



{kind=link}
{kind=link}
{kind=link}
{kind=link}
外源ABA对低温胁迫下玉米幼苗内源激素含量及 Asr1基因表达的调节
[李馨园1, 2, *  , 杨晔
, 杨晔1, * , 张丽芳1 , 左师宇1 , 李丽杰1 , 焦健1 , 李晶1*, * ]
, 杨晔, 张丽芳]
|
|


第一作者联系方式: 李馨园, E-mail:lipan.08@163.com; 杨晔, E-mail:125915501@qq.com
脱落酸(ABA)是低温逆境下的重要信号因子, 为了探讨外源ABA对低温胁迫下玉米幼苗的生长调节作用, 以耐低温玉米品种久龙5号为试验材料, 采用不同浓度(5、15、25、35 mg L-1) ABA于玉米三叶一心时喷雾于叶片, 并进行低温梯次处理。分析处理后玉米叶片相对电导率、抗氧化酶活性及内源激素ABA、IAA的含量变化, 并采用Real-time PCR明确 Asr1基因表达水平变化。结果表明, 低温胁迫下不同浓度外源ABA处理的玉米叶片相对电导率整体呈上升趋势, SOD和POD活性加强, 15 mg L-1和25 mg L-1ABA处理的SOD活性均显著高于未应用ABA处理, 玉米內源ABA和IAA合成水平上升, 应用ABA后 Asr1基因相对表达水平上调, 其中5、15和25 mg L-1浓度处理基因表达上调显著。相关分析表明, ABA含量与 Asrl基因相对表达量、SOD活性均表现为极显著正相关, 与POD活性显著正相关。说明 Asr1基因表达受ABA的介导调控, Asr1基因表达量的提升, 也促进了内源ABA的合成, 抗氧化酶活性加强, 提升了应用ABA后玉米的抗低温能力。但外源ABA的介导调控具有一定浓度效应, 表现为低促高抑。
Abscisic acid (ABA) is an important signal factor under low-temperature stress. This study aimed to investigate the effect of exogenous ABA on growth regulation of maize seedlings under low-temperature stress. Maize variety “Jiulong 5” resistant to low temperature was used with foliage spray of ABA (5, 15, 25, 35 mg L-1) at three-leaf period under varying degrees of low-temperature, to analyze relative electric conductivity, activities of autioxidant enzymes and contents variation of endogenous ABA and IAA of maize blades, as well as Asr1 gene expression level by Real-time PCR. The relative electric conductivity of maize blades showed an increasing tendency, meanwhile, activities of SOD and POD were enhanced by exogenous ABA treatments among which SOD activity was significantly higher in ABA treatments of 15 mg L-1 and 25 mg L-1than in the control under low-temperature stress. Also the synthesis of endogenous ABA and IAA were increased significantly. The expression level of Asr1 gene significantly promoted in ABA treatments of 5, 15, and 25 mg L-1. ABA contant had a highly significant correlation with the relative expression of Asr1 gene and SOD activity and a significant correlation with POD activity. In conclusion the expression of Asr1 gene is mediated and regulated by ABA, the promotion of Asr1 gene expression also promotes the synthesis of endogenous ABA, and enhances antioxidant enzyme activities, resulting in improved maize resistance to low temperature. But the mediation and regulation of exogenous ABA have a concentration effect, showing promotion at low concentation and inhibition at high concentation.
