗青枯病、耐逆、高固氮效率、适应性广等突出特点, 是目前我国珍珠豆型花生的主栽品种[13]。此外, 栽培种与A. glabrata的衍生系对花生芽坏死表现高抗性[14], 与A. kretschmeri衍生系抗网斑病[15], 与A. cardenasii衍生系抗根结线虫病和晚斑病等[16, 17], 显示了野生花生基因资源在栽培种改良中的巨大利用潜力。
A. oteroi是河南省农业科学院经济作物研究所从中国农业科学院油料作物研究所引进的野生花生种质, 具有含油量高、中抗根结线虫病等优良特征, 是改良花生的重要基因资源[4, 18]。本研究拟通过花生品种豫花9331与野生种A. oteroi人工杂交获得双二倍体杂种, 对该双二倍体表型分析和分子标记筛选, 旨在创制育种的中间材料, 以开拓野生种在育种中应用。
以栽培种品种豫花9331为母本(2n = 4x = 40, AABB)、二倍体野生种A. oteroi为父本(2n = 2x = 20)进行种间杂交。其中豫花9331为河南省农业科学院经济作物研究所选育保存, A. oteroi由中国农业科学院油料作物研究所引进。SSR分子标记引物由华大基因(深圳, 中国)合成, 序列分别来自He等[19]、Moretzsohn等[20]、Palmieri等[21]、Ferguson等[22]和山东省农业科学院王兴军研究员。
杂交技术同常规有性杂交, 于生育后期从母本植株上收获杂交荚果, 在无菌条件下用5%的次氯酸钠对荚果表面消毒, 然后打开荚果果壳, 将幼胚接种到由MS+0.075 mg L-1 IAA (3-吲哚乙酸)+0.01 mg L-1 Kn (激动素)+5%蔗糖组成的固体培养基上, 于25℃、18 h/6 h光暗交替条件下培养, 幼胚发育成苗后将幼苗转移至由MS、0.8 mg L-1 NAA和3%蔗糖组成的固体生根培养基上培养, 直至形成根茎叶完整的种间杂种F1植株。
在无菌条件下将F1组培苗从顶端数第3片叶下切开, 取顶芽部分, 将其置MS+0.1 mg L-1 NAA+0.4 mg L-1 6-BA+3%蔗糖+0.05%秋水仙素固体培养基上, 在25℃恒温光照培养间中诱导芽分化。约14 d后, 把苗转移到MS+0.8 mg L-1 NAA+3%蔗糖固体生根培养基, 培养15~30 d, 直至形成完整植株S0, 然后移栽于生长池中。S0成株后部分枝条能够产生果针, 观察统计S0有果针枝条(双二倍体枝条)与无果针枝条(三倍体枝条)花粉育性。具体方法是S0开花盛期取每种枝条两朵花, 用1%醋酸洋红染色, 在显微镜下观察, 统计每朵花5个视野的可育与不育花粉粒数, 花粉活力(%) = 可育花粉数/(可育花粉数+不育花粉数) × 100%[23]。待S0荚果成熟后, 取荚果, 用5%的次氯酸钠消毒, 再将种子接到1/2 MS+5%蔗糖固体培养基上, 置于25℃恒温光照培养间培养, 直至发育成幼苗S1(AmE-4)。经过组织培养扩繁后, 次年移栽于花盆或生长池中, 并同时种植豫花9331。生育期内对豫花9331和AmE-4各4株进行表型性状调查, 始花期记录第一朵花开花日期, 成熟期调查株型、总分枝数、叶柄长、主茎第四片叶上部小叶长和宽、主茎高和侧枝长等性状。
参照杜培等[24]根尖细胞中期染色体制片的方法。基因组荧光原位杂交是采用CTAB法提取A. oteroi全基因组DNA, 以缺刻平移法将绿色荧光素(Fluorescein-12-dUTP)标记到A. oteroi全基因组DNA上, 以栽培品种豫花9331基因组DNA为封阻(封阻与探针浓度比例约为1︰50)。杂交液含去离子甲酰胺7.5 µ L, 20× SSC (0.015 mol L-1 C6H5Na3O7· 2H2O-0.15 mol L-1 NaCl) 1.5 µ L, 探针2.5 µ L, 封阻1 µ L, 鲑鱼精DNA 0.5 µ L, 50%硫酸葡聚糖2 µ L, 103℃变性13 min, 立即将杂交液离心管置-20℃的100%无水乙醇浴10 min; 染色体制片以70%甲酰胺78℃变性70 s后立即置-20℃的70%、95%和100%无水乙醇中梯度脱水各5 min, 气干。