
{kind=link}
{kind=link}
水分亏缺对小麦灌浆中后期穗部光合特性和14C-同化物转运的影响
[米慧聪1  , 谢双泽
, 谢双泽2 , 李跃1 , 丁寒2 , 吕金印1, * ]
, 谢双泽]
|
|


第一作者联系方式: E-mail:mihuicong@163.com, Tel: 18220827092
为探讨水分亏缺对小麦灌浆中后期穗部同化物转运的影响, 利用14CO2同位素示踪技术, 对于2个抗旱性不同小麦品种的盆栽试验, 测定中度水分亏缺下小麦穗部光合特性及灌浆中后期碳同化物的转运。结果表明, 花后20 d, 中度水分亏缺下水地品种郑引1号旗叶和穗部净光合速率( Pn)分别下降50.2%和19.9%, 旱地品种普冰143分别下降33.7%和12.8%, 后者显著小于前者。14CO2示踪试验表明, 花后15~20 d籽粒中14C-同化物快速积累, 花后25 d达到最高值; 灌浆中后期(花后15~20 d)穗部苞片中14C-同化物向外快速转运, 灌浆末期(25 d)碳同化物已彻底转移。成熟期, 中度水分亏缺下小麦籽粒中14C-同化物积累高于正常供水处理, 且旱地品种普冰143积累量高于水地品种郑引1号。中度水分亏缺下, 郑引1号产量下降36.7%, 高于普冰143的23.2%。适度水分亏缺对旱地品种穗部净光合影响较小, 并促进了灌浆中后期穗部苞片中碳同化物的向外转运, 可能是维持旱作小麦稳产的生理基础。
The objective of this study was to understand the effect of water deficit on photosynthetic and assimilate translocation in wheat spike during mid- to late-filling stage by using two pot-cultured14C-labelled varieties differing in drought tolerance. Under moderate water deficit, the net photosynthetic rate ( Pn) of flag leaf and spike at 20 days after anthesis (DAA) decreased by 50.2% and 19.9% in Zhengyin 1 (drought sensitive) and by 33.7% and 12.8% in Pubing 143 (drought tolerance), respectively. Obviously, the decrease of photosynthetic capacity in the drought-sensitive variety was greater than that in the drought-resistant variety in response to water stress. The14C-assimilates accumulated rapidly in grains during 15-20 DAA and reached the peak at 25 DAA. Simultaneously, the14C-assimilates in glume, lemma, and awn had a quick outward transportation during 15-20 DAA, and completely transferred to grains at 25 DAA. Compared with normal water condition, moderate water deficit resulted in significantly higher14C-assimilates in grains at maturity. The14C-assimilate accumulation in Pubing 143 was significantly higher than that in Zhengyin 1, and the yield loss caused by drought stress was 23.2% in Pubing 143 and 36.7% in Zhengyin 1. Compared with drought-sensitive variety, drought-tolerant variety received less influence of moderate water deficit on spike Pn and stronger assimilate translocation from spike bracts to grain. This might be the physiological basis of stable yield in drought-resistant wheat variety.