低温是限制作物分布及其生长最重要的环境因素[1]。玉米是中国第一大粮食作物, 2014年种植面积达到3.69× 107 hm2。玉米生长期遭遇低温, 会引起植株体内一系列生理代谢反应, 进而影响产量和品质。黑龙江省地处高纬度, 低温冷害发生概率为28%, 严重低温冷害每5年发生一次, 低温导致玉米减产22%~34% [2]。近些年来, 随全球气候变暖, 黑龙江省成为全国气候变暖中心, 但气候的自然波动依然存在, 加上盲目北移和晚熟品种增加等人为因素, 低温冷害仍存在潜在威胁[3]。脱落酸(abscisic acid, ABA)是低温逆境下的重要信号因子, 对细胞的微管结构有保护作用。低温环境下时植物体内ABA大量累积, 对植物起到保护性作用, 植物抗冷性提高[4]。已有研究表明, 胁迫环境下小麦[5]、甘蔗[6]、水稻[7]、大麦[8]等作物施加外源ABA, 增加渗透调节物质含量, 使SOD和POD活性增强, 影响植物体内内源激素含量, 促进脱落酸的合成, 显著缓解胁迫效应。黄杏等[9]研究表明, 低温胁迫下, 外源ABA能有效降低甘蔗体内GA3含量, 提高ABA含量。杨东清等[10]研究表明, 高温处理下喷施外源ABA, 使小麦ZR含量降低, ABA含量显著提高。ABA亦被作为抗寒基因表达过程中重要的逆境信号分子及启动因子。水稻中Os-ASR基因在茎秆中受外源ABA诱导上调表达。我们课题组[11]在以往玉米低温逆境研究中, 已通过蛋白质双向电泳和肽指纹图谱技术分析筛选到一个低温上调表达的Asr1基因, ASR (the abscisic acid, stress, ripening)于1993年首次被发现[12], 迄今该基因家族成员在作物非生物胁迫中表达广泛[13, 14, 15]。Frankel等[16]研究发现, 在干旱条件下, Asr1基因在番茄叶片中大量表达, 增强其抗旱性。Chin等[17]用花椰菜花叶病毒35S启动子使转LLA23基因(LLA23是ASR基因家族中的一个成员)拟南芥种子对ABA的敏感性降低, 证明拟南芥体内的LLA23可以介导ABA信号的胁迫应答, 使植株表现出很强的抗干旱和抗盐能力。为探讨低温胁迫下外源ABA对玉米幼苗抗冷基因Asr1的调控机制, 揭示低温诱导过程中抗氧化酶活性变化及内在激素平衡状态与ABA调控关系, 从总体上把握了解 ABA 对冷胁迫下植株激素代谢及在信号作用下代谢基因调节过程的影响, 为揭示春玉米抗冷机理提供新的研究思路和理论依据。
据课题组前期筛选鉴定所得, 选用耐低温玉米品种久龙5号, 种子由黑龙江久龙种业有限公司提供。试验土壤取自东北农业大学香坊实验实习基地常规玉米地(黑土) 0~20 cm耕层。土样含全氮1.78 g kg-1, 速效钾183.48 mg kg-1、速效磷66.13 mg kg-1、有机质26.31 g kg-1、碱解氮122.34 mg kg-1、pH 6.88。试验所用信号物质脱落酸(abscisic acid, ABA)购自哈尔滨伟业生物工程有限公司, 纯度为95%~98%。