滴加杂交液, 盖上盖玻片, 37℃杂交至少6 h。采用42℃水浴在2× SSC中洗片10 min, 50%甲酰胺洗5 min, 再2× SSC 10 min洗2次, 然后用DAPI染色, LeicaDM6000荧光显微镜观察照相。5S rDNA和45S rDNA原位杂交杂交程序同上, 不同的是5S rDNA和45S rDNA分别用Biotin-16-dUTP和Digoxigenin-11-dUTP标记, 再分别用Anti-Biotin-FITC和Anti-Digoxigenin- Rhodamine检测。
采用已获得的豫花9331和A. oteroi共显性SSR标记E420 (F: 5′ -TCCATCGTTAGTGGCACTGT-3′ ; R: GTCGA CTCCTGCCCAATCTA-3′ )和B051 (F: 5′ -TACAGCATTG CCTTCTGGTG-3′ ; R: 5′ -CCTGGGCTGGGGTATTATTT-3′ )鉴定AmE-4, 再利用840对SSR标记对豫花9331、A. oteroi和AmE-4进行PCR扩增和标记筛选, 用于特异追踪外源染色体。PCR体系为10 µ L, 包括模板基因组DNA 0.9 µ L (25 ng mL-1), 10 µ mol L-1SSR上下游引物各0.2 µ L, 10× 缓冲液1 µ L, dNTPs (2.5 mmol L-1) 0.9 µ L, MgCl2(25 mmol L-1) 0.8 µ L, Taqase 0.1 µ L (0.5 U), ddH2O 5.9 µ L。PCR扩增程序为94℃预变性3 min; 94℃变性30 s; 55℃退火45 s; 72℃延伸1.2 min; 33个循环后72℃延伸10 min; 反应结束以后聚丙烯酰胺凝胶电泳检测。
豫花9331与A. oteroi人工杂交后, 获得17个荚果、20枚胚珠, 将胚珠解剖后进行胚拯救, 待F1成苗后观察根尖细胞中期染色体, 最终获得12株(E-1~E-12)染色体数为30条的种间杂种F1植株(2n= 3x= 30)。然后对E-1~E-5试管苗进行染色体加倍处理, 组培苗出现明显的中毒症状(如叶子卷曲, 部分叶黄萎等), 约14 d后转移至生根培养基, 最终获得植株S0。
对F1和S0花粉育性观察结果显示, 杂种F1和S0上无果针枝条的花粉活力分别为1.03%和1.48%, 均表现高度不育, 而S0上有果针枝条花粉活力为62.67%, 花粉活力大幅度提高(表1, 图1-A、1-B)。生育后期收获时, 发现5株S0中有1株(E-4)结实, 占20%。E-4荚果经1/2MS+5%蔗糖固体培养基培养后获得幼苗S1(AmE-4)。
| 表1 豫花9331与A. oteroi杂种F1和S0的花粉活力 Table 1 Pollen viability of S0and interspecific hybrid F1 of Yuhua 9331 and A. oteroi |
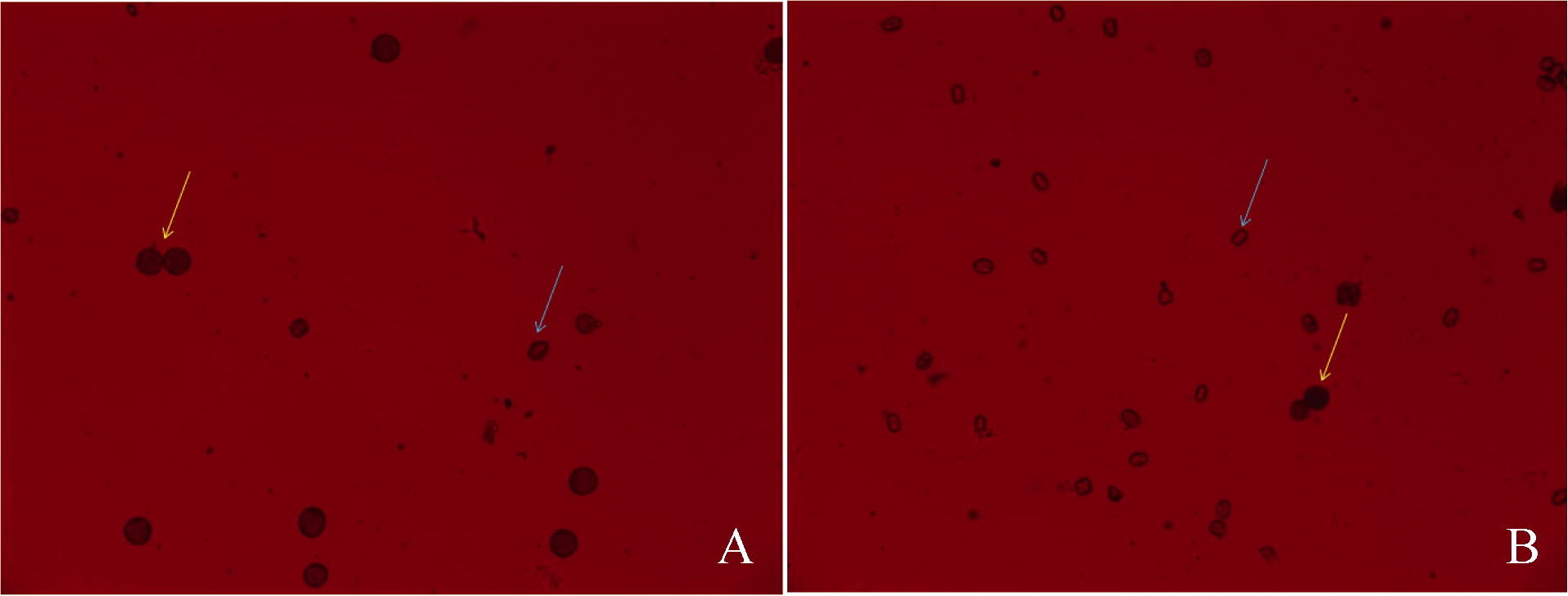 | 图1 S0有果针枝(A)和无果针枝(B)花的花粉染色 黄色箭头所示为可育花粉粒, 蓝色箭头所示为不育花粉粒。Fig. 1 Pollen staining from flowers in branches with (A) and without pegs (B) Yellow arrows show fertile pollens and blue arrows show sterile pollens. |
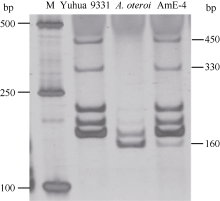
利用已获得的豫花9331和A. oteroi共显性SSR标记E420对豫花9331、A. oteroi和AmE-4进行分析, 发现在豫花9331上约450 bp和330 bp处有特异扩增条带, 父本A. oteroi在约160 bp处有特异条带, 而AmE-4在450、330和160 bp处均有特异条带(图2), 说明异源六倍体花生AmE-4同时包含了豫花9331和A. oteroi的遗传物质, 用SSR标记B051也显示了同样的结果。
