



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻早衰突变体 psls1的基因定位及克隆
[黄雅敏1, *  , 朱杉杉
, 朱杉杉1, * , 赵志超1, * , 蒲志刚2 , 刘天珍1 , 罗胜1 , 张欣1, * ]
, 朱杉杉, 赵志超, 蒲志刚]
|
|


第一作者联系方式: 黄雅敏, E-mail:fanxianxinglou@126.com; 朱杉杉, E-mail:zhushanshan@caas.cn; 赵志超, E-mail:zhaozhichao@caas.cn
水稻叶片早衰直接影响作物的光合效率, 减少产量, 降低品质。深入研究早衰的分子机制对控制和延缓衰老具有重要意义。本文报道了一个早衰突变体 psls1(premature senescence leaf with spots)的基因定位及克隆结果。突变体在发育到七叶期以后, 叶片自下而上叶绿素含量下降, 过氧化氢过量积累, 突变体叶片逐渐黄化至枯萎; 其他农艺性状如株高、分蘖数、主穗长、结实率和穗粒数也相应变差。电镜观察进一步发现, psls1衰老叶片中叶绿体降解、类囊体基粒片层模糊, 嗜锇颗粒明显增多。遗传分析表明, psls1受1对隐性基因控制, 利用 psls1× IRAT129杂交组合F2分离群体中的1690个早衰个体, 将基因 PSLS1定位在第7染色体分子标记ZS-3和ZS-8之间89 kb的范围内。测序研究发现, 区间内一个编码铁氧还依赖的谷氨酸合酶基因 LOC_Os07g46460的第2外显子末位的G被替换为A, 导致转录本的错误剪切, 突变体的cDNA缺失了57 bp碱基片段。在突变体中该基因表达量下降, 谷氨酸合酶活性降低, 其产物谷氨酸含量显著下降、其他氨基酸代谢紊乱。水培低氮处理下可诱发突变体 psls1早衰。研究结果表明, 由于 PSLS1突变使得谷氨酸合酶失活, 氮代谢异常而导致突变体 psls1早衰。
, ZHU Shan-Shan, ZHAO Zhi-Chao, PU Zhi-Gang
Leaf early senescence directly reduces its photosynthetic capacity, decreasing crop yield and grain quality. It is of great importance to identify novel mutants and characterize their molecular and physiological mechanisms. In this paper, we reported gene mapping and cloning of a psls1 mutant (premature senescence leaf with spots) in rice. The mutant showed early senescence symptoms, including decreased chlorophyll content, over-accumulated H2O2, yellow-withered leaf and increased dead cell from bottom to top of plant after the 7-leaf stage. Moreover, the plant height, tiller number, panicle length, and the number of grains per panicle were significantly lower in psls1. We observed the degradation of chloroplast, unclean thylakoid and numerous osmiophilic granules in psls1leaves by transmission electron microscope (TEM). Genetic analysis demonstrated that the phenotype of psls1 was determined by a single recessive gene. Using genetic population derived from psls1× IRAT129, the psls1 locus was mapped in a 89 kb region flanked by markers ZS-3 and ZS-8 on chromosome 7, containing 12 putative open reading frames (ORFs). Sequence analysis revealed a single-base substitution occurred in the genomic sequence of LOC_Os07g46460, which led to a 59 bp deletion in its cDNA, and therefore, predicted LOC_Os07g46460( PSLS1), encoding a ferredoxin-dependent glutamate synthase as the candidate gene. The mRNA expression level of mutated PSLS1 decreased sharply, resulting in reduction of glutamate synthase activity and abnormal amino acid metabolism in psls1. Under low nitrogen treatment, the senescence phenotype of psls1could occur as early as at the 3-leaf stage. These results indicated that psls1 senescence phenotype might be associated with the loss of glutamate synthase activity and the abnormal amino acid metabolism.