干旱是限制我国小麦北方主产区小麦产量的主要非生物胁迫因素之一[1]。水分亏缺导致叶片光合能力快速下降, 影响碳同化物转运, 降低小麦产量[2]。已有研究表明, 小麦穗部苞片、穗下节、叶鞘、茎等非叶器官对水分胁迫不敏感, 是干旱条件下籽粒灌浆期间碳同化物的重要来源[3, 4]。穗部具有重新固定籽粒呼吸释放的CO2、接受外界光照和CO2等明显生理优势[5, 6, 7], 即使在干旱条件下仍能保持相对稳定的光合能力[3, 8, 9]。
Gebbing和Schnyder[5]通过减源试验, 提出面包小麦穗部光合作用对籽粒的贡献可达22%~45%; Sanchez- Bragado等[10]利用13C标记方法, 研究发现小麦地方品种穗部对籽粒的贡献高于现代品种。适度干旱可以显著促进花后茎鞘中14C同化物的运转[11]。干旱条件下穗部光合对籽粒灌浆的贡献增加[12, 13, 14], 适度干旱胁迫促进了小麦花前和灌浆前期穗部同化物向籽粒的转运[15, 16]。Jia等[17]对水分亏缺下不同小麦品种灌浆速率进行Logistic方程拟合表明, 花后15 d灌浆速率达到高峰。花后18 d小麦植株开始衰老, 籽粒经灌浆脱水在花后24 d生理成熟[3]。灌浆中后期相对于旗叶穗部器官具有衰老延迟性[3], 但水分亏缺下不同抗旱性小麦灌浆中后期穗部光合及碳同化物的转运规律还不十分清楚。
本试验研究水分亏缺下不同抗旱性小麦穗部光合特性及灌浆中后期碳同化物的分配率, 探讨穗苞片灌浆中后期光合及籽粒的调运差异, 为揭示旱作小麦的稳产高产机制提供依据。
选用不同抗旱性冬小麦(Triticum aestivum L.)品种, 其中普冰143由西北农林科技大学培育, 2004年9月经过陕西省审定, 为耐旱品种, 并适合旱地种植; 郑引1号原产意大利, 由河南省农业科学院于20世纪80年代引进, 为水地品种。
2014年10月至2015年6月在西北农林科技大学北校区遮雨网室进行盆栽土培试验。供试土壤为关中平原农田耕作层(0~22 cm)风干红油土, 土壤最大田间持水量时净含水量为29.2%。2014年10月18日播种, 设正常供水和中度水分亏缺两种处理, 土壤含水量分别为最大田间持水量的70%~75%和45%~50%。抽穗期开始控水, 每天按标准称重法补水, 每处理重复25盆。
1.2.1 旗叶和穗部净光合速率(Pn) 采用便携式光合仪(LI-6400 XT, LI-COR, USA)在花后10、15、20、25 d晴天9:00— 11:00测定旗叶和穗部净光合速率。采用光合仪测定旗叶光合速率, 配置的LED红蓝光源, 光强为1000 μ mol m-2 s-1, 大气CO2浓度390 μ mol mol-1, 每处理重复4次。采用特制的圆柱形叶室与便携式光合仪(LI-6400 XT, LI-COR, USA)配套测量穗部, 依据当时自然光变化配合使用人工红蓝光源, 使测定时光照强度为1000 μ mol m-2 s-1 [15], 根据Teare和Peterson[18]和裘昭峰等[19]的方法计算穗表面积, 分别测穗颖片及芒的表面积, 穗面积 = 穗长 × 穗宽 × 3.8; 芒面积=顶3小穗的芒长× 直径× 结实小穗数× π 。
1.2.2 14CO2标记测定同化物转运分配 在灌浆中后期(花后15 d)选择长势及花期一致的小麦, 于上午9:00用聚乙烯塑料袋套住穗部, 用注射器注入5 mL 14CO2, 标记强度为81.77× 104 Bq L-1, 立即封口。光合1 h后用NaOH溶液回收残留于袋中的14CO2, 去除塑料袋。
于花后15、20、25、30 d, 每次采样4株, 将旗叶、颖片、外稃、芒、籽粒等器官分开, 在105℃杀青, 70℃烘箱烘干至恒重磨碎, 称取50 mg, 加0.5 mL 60% HClO4和0.7 mL 30% H2O2。80℃消化4 h, 加10 mL闪烁液, 暗处过夜, 测定总放射性活度。以上操作均重复3次。用多功能液体闪烁计数系统(LS-6500, Beckman, USA, 计数效率≥ 95%), 测定14C cpm值, 通过计数效率校正, 将cpm值校正为dpm值。