于2014年田间生产季即4月25日至4月29日在东北农业大学玉米研究所生理生态实验室进行室外盆栽种植。盆直径20 cm, 高23 cm, 每盆装土18 kg。室外正常培养至三叶一心, 选择无病、健壮、生长势均一植株, 将试验材料分为3组, 第1组清水常温, 为对照, 记为CK。第2组清水低温, 为低温处理, 记为C0。第3组在叶面喷施ABA的基础上低温处理, ABA浓度为5、15、25、35 mg L-1, 分别记为C5、C15、C25、C35, 每株喷施5 mL, 喷施诱导24 h后, 将植株放入光照培养箱中进行低温胁迫(昼温/夜温=14± 0.5℃/5± 0.5℃)处理[18], 光照时间为12 h, 光强为140 μ mol m-2 s-1, 低温胁迫0 d、2 d、4 d、6 d、8 d后取样。
1.3.1 电导率 选取玉米幼苗第2片叶片, 用清水轻轻擦洗表面玷污物, 然后分别用蒸馏水和去离子水清洗1~2次, 用滤纸将叶片表面水分擦拭掉, 用打孔器打取等量叶片(避开叶脉), 将同处理的样品混匀后分成3份, 分别将样品完全浸入装有20 mL去离子水的三角瓶中, 在室温下静置15 h后用电导仪(DDS-IIAGA型)测定浸泡液电导率R。然后将样品煮沸25 min, 待冷却至室温后用同样的方法测定各瓶溶液的电导率R1。
相对电导率(%) = R/R1× 100
1.3.2 保护酶活性 采用NBT (氮兰四唑)光化还原法测定超氧化物歧化酶(SOD)活性; 愈创木酚法测定过氧化物酶(POD)活性。
1.3.3 内源激素含量 采用酶联免疫吸附分析法(ELISA)[19], 从低温胁迫0 d开始取样, 选用倒数第2叶, 选择长势一致5株, 每个处理测定4次, 取平均值。使用Thermo Multlskan FC型酶标仪测定ABA (脱落酸)含量、IAA (生长素)含量。所用试剂盒由中国农业大学作物化学控制研究中心提供。
1.3.4 总RNA提取和Real-time PCR分析 取不同处理玉米叶片100 mg(从下数第2片叶和第3片叶), 于研钵中加入少量液氮碾磨至粉末, 移入1.5 mL离心管中, 采用 Trizol Kits抽提总RNA。采用多聚T 引物以及RevertAid第一条cDNA链合成试剂盒(Fermentas, 美国), 将约3 μ g总RNA反转录为cDNA, 用于Real-time PCR分析。
根据Asr1基因序列设计特异性引物(正向引物: 5′ -C TACGAGAAGCACGAGGCAAAGAAGG-3′ ; 反向引物: 5′ -GGAGGGATCAATCAGCCGAAGAAGT-3′ ); 根据玉米EF1α 设计内参特异性引物(正向引物: 5′ -TATGGTGAAGA TGGTTCCCACTAAG-3′ ; 反向引物: 5′ -TTGGCTCCAGT TGGGTCCTT-3′ )。所有引物均由生工生物工程(上海)有限公司合成。Real-time PCR反应程序为95℃预变性30 s后, 95℃变性5 s, 60℃退火30 s, 采集荧光, 循环40次。每个样本至少3个重复, 用2-Δ Δ Ct对样本基因进行表达差异相对定量分析。