叶片是水稻的重要光合器官。早衰叶片过早丧失同化作用, 植株生长发育迟缓、结实率低、灌浆不足, 导致品种的产量减少、品质下降。反之, 合理延长叶片寿命, 可使水稻产量增加。有研究表明, 水稻叶片延迟一天衰老, 可使水稻增产1%左右[1]。因此, 探究早衰发生的生理和分子机制, 控制和延缓衰老对提高品种的产量潜力具有重要意义。
植株衰老性状由一个精细复杂的调控网络所控制。在分子水平上, 控制衰老的基因大体可以分为以下4类: (1)激素信号转导与合成相关基因。乙烯(ETH)合成及相关信号传导基因突变会影响植株衰老, 乙烯不敏感突变体ein1和ein2表现出延迟衰老的表型[2, 3], 过表达EIN3则造成早衰表型[4]。其他激素如脱落酸(ABA)[5]、茉莉酸(JA)[6]、细胞分裂素(CK)[7]、水杨酸(SA)[8]等也与植株衰老相关。(2)调节基因。涉及衰老性状的转录因子一般会调节植株的衰老, OsNAP受ABA的特异性诱导, 通过直接调控叶绿素降解、营养再转运及其他衰老相关基因的表达, 调控叶片衰老进程[5]。SUB1转录因子有3个ERF结构域, 在持续黑暗条件下, 该基因通过限制乙烯合成, 阻止叶绿素和碳水化合物的降解以及衰老相关mRNAs的积累来延缓水稻叶片衰老[9]。WARK类转录因子OsWARK42通过抑制ROS基因OsMT1d的表达, 加速叶片衰老[10]。(3)蛋白酶或物质转运代谢相关基因。ES1编码SCAR-LIKE PROTEIN2, 该蛋白是CAMP受体蛋白抑制子的组分, 其突变体不仅表现早衰, 而且失水速率提高[11]。OsABC1-2编码ABC1激酶, 该基因参与黑暗胁迫的叶片衰老调控[12]。NYC1编码叶绿素b还原酶, 在突变体中抑制了叶绿素b和光捕获复合体LHC II的降解, 植株叶片表现出功能性的滞绿表型[13]。水稻滞绿SGR基因, 定位于类囊体膜上, 通过调节脱镁叶绿酸加氧酶的合成与降解, 引起衰老过程中叶绿素的降解; ABA可以促进该基因的表达, 细胞分裂素则抑制其表达; 过表达植株的类囊体膜的片层的数量减少, 且叶绿素含量降低[14]。(4)防御反应相关基因。植株防御反应与衰老也存在一定的联系。拟南芥hys1 (hypersenescence 1)突变体在暗光照条件下表现出早衰性状, 由于HYS1基因也是病程相关基因[15], 表现出防卫反应相关的表型[16]。
氮素是植物生长发育过程中重要营养元素, 通常施氮可以延长叶片寿命, 缺氮加快衰老, 但氮素吸收与衰老关系的分子机制仍有待阐明。植物吸收无机态氮, 再利用谷氨酰胺合酶(GS)/谷氨酸合酶(GOGAT)循环将无机态氮转化为有机氮[17]。高等植物中有两类GOGAT, 即NADH-GOGAT和Fd-GOGAT, 它们分别以NADH和铁氧还蛋白(Fd)为辅酶[18]。在拟南芥中Fd-GOGAT占叶片总谷氨酸合酶活性的96%, 根中总谷氨酸合酶活性的68%[19, 20]。拟南芥有2个Fd-GOGAT (AtGLU1和AtGLU2)基因, GLU1主要在叶片氮素初吸收途径及光呼吸氮的再吸收中起作用, 受光照和蔗糖的特异性诱导。而GLU2则主要负责根中氮素的初吸收[21]。同样地, 水稻也有两个Fd-GOGAT同源基因[22, 23], 即编码完整蛋白的OsFd-GOGAT1和非活性的OsFd-GOGAT2。抑制谷氨酸合酶基因家族的表达可导致水稻分蘖减少、单株产量下降, 叶片中部分游离氨基酸、叶绿素、糖等含量相应下降[24]。OsFd-GOGAT1主要在水稻植株的叶片中表达, 其表达量受光照和氮素浓度影响[22, 23, 25], 但对其功能研究较少, 也未见OsFd-GOGAT1突变引起叶片衰老的报道。本研究通过对一个早衰突变体psls1的图位克隆, 发现编码谷氨酸合酶的PSLS1基因的突变造成植株早衰。该结果为进一步研究氮素代谢异常引起叶片衰老的分子机制奠定了基础。