1.2.3 农艺性状指标及水分利用效率(WUE) 小麦成熟后, 分别统计小穗数、可育小穗粒数、穗粒数、穗粒重、千粒重, 统计籽粒产量, 计算收获指数。WUE=每盆籽粒产量(g) /每盆总耗水量(kg)。
用Microsoft Excel 2013及SPSS 20.0软件统计分析数据, 用Duncan’ s检验差异显著性。
在灌浆中后期2个供试小麦品种旗叶Pn快速下降(图1-A)。与对照相比, 中度水分亏缺处理下2个品种旗叶Pn下降显著(P< 0.05), 尤其灌浆后期(花后20 d), 水地品种郑引1号下降50.2%, 旱地品种普冰143下降33.7%, 前者下降幅度明显大于后者。
2个品种灌浆中后期穗部Pn呈缓慢下降趋势(图1-B)。与正常供水相比, 花后20 d, 郑引1号穗部Pn下降19.9%, 大于普冰143的12.8% (P< 0.05)。中度水分亏缺处理下穗部净光合速率下降幅度显著小于旗叶, 与旗叶相比, 穗部保持相对较高的净光合速率, 表现出一定的光合优势, 且郑引1号穗部光合速率受水分亏缺影响大于普冰143。
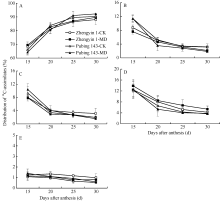
14CO2示踪试验表明, 籽粒中14C-同化物花后15~20 d快速积累, 花后25 d (标记后10 d)达到最高值(图2-A), 中度水分亏缺下旱地品种普冰143籽粒中14C-同化物分配率为91.1%, 而水地品种郑引1号为86.5%。与正常供水相比, 花后30 d (收获期)中度水分亏缺下普冰143籽粒14C-同化物分配率增加1.9%, 而郑引1号增加1.7%。
中度水分亏缺促进了灌浆中后期(花后15~20 d )穗部苞片中14C-同化物向外转运。与正常供水相比, 花后30 d (收获期)水分亏缺下普冰143和郑引1号颖壳14C-同化物分配率降幅分别为1.3%和0.2% (图2-B)。花后30 d, 中度水分亏缺下普冰143和郑引1号外稃中14C-同化物分配率分别为1.6%和2.0%, 而正常供水处理分别为2.0%和3.3% (图2-C), 中度水分亏缺下水地品种外稃中14C-同化物的滞留明显高于旱地品种。而花后30 d (收获期), 中度水分亏缺下普冰143和郑引1号芒中14C-同化物分配率分别为3.9%和5.4%, 略高于正常供水处理的3.7%和4.2% (图2-D), 可能与灌浆期芒的早衰有关。收获时中度水分亏缺下水地品种芒中14C-同化物分配率明显高于旱地品种, 表现出少量的滞留现象。尤其指出的是, 花后15 d, 芒中14C-同化物分配率(12.2%~13.9%)明显高于颖壳(7.6%~11.4%)和外稃(7.9%~1.4%)中(图2-B, C, D), 灌浆中后期旗叶中有少量14C-同化物(0.5%~1.4%) (图2-E)。中度水分亏缺促进了穗部苞片(颖壳、外稃、芒)碳同化物向外转运, 且旱地品种穗部14C-同化物的滞留小于水地品种。
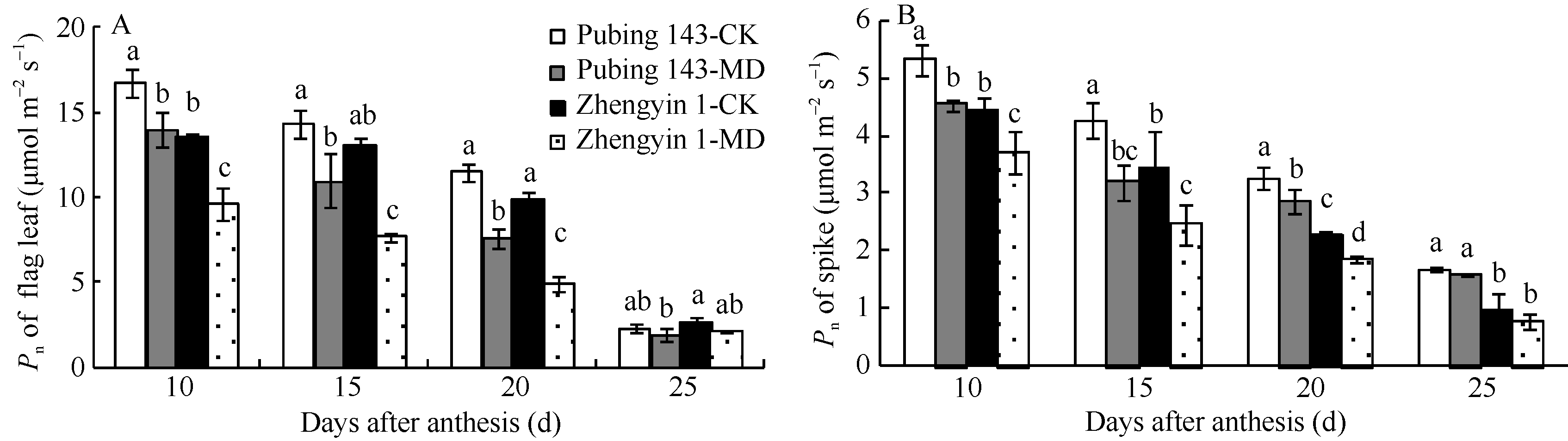 | 图1 中度水分亏缺对小麦灌浆中后期旗叶(A)和穗部(B)净光合速率的影响 CK: 正常供水; MD: 中度水分亏缺。数据为3次生物学重复的平均值± 标准差, 误差线上不同字母表示相同生育期处理间有显著差异(P< 0.05)Fig. 1 Effect of water deficit on net photosynthetic rate (Pn) of flag leaf (A) and spike (B) after anthesis in wheat CK: normal water supply; MD: moderate water deficit. Data are means ± standard deviations of three biological repeats. Different letters above the error bars indicate significant difference among treatments on the same sampling day (P < 0.05). |