1.3.5 干重 将鲜样放入烘箱内105℃杀青 30 min, 80℃烘干至恒重并称重。
1.3.6 数据分析 用Microsoft Excel 2007整理数据, 用DPS7.05软件进行数据方差分析和显著性检验, 差异显著性水平为0.05。
植物受到低温胁迫时, 质膜首先遭破坏。膜脂过氧化程度加剧, 细胞膜透性增大, 膜内电解质大量外渗, 相对电导率增大, 细胞膜功能和结构的损伤程度可以由质膜的透性变化显示。随着低温胁迫时间的延长, 不同浓度外源ABA处理的玉米幼苗相对电导率整体呈上升趋势, 最高值均出现在第8天, 表明随低温胁迫强度增加, 细胞膜损伤程度加剧(图1)。C35处理的增加幅度最大, C15最小。C35处理相对电导率低温胁迫第8天比0 d高91.0%, 而C15只增加了56.6%, 表明C35的细胞膜损伤程度最大, C15细胞膜损伤程度最小。低温进行2 d时, 低温下各ABA浓度处理差异不显著, 随胁迫天数增加, 即4~8 d时, C15和C25处理相对电导率最低, 且与C0处理差异显著。表明ABA的使用提高了幼苗的抗冷性, 质膜损伤减小。而在各低温胁迫时间段C35处理的相对电导率均高于C0处理, 差异不显著, 表明高浓度的外源ABA对玉米幼苗耐低温特性无提升作用, 反而出现一定伤害。
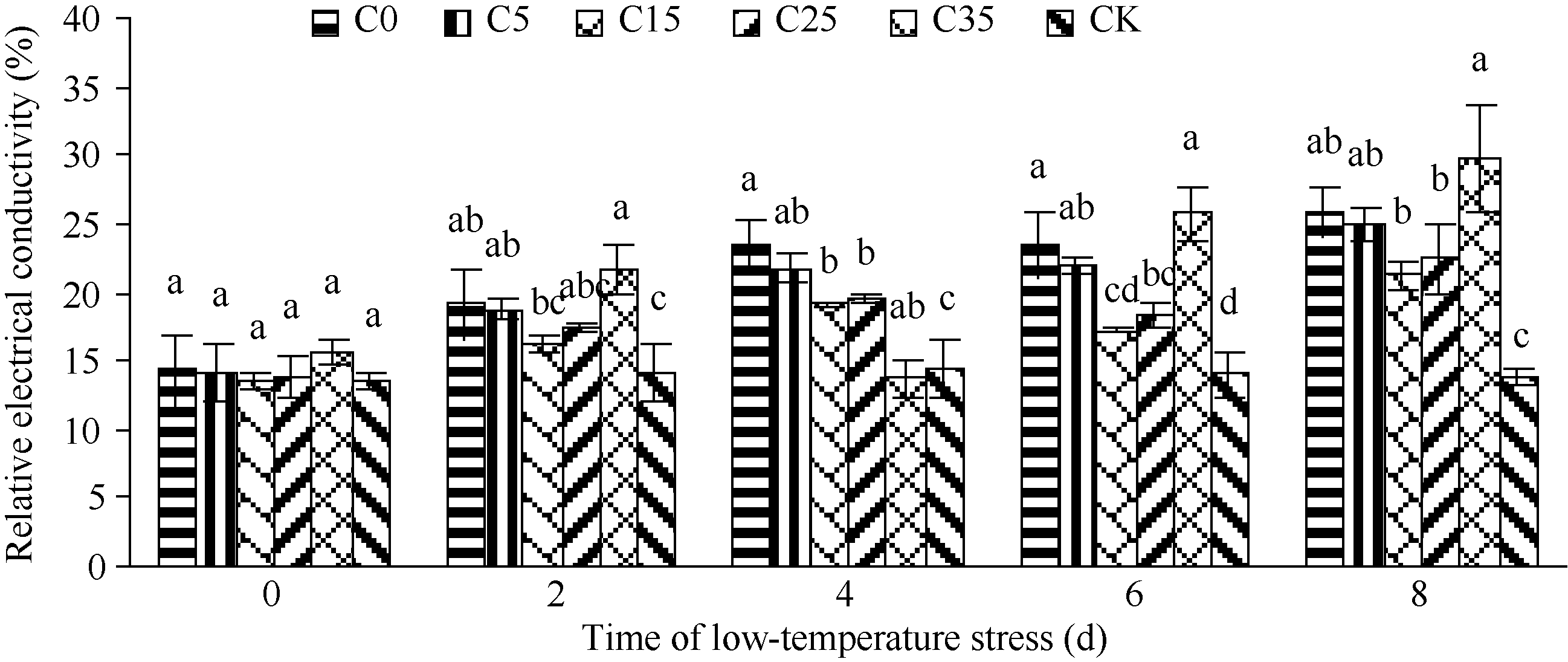 | 图1 外源ABA对低温胁迫下玉米苗期相对电导率的影响 C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。柱形图上不同字母表示处理间存在显著性差异(P< 0.05)。Fig. 1 Effect of exogenous ABA treatments on cell membrane permeability in maize seedlings under low temperature stress C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. Different letters above the bar mean significant difference between them (P< 0.05). |
抗氧化酶在植物体内具有清除超氧自由基、防止膜脂过氧化物伤害的作用, 保护膜结构、维持体内活性氧代谢的平衡, 一定程度上可使植物减缓或抵御逆境胁迫带来的伤害。
2.2.1 幼苗叶片超氧化物歧化酶(SOD)活性变化 由图2可见, SOD活性随着低温胁迫时间的延长逐渐升高, 在处理第6天达到最高, 随后下降。表明在短时间胁迫下玉米幼苗能够依靠自身的保护酶系统抵御低温的伤害, 但随着胁迫时间延长, 植物体内超氧负离子含量增加, 植株自身保护系统平衡被打破, SOD活性下降。适宜浓度的外源ABA可以提高低温胁迫下SOD活性, 胁迫2~8 d间, C15和C25处理SOD活性均显著高于C0处理, C5处理的作用不显著。C35处理玉米幼苗SOD活性在低温胁迫2 d后低于C0处理, 说明35 mg L-1的ABA浓度对玉米幼苗抗低温能力有抑制作用。