水稻早衰突变体psls1, 来源于田间育种群体自然变异株系, 其野生型为籼稻品系P22382。2013年配制杂交组合psls1× IRAT129(粳稻), 经南繁加代后, 于2014年正季将F2群体种于北京昌平基地, 共取样1690株早衰个体用于基因定位。分别考察psls1突变体和野生型各10株的株高、分蘖数、结实率、穗粒数、千粒重等相关农艺性状。
1.2.1 叶绿素含量测定 测定叶绿素含量参照Wu等[26]方法, 选取幼苗期未早衰的第4片真叶和拔节后期严重早衰植株的绿色心叶及黄化叶片, 利用紫外可见分光光度计(Beckman DU800 USA)分别测定叶绿素a、叶绿素b及类胡萝卜素含量。
1.2.2 活性氧检测 分别取野生型和psls1的新鲜叶片(心叶和倒三叶), 用试剂盒(碧云天公司)测定早衰植株叶片过氧化氢含量变化。同时采用二氨基联苯胺(diaminobenzidine, DAB)染色法检测叶片中H2O2积累量[27], 采用台酚蓝(trypan blue)染色法鉴定叶片死亡细胞[28], 观察并照相。
1.2.3 游离氨基酸含量测定 在抽穗期, 选取突变体和野生型的上部叶片, 液氮磨样冷冻干燥后称取约1 g样品(设置3个生物重复), 委托中国农业科学院饲料研究所中心分析室检测游离氨基酸含量。
参照Dong等方法[29], 利用透射电镜(Transmission Electron Microcopy, TEM)观察叶绿体超微结构。分别取psls1植株上部微黄、下部严重黄化的叶片, 野生型对应部位的叶片, 固定于含有2.5%的戊二醛固定液中, 抽真空, 室温固定12 h, 切片后以电镜观察。
在F2群体中分别选取10株正常单株和10株突变体单株, 按CTAB法构建基因池[30]。参照Gramene (http://ensembl.gramene.org/)中公布的日本晴和93-11的全基因组序列寻找插入/缺失(Indel)位点, 利用Primer3 (http://bioinfo.ut.ee/primer3-0.4.0/)设计Indel分子标记, 由上海立菲生物技术有限公司合成引物。10 μ L的PCR扩增体系含10 ng DNA模板、4 nmol引物、0.5 mmol dNTP、1 μ L 10× buffer (不含Mg2+)、15 mmol MgCl2和0.5 U Taq DNA聚合酶。PCR条件为94℃ 5 min; 35个循环的94℃ 30 s、58℃ 30 s、72℃ 30 s; 72℃ 5 min。扩增产物经8%非变性聚丙烯酰胺凝胶电泳, 银染后观察。
从水稻基因组注释数据库(http://rice.plantbiology. msu.edu/)中下载候选基因的DNA和CDS序列, 设计测序引物, 分别对突变体psls1和野生型中的候选基因进行PCR扩增并测序, 用DNAMAN软件分析, 确定突变位点。
用RNA试剂盒(TIANGEN公司)提取水稻叶片RNA。活性氧相关基因表达分析选择在突变体出现黄化表型时期取样测定。PSLS1的基因表达分析分别取幼苗期、分蘖期、拔节期和抽穗期的水稻叶片。再用Prime Script II 1st Strand cDNA Synthesis Kit 试剂盒(TaKaRa公司)合成cDNA。以水稻多聚泛素酶基因(UBQ)为内参, 引物正向序列为5° -GCTCC GTGGCGGTATCAT-3° , 反向序列为5° -CGGCAGTT GACAGCCCTAG-3° , 在7500HT Real-time PCR仪(Applied Biosystems)上进行Real-time PCR扩增, 采用2-Δ Δ CT方法计算基因表达量(荧光定量引物见附表2)。
利用购自上海研生公司的植物谷氨酸合成酶ELISA试剂盒, 参照说明测定GOGAT酶活性。
以野生型cDNA为模板扩增LOC_Os07g46460的CDS, 引物为F: 5° -TCTGCACTAGGTACCTGCA GATGGCCACGCTCCCACG-3° /R: 5° -ATGGATCCGT CGACCTGCAGCTTCGCCGATTGTACTGTTGTTG-3° , 将扩增片段连接到pCUbi1390-FLAG载体上, 农杆菌介导遗传转化。利用Crispr技术构建干扰载体, 正反向引物为F: 5° -AGATGATCCGTGGCACACGC TCCCACGTGCGTCGGGTTTTAGAGCTATGC-3° /R: 5° -GCATAGCTCTAAAACCCGACGCACGTGGGAGCGTGTGCCACGGATCATCT-3° , 遗传转化粳稻品种Kitaake的胚性愈伤组织。