 | 图2 中度水分亏缺对灌浆中后期小麦籽粒(A)、颖壳(B)、外稃(C)、芒(D)和旗叶(E)中14C-同化物分配率的影响 CK: 正常供水; MD: 中度水分亏缺。数据为3次生物学重复的平均值± 标准差。Fig. 2 Effect of water deficit on the distribution of 14C-assimilates accumulated in grain (A), glume (B), lemma (C), awn (D), and rachis (E) of wheat after anthesis CK: normal water supply; MD: moderate water deficit. Data are means ± standard deviations of three biological repeats. |
中度水分亏缺下旱地品种普冰143穗粒重和千粒重降低的幅度分别为30.1%和10.0%, 而水地品种郑引1号分别为39.6%和16.2% (表1), 前者降幅显著小于后者。中度水分亏缺下, 普冰142和郑引1号产量下降显著(P< 0.05)。普冰143收获指数降幅为2.9%, 小于郑引1号的15.5%。中度水分亏缺下, 两品种的WUE变化显著(P< 0.05), 普冰143升高10.3%, 而郑引1号则下降11.1% (表2)。表明中度土壤干旱对旱地小麦产量和收获指数的影响小于水地品种。
| 表1 干旱处理对不同抗旱性品种产量相关性状的影响 Table 1 Yield of different wheat varieties under different treatments |
| 表2 水分亏缺对不同抗旱性品种产量、生物量、收获指数和水分利用效率的影响 Table 2 Effects of water deficit on yield, biomass, harvest index and water use efficiency (WUE) in different drought-resistant varieties |
小麦穗部光合作用对籽粒产量具有重要贡献[9, 20]。而干旱等不利条件加速了叶片的衰老进程[15], 降低了旗叶光合产物对籽粒的贡献率。近年研究表明, 逆境下小麦穗部表现出更强的干旱适应能力[21], 而叶片对干旱胁迫更为敏感[22]。本研究中, 中度水分亏缺下两种不同抗旱性小麦灌浆中后期穗部净光合速率缓慢下降, 而旗叶净光合速率呈快速下降趋势。且水分亏缺下水地品种郑引1号穗部、旗叶净光合速率降幅均大于旱地品种普冰143。Jia等[17]认为, 水分亏缺条件下穗部比旗叶具有光合优势。本研究也有类似结果, 适度干旱对穗部光合影响小于对旗叶的影响, 逆境下穗部表现出光合持久性。
干旱诱导了小麦早衰[23, 24], 缩短了生育期, 增加了茎鞘等临时库14C-同化物的转运[13, 24, 25], 并且穗部对籽粒的贡献增大[26]。Evans等[27]报道, 干旱下小麦穗部光合碳同化物对籽粒产量的贡献率增幅为34%~43%, 且水分胁迫下小麦穗部光合对籽粒贡献率高于正常供水处理[11]。本研究发现, 中度水分亏缺促进了灌浆中后期小麦穗部碳同化物向籽粒的转运(图2-A), 旱地品种籽粒调用量相对较高; 同时, 中度水分亏缺促进了旱地品种小麦穗部苞片(颖壳、外稃)的14C-同化物向外转运(图2-B, C), 减少了滞留。本研究还佐证了Bidinger等[26]穗部碳同化物的再转运对籽粒灌浆至关重要的结论。Abebe等[28]认为芒是穗部主要的光合器官, 大麦中芒对穗部光合贡献可达90%[29]。本研究中芒14C-同化物分配率明显高于颖壳和外稃(图2-D)。Olugbemi[30]用14C同位素标记试验, 证实芒中约有99%的光合同化产物转运至着生该芒的小穗中。因此, 穗部器官中芒的光合固碳作用不容忽视。另外, 2个品种旗叶中存在少量来自穗部的14C-同化物(图2-E), 可能是干旱使旗叶早衰的情况下, 穗部调运碳同化物以维持其代谢活动。中度水分亏缺下, 旱地品种产量下降幅度小于水地品种, 可能是由于旱地品种灌浆中后期穗部苞片碳同化物滞留少, 更多的光合产物运输到籽粒中。本研究结果表明, 中度水分亏缺加速了小麦叶片衰老, 降低了灌浆期旗叶光合能力, 而穗部器官光合能力可维持在相对较高水平, 且对旱地品种影响较小; 适度干旱处理促进了旱地品种籽粒对穗部苞片碳同化物的调运, 增加了籽粒干物质的积累。这可能是旱地品种相对高产的原因之一, 充分挖掘小麦穗部等非叶器官光合固碳潜力, 可为实现小麦高产提供重要途径。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|