2.2.2 幼苗叶片过氧化物酶(POD)活性变化 不同ABA浓度处理的玉米幼苗POD活性变化趋势相似, 整体上POD活性均随着低温胁迫时间的延长先上升后下降, 活性最高值均出现在低温胁迫第6天(图3)。胁迫2~8 d间, C15和C25处理POD活性均显著高于C0处理。在低温胁迫后期, C35处理的玉米幼苗POD活性急剧下降, 说明在长时间低温胁迫下, C35处理对玉米幼苗抗冷性抑制增强。
2.3.1 ABA含量 玉米叶片内源激素ABA含量测定结果见表1。C0处理玉米叶片ABA含量随低温时间的延长有增加趋势, 处理8 d比0 d增加了29.2%, 增加幅度达到最大。与对照(CK)相比, 各低温处理玉米叶片ABA含量显著增加, 且随外施ABA浓度的提高, 叶片内源ABA含量有先升高后下降趋势, 差异达显著水平。其中C15处理和C25处理的含量增加最为显著, 最大值出现在低温处理第4 天和第8天, 低温8 d分别比对照(CK)增加了165.3%和159.4%。C5和C35浓度处理下也有一定的增加, 增幅较小。与C0处理相比, 各喷施ABA处理内源ABA含量均表现增加, 达到显著差异水平。说明低温胁迫下应用外源ABA提高了玉米幼苗內源ABA含量。
 | 图2 外源ABA对低温胁迫下玉米苗期SOD活性的影响 C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。柱形图上不同字母表示处理间存在显著性差异(P< 0.05)Fig. 2 Effect of ABA on SOD activity in maize seedlings under low temperature stress C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. Different letters above the bar mean significant difference between them (P< 0.05). |
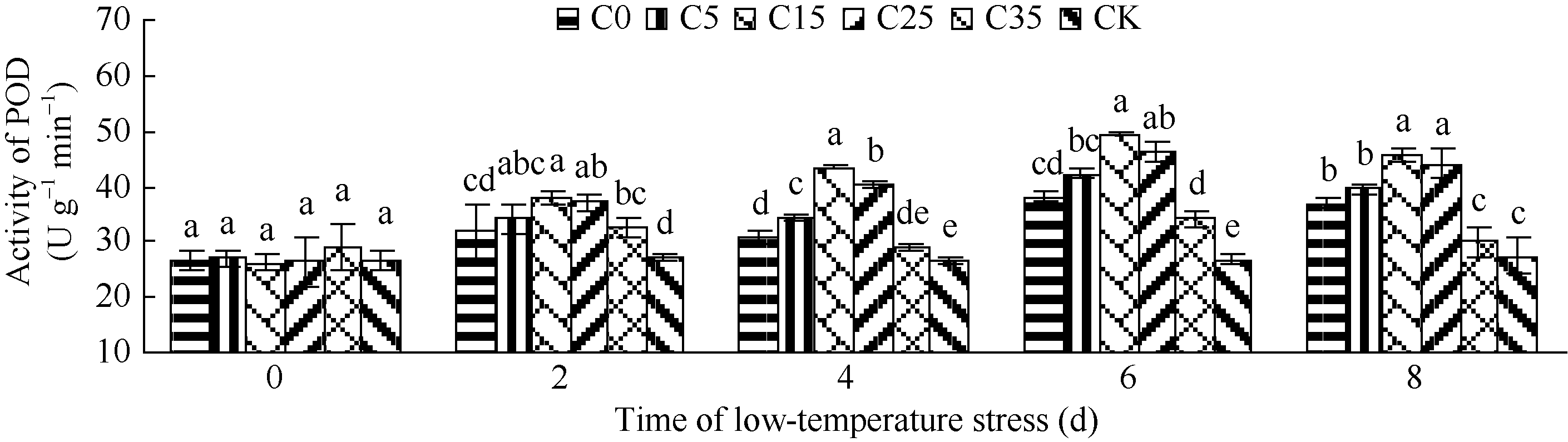 | 图3 外源ABA对低温胁迫下玉米苗期POD活性的影响 C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。注: 柱形图上不同字母表示处理间存在显著性差异(P< 0.05)。Fig. 3 Effect of ABA on POD activity in maize seedlings under low temperature stress C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. Different letters above the bar mean significant difference between them (P< 0.05). |