参照Liu等[31]水稻营养液配制方法, 设置不同氮浓度梯度NH4NO3 (2 mmol L-1、1 mmol L-1、0.5 mmol L-1、0.1 mmol L-1、0.01 mmol L-1)培养液, 待野生型种子萌发后水培。pH调至5.4~5.6, 每隔3 d换一次营养液。
psls1幼苗期生长正常, 七叶期以后叶片自下而上逐渐黄化褪绿至枯萎(黄化由叶尖开始向内蔓延并伴有黄褐色小斑点); 生长后期突变体衰老明显加快, 植株整体呈黄褐色枯死状态(图1)。在成熟期, 与野生型相比, psls1株高、分蘖数、主穗长、穗粒数和千粒重均显著下降(表1)。
与野生型相比, psls1苗期幼叶以及早衰表型出现后的绿色心叶中色素含量并没有降低, 但对于严重早衰黄化叶而言, 其叶绿素a、叶绿素b和类胡萝卜素含量均显著下降(图2)。
与野生型相比, 在早衰严重的psls1植株, 无论是绿色心叶还是黄化叶的H2O2含量都显著增加, 衰
老叶片经DAB染色后出现棕褐色斑点。经台酚蓝染色后呈现大量蓝色斑点, 表明psls1叶片中存在大量死亡细胞(图3)。对12个活性氧相关基因的表达定
量分析结果表明, 3个与活性氧清除有关的基因OsAPX2、OsCATC、OsPOX1在psls1植株中表达量显著下降(图4)。
 | 图1 野生型和突变体psls1表型 A: 野生型(WT)和突变体psls1幼苗期表型; B: 野生型(WT)和突变体psls1分蘖初期表型(下部叶片见图C左, 上部叶片见图C右); D: 野生型(WT)和突变体psls1拔节后期表型(上部叶片见图E)。Fig. 1 Phenotype of wild type and psls1 mutant A: phenotype of WT andpsls1 at seedling stage; B: phenotype of WT andpsls1 at early tillering stage (lower leaves and upper leaves are displayed in C); D: phenotype of WT andpsls1at late elongation stage (upper leaves are displayed in E). |
| 表1 野生型与psls1突变体农艺性状比较 Table 1 Comparison on agronomic traits between wild type and psls1 mutant |
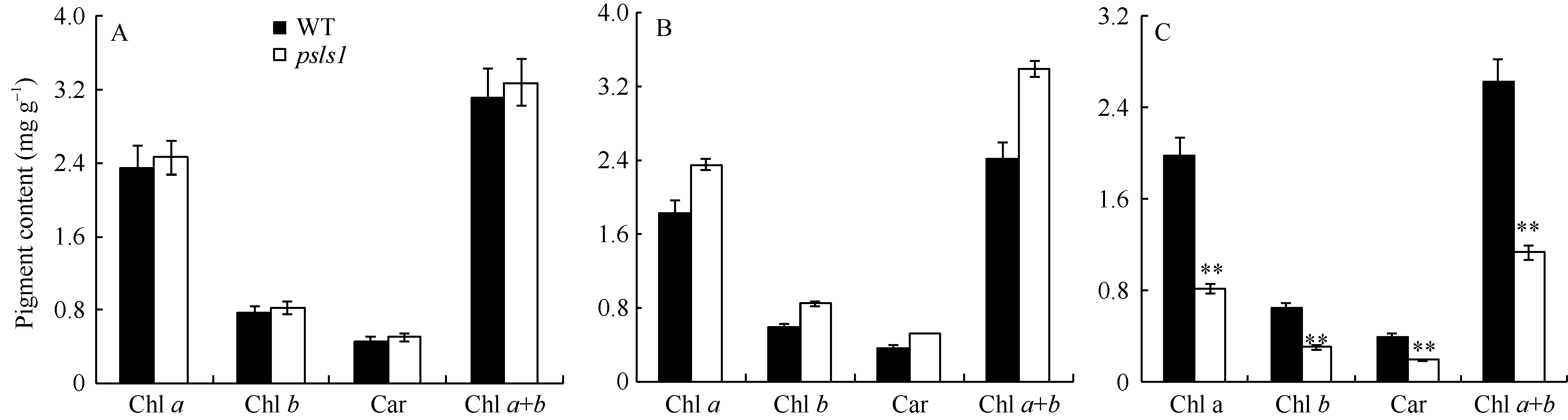 | 图2 野生型(WT)和突变体psls1光合色素含量 A: 野生型与突变体psls1幼苗(未出现早衰)第4片真叶光合色素含量; B: 野生型WT与突变体psls1 (严重早衰表型)绿色心叶光合色素含量; C: 野生型与突变体psls1抽穗前(严重早衰表型)倒三叶光合色素含量。Fig. 2 Photosynthetic pigment content of wild type (WT) and psls1 mutant A: pigment content of the fourth leaf in seedling (non chlorisis); B: pigment content of the green leaf before heading stage (serious chlorisis); C: pigment content of the 3rd leaf from the top before heading stage (serious chlorisis). |