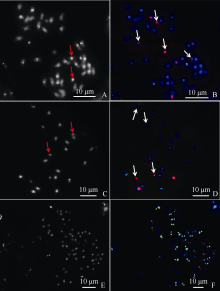
为了识别大随体染色体, 准确鉴定豫花9331和A. oteroi染色体数目, 以5S rDNA和45S rDNA为探针对豫花9331和A. oteroi进行FISH分析和DAPI染色, 发现豫花9331与A. oteroi各有1对“ 大随体染色体” (图3-B、3-D), 染色体数分别是40和20 (图3-A、3-C), 而AmE-4
染色体数为60条, 是豫花9331与A. oteroi全部染色体组数目总和(图3-E)。A. oteroi染色体组与栽培花生的A染色体组类似, 都存在明显的着丝粒DAPI带和1对小染色体(图3-A、3-C), 显示A染色体组特征[25]。进一步利用A. oteroi基因组荧光原位杂交分析, 发现AmE-4有20条染色体显示出明亮的杂交信号, 12条显示弱的杂交信号(图3-F), 并且有杂交信号的染色体均有明亮的DAPI带, 显示A染色体组特征, 这说明A. oteroi基因组探针除了与AmE-4中A. oteroi的全部染色体杂交外, 也与豫花9331的部分A染色体组产生了交叉信号。
 | 图2 SSR标记E420在豫花9331、A. oteroi和AmE-4中的扩增结果Fig. 2 PCR amplification pattern of SSR E420 in Yuhua 9331, A. oteroi and AmE-4 (M: marker) |
 | 图3 豫花9331、A. oteroi和AmE-4荧光原位杂交 A~B: 豫花9331 DAPI染色及45S rDNA (红)和5S rDNA (绿)探针荧光原位杂交; C~D: A. oteroi DAPI染色及45S rDNA (红)和5S rDNA (绿)探针荧光原位杂交; E~F: AmE-4 DAPI染色和A.oteroi全基因组探针原位杂交。红色箭头所示为小染色体, 白色箭头所示为“ 大随体染色体” ; F图中的白色* 表示强基因组杂交信号的染色体, F图中的红色+表示弱基因组杂交信号的染色体。Fig. 3 Fluorescence in situ hybridization of Yuhua 9331, A. oteroi and AmE-4 A-B: DAPI staining and FISH of Yuhua9331 with 45S rDNA (red) and 5S rDNA (green) probes; C-D: DAPI staining and FISH of A. oteroi with 45S rDNA (red) and 5S rDNA (green) probes; E-F: DAPI staining and GISH of AmE-4 with A. oteroi total genomic DNA probes. Red arrows show the small chromosomes, white arrows show “ the big SAT-chromosomes” ; * show chromosomes with strong signals of A. oteroi total genomic DNA probes, and + show chromosomes with weak signals of A. oteroi total genomic DNA probes in Fig. F. |
利用840对SSR引物对豫花9331、A. oteroi和双二倍体AmE-4分析, 共获得57个标记能特异追踪双二倍体中A. oteroi的染色体(片段), 占6.8%, 其中共显性标记46个, 显性标记11个(表2)。
| 表2 双二倍体AmE-4中A. oteroi特异扩增分子标记 Table 2 Dominant or co-dominant molecular markers of A. oteroi in AmE-4 |