| 表1 外源ABA对低温胁迫下玉米苗期ABA含量的影响 Table 1 Effect of exogenous ABA treatments on ABA content in maize seedlings under low temperature stress (ng g-1 FW) |
2.3.2 IAA含量 IAA含量的增加可以提高细胞壁的可塑性, 增强细胞伸展性, 在低温胁迫下, 细胞膜不至于发生形变而破裂。随胁迫时间的延伸, 玉米叶片内IAA含量提高, 在低温胁迫第8天含量达到最大值。施加外源ABA后, 叶片内IAA含量有明显升高, 且随着ABA浓度的增加呈先升高后降低的趋势。C15处理在低温胁迫0 d比对照(常温条件下) IAA含量提高23.92%, 低温胁迫2 d、4 d、6 d、8 d, IAA含量分别比对照提高93.5%、72.2%、60.8%、57.2%, 说明低温胁迫对内源IAA含量提高的促进效应大于常温状态。低温胁迫第4天IAA增长量达到最大, C5、C25和C35各处理间的IAA含量变化不显著。说明适宜浓度ABA处理可增强玉米幼苗内源IAA的合成能力。
| 表2 外源ABA对低温胁迫下玉米苗期IAA含量的影响 Table 2 Effect of exogenous ABA treatments on IAA content in maize seedlings under low temperature stress (ng g-1 FW) |
不同浓度外源ABA处理后玉米叶片中Asr1基因均有表达(图4)。低温胁迫0 d, C5、C15、C25和C35处理的玉米幼苗叶片中Asr1基因相对表达水平较未施ABA处理均有不同程度上调, 其中C5、C15和C25处理的玉米幼苗叶片中Asr1基因相对表达显著上调, 分别为对照的24.85倍、25.56倍和23.26倍。这表明Asr1基因表达受外源ABA的调控, 在玉米中属于依赖ABA型, 喷施15 mg L-1和25 mg L-1外源ABA显著促进了Asr1基因的表达。各浓度ABA处理的玉米幼苗叶片Asr1基因表达水平与低温处理0 d相比, 2~6 d表达量不同程度下降, 8 d小幅上调表达。随低温胁迫时间延长, C35处理的玉米幼苗叶片中Asr1基因表达受到抑制。说明喷施高浓度35 mg L-1外源ABA玉米幼苗抗冷性基因Asr1表达受到抑制, 这与生理指标的测定结果吻合。
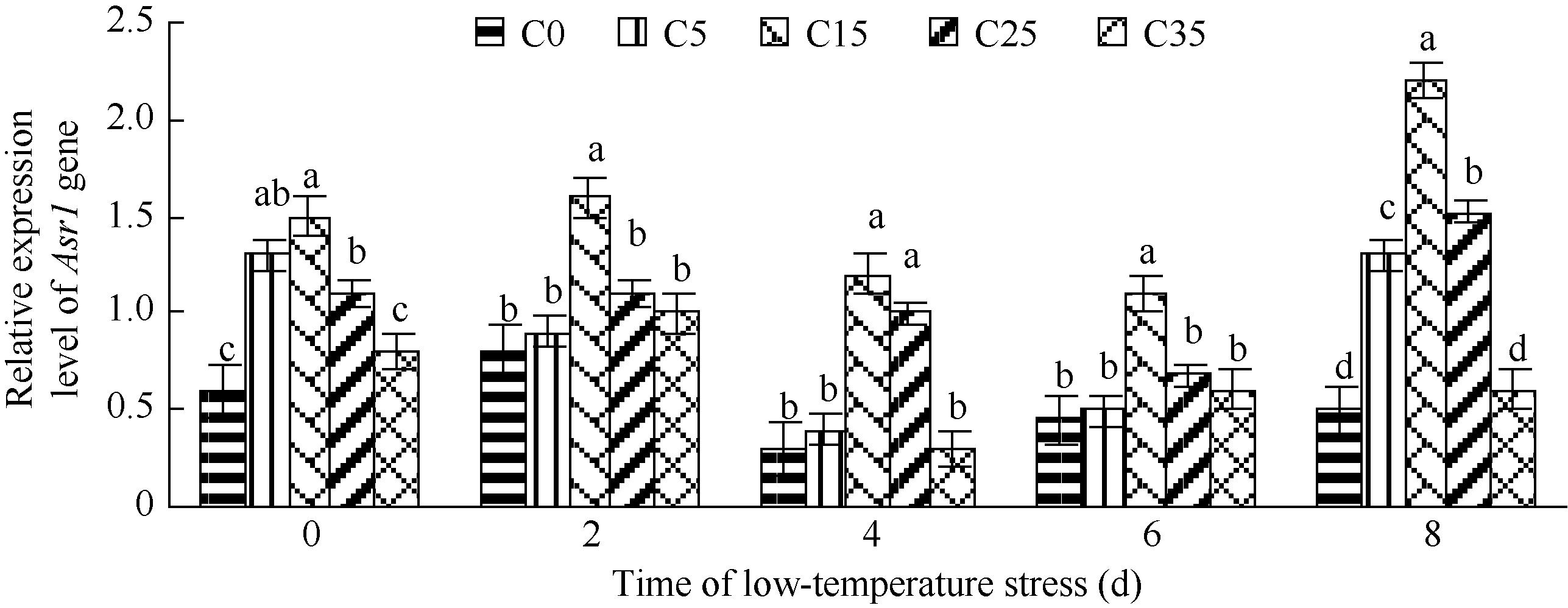 | 图4 低温胁迫下不同浓度外源ABA处理玉米幼苗Asr1基因相对表达量 C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。柱形图上不同字母表示处理间存在显著性差异(P< 0.05)。Fig. 4 Expression patterns of gene Asr1 in maize seedlings with different exogenous ABA treatments under low temperature stress C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. Different letters above the bar means significant difference between them (P< 0.05). |