为了进一步了解豫花9331-A. oteroi双二倍体AmE-4的遗传特性, 本研究对盆栽豫花9331和双二倍体AmE-4的始花期、株型、生育后期主茎叶叶柄长、叶长、叶宽、主茎高、侧枝长和总分枝数进行调查分析, 结果表明, 豫花9331为直立型, A. oteroi为蔓生型, AmE-4为半直立型(图4-A), AmE-4较豫花9331始花期约延迟60 d, 开花后AmE-4能正常下针, 但结实性差, 荚果发育不良, 饱满度差。AmE-4叶片浓绿, 叶片明显加厚且向背面稍翻, 叶柄长、主茎叶长和叶宽与豫花9331差异显著(图4-B), 而在主茎高、侧枝长和总分枝数上与豫花9331差异不显著(表3)。
 | 图4 双二倍体AmE-4、豫花9331和A. oteroi的株型(A)和叶片形态(B)Fig. 4 Morphologies of plant (A) and leaves (B) of AmE-4, Yuhua 9331, and A. oteroi. |
| 表3 双二倍体AmE-4与豫花9331的农艺性状比较 Table 3 Comparison of amphidiploid AmE-4 withYuhua 9331 in certain agronomic traits |
以往花生种间杂种染色体研究多采用常规染色的方法[14, 30], 然而Seijo等[31]认为花生栽培种和野生种中包含一对或几对“ 大随体染色体” , 通常情况下这对染色体次缢痕位置容易被拉伸, 使随体和染色体臂离得很远。因此这种方法不能有效识别“ 大随体染色体” , 也不能区分杂种后代中栽培种与野生种的染色体组。本研究首先利用5S rDNA和45S rDNA探针识别“ 大随体染色体” , 确定栽培种和野生种染色体数, 进而准确计数杂种后代染色体。又利用基因组原位杂交和DAPI染色识别AmE-4中包含的A. oteroi染色体组。结果发现A. oteroi染色体组与栽培种A染色体组类似, 都具有明亮的染色体DAPI着丝粒带和小染色体, 而不同于B染色体组的没有或弱的DAPI带[32]。当以A. oteroi全基因组DNA为探针与AmE-4染色体杂交时, 来自栽培种的A染色体组也显示了弱的杂交信号, 表明A. oteroi染色体组与栽培种的A染色体组有较近的亲缘关系, 至少明显比B染色体组近。根据分类, A. oteroi为直立区组(Er)野生种[33], 其染色组不同于花生区组A、B染色组, 本研究的结论显然与以往的报道不一致, 这可能是所用A. oteroi的来源不同所致; 也可能由于以往的研究主要基于染色体形态的观察, 并没有反应出2个染色体组在分子水平上相似程度, 因而不够可靠。
二倍体野生种A. oteroi除了具有高含油量和抗根结线虫病等性状外[4, 18], 我们发现它还具有较强的叶斑病和锈病抗性。然而由于A. oteroi在河南原阳生态条件下不结实, 缺少足够的实生苗进行系统的抗病性研究, 双二倍体AmE-4的合成为我们下一步系统研究A. oteroi抗病等有利基因在栽培花生遗传背景下表现提供了材料基础。性状观察结果表明, AmE-4叶部性状与豫花9331有明显差异, 其主茎高、侧枝长和总分枝数等农艺性状与豫花9331差异不明显, 株型与母本豫花9331更为接近; 而AmE-4开花期严重推迟, 结实性差, 荚果发育不良, 可能是由于河南9月后光照温度不能满足其生长, 也可能是由于A. oteroi与A染色体组亲缘关系较近, 染色体加倍后容易形成复杂的多价体联会进而影响染色体的稳定性, 这些均不利于AmE-4的利用。为了能够有效利用AmE-4中有利基因, 需要打破不良连锁, 发展生产上具有利用价值的外源染色体补偿性易位或渐渗系[34]。目前我们正进一步利用AmE-4与豫花9331杂交创制易位系或渐渗系。而分子标记是追踪易位或渐渗片段的重要手段, 本研究开发的57个追踪AmE-4中A. oteroi染色体的显性或共显性标记, 将在今后A. oteroi易位系或渐渗系鉴定过程中发挥重要作用。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|