玉米幼苗干重随着低温胁迫天数延长逐渐增加。C5、C15、C25处理与CK之间在低温胁迫期间幼苗干重差异不显著; C15处理与C0处理除低温胁迫2 d, 其余胁迫天数下幼苗干重差异均达到显著水平; C5、C25、C35处理与C0处理各时期幼苗干重差异不显著, 表明适宜的外源ABA, 缓解了低温的胁迫作用, 有利于增加幼苗干物质的积累。
由表4可知, 除电导率外, ABA含量与Asr1基因相对表达量、SOD活性均极显著正相关, 与POD活性显著正相关。IAA含量与Asr1基因相对表达量极显著负相关, 与电导率极显著正相关。玉米叶片干重与Asr1基因相对表达量呈极显著正相关, 与SOD活性极显著正相关, 与POD活性显著正相关。可见, ABA对抗冷基因Asr1的表达水平及玉米苗期抗低温能力具有更为积极的作用。
| 表3 玉米幼苗干重 Table 3 Corn seedling dry weight (g) |
| 表4 玉米叶片激素含量、叶片干重与Asrl基因相对表达量、电导率及抗氧化酶活性的相关系数 Table 4 Correlation coefficients of leaf hormone content and leaf dry weight with Asrl gene relative expression, electrical conductivity and antioxidant enzyme activities in maize |
ABA作为“ 胁迫激素” 在感受胁迫信号时, 能提高作物的抗逆性并起到重要作用[20]。对逆境下外源ABA的调控研究表明, 渗透胁迫条件下, ABA能降低水势, 提高渗透势, 增强渗透调节能力[21]。高温逆境下, 喷施外源ABA通过调节内源激素水平, 扩大籽粒库容量, 从而提高产量。本试验表明, 外源ABA通过对Asr1基因表达量以及内源激素水平的调控, 促进脱落酸合成, 减少细胞膜损伤, 进而提高玉米幼苗抗冷能力。
激素是植物生长发育的重要调节物质, 对逆境产生响应的激素信号分子, 广泛参与作物生理过程的调节。研究表明, ABA和植物的抗逆性密切相关, 外源激素ABA可以通过改变内源激素水平来调节植物内在生理代谢的各个环节[22, 23]。喷施外源ABA可以促进脱落酸在植物体内的合成和运输, 增加渗透调节物质的含量, SOD、POD等酶活性增强, 诱导抗寒基因表达, 从而提高植物抗寒能力[24]。本文ABA含量与抗氧化酶活性相关性分析也证明了此观点。本研究表明, 低温胁迫下作物的内源ABA含量增加, 而外施ABA处理会进一步促进内源ABA的合成, 使其含量达到更高水平, 这与Iqbal等[25]的研究结果相符, 且在柑橘[26]、小麦[25]等作物的研究中得到证实。同时施用ABA的C15和C25处理低温胁迫后抗氧化酶活性显著高于未施用处理, 也证实了ABA对玉米苗期抗性的提升作用, 但这种提升作用并不随着外源施用浓度的增加而正向增加, 反而表现出低促高抑的效应, 从文中C35处理ABA含量低于其他浓度处理即可看出, 可能外源激素类物质过高浓度抑制作物叶片及根系生长, 作物吸收能力减弱, 从而出现抗性减小, 具体原因还有待于对根系等营养器官的吸收能力的测试来分析。与之相比, IAA在植物抗冷性研究中较少。低温胁迫下IAA呈增加趋势, 外施ABA后效果更为显著, 这与白洁等[27]研究番茄的结果先下降后升高不一致, 这可能与低温的逆境胁迫相关, 启动了抗性基因的表达, 激素间拮抗作用减弱, 亦或者激素间的代谢与运输存在组织器官表达性, 本研究只关注叶片的激素效应而未测试玉米根系的含量, 根系数据的补充将更有效说明低温下IAA含量增加的原因。
不同物种中, ASR基因家族在不同的器官中表达, 玉米幼苗的根和叶中均有表达。由ASR基因家族编码的ASR 蛋白是生物体中广泛存在的一类与逆境有关的家族蛋白[28], 植物受到逆境胁迫(如低温、干旱、盐胁迫等)后, ASR蛋白大量表达, 可以缓解逆境引起的伤害[29]。Corté s等[30]研究发现菜豆的Asr1基因也与植物的耐旱性有着重要的关系。Goldgur等[31]用番茄的Asr1基因创制的转基因植株, 抗盐性增强。在水培番茄植株的根系和地上部分检测到Asr1 mRNA和蛋白在盐胁迫和水分胁迫下均上调表达, 表明Asr1基因的调控主要由RNA的转录或RNA的稳定性所决定[32]。在火炬松幼苗时期喷施外源ABA, 水分胁迫下松树根系中的lp3 (ASR基因家族成员)的表达部分受ABA的介导调控[33]。目前对于ASR基因家族的抗冷性研究较少。本研究表明, 低温胁迫下玉米幼苗叶片中Asr1基因的表达量增加, 推测可能与胁迫下ASR蛋白含量升高有关, 从而导致ASR蛋白在胞质中表现出分子伴侣活性, 在解冻周期中能稳定一定数量的蛋白质, 以避免它们因反复冻结而引起的变性, 最终提高玉米幼苗的耐冷性。本研究同时也证明外源ABA的施加可以促进Asr1基因表达量的增加, 并随ABA浓度的增加Asr1基因表达量先增加后降低。ABA诱导的Asr1基因表达量的提升能够加快ASR蛋白的合成, ASR蛋白的积累对植物在胁迫条件下起到潜在保护作用, 这对玉米低温胁迫研究具有重要意义[34]。相关分析也表明, 依赖于ABA介导的Asr1的表达, 也促进了内源ABA的合成, 抗氧化酶活性加强, 提升了应用ABA后玉米的抗低温能力。
此外, 幼苗叶片干重与生理指标相关分析表明, ABA对玉米幼苗抗低温胁迫表现出调节能力, 15 mg L-1浓度外源ABA的缓解低温胁迫的作用表现最强。且调节能力呈现一定的浓度效应, 即小于35 mg L-1的外源ABA处理时, 内源激素含量与Ars1基因的表达量均随ABA浓度增大而升高, 而施加更高浓度的外源ABA, 并没有一直缓解这种胁迫反应, 而表现出一定的抑制作用。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|