 | 图3 野生型(WT)与突变体psls1组织化学染色及过氧化氢检测 A: 分蘖后期野生型(绿叶)和突变体psls1 (黄化叶)DAB染色; B: 野生型(绿叶)和突变体psls1 (黄化叶)台酚蓝染色; C: 抽穗前期心叶及倒三叶过氧化氢含量。Fig. 3 Histochemical staining and detection on hydrogen peroxide content of wild type (WT) and psls1 mutant A: DAB staining of wild type (green leaf) and psls1(chlorotic leaf); B: Trypan Blue staining of wild type (green leaf) and psls1 (chlorotic leaf); C: H2O2 content of green leaf and chlorotic leaf (the 3rd leaf from the top) before heading stage. |
 | 图4 野生型(WT)与突变体psls1 (明显早衰)叶片活性氧相关基因表达量分析Fig. 4 Quantitative analysis of ROS genes in wild type (WT) and psls1 mutant (serious chlorisis) leaves |
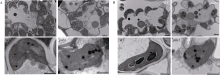
透射电镜观察发现, 野生型(上、下部均为绿色)下部叶片叶绿体数量比上部叶片略微减少, 但叶绿体结构较完整。而突变体psls1上部微黄叶片叶绿体数量仅约为野生型的一半, 形状不规则, 膜界面不清晰, 类囊体基粒片层模糊, 嗜锇颗粒明显增多; 下部严重衰老叶片叶绿体数量更少、嗜锇颗粒增多更明显(图5)。这表明psls1黄化早衰伴随着叶绿体的降解和细胞的死亡。
 | 图5 野生型WT与突变体psls1叶肉细胞超微结构 A: 野生型WT与突变体psls1上部叶片叶绿体结构(psls1微黄); B: 野生型WT与突变体psls1下部叶片叶绿体结构(psls1严重黄化)。Fig. 5 Ultrastructure of mesophyll cell in mutant psls1 and the wild type A: chloroplast structure of upper leaves in wild type and psls1 (psls1slightly yellowing); B: chloroplast structure of lower leaves in wild type and psls1 (psls1serious chlorisis). |
为研究早衰突变体的遗传特性, 将psls1与叶片正常衰老的粳稻品种IRAT129杂交, F1植株叶片表型正常, F2出现叶片衰老和正常两种表型。在随机调查的1350个植株中, 野生型植株和突变型植株分别为1033株和317株, 符合3∶ 1 (χ 2 = 1.583, P > 0.05)的分离比例, 说明psls1突变性状受1对隐性基因控制。
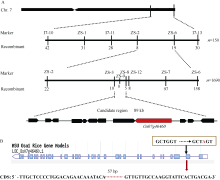
从F2群体中选取10株具有早衰表型的个体构建DNA混池。选用实验室保存的176对均匀分布于12条染色体上的InDel和SSR引物, 对混池进行连锁分析, 之后对150个F2早衰个体进行验证和初定位, 最后将PSLS1初步连锁到第7染色体长臂末端分子标记ZS-1与I7-13之间。根据http://www. gramene.org/网站上的基因序列信息, 在I7-11和I7-13标记之间又新开发了4对分子标记ZS-2、ZS-3、ZS-6、ZS-7 (见附表1), 进一步对793个早衰单株分析, 将目标区段定位于分子标记ZS-3和ZS-7之间, 利用进一步开发的分子标记ZS-8、ZS-9、ZS-12和897个F2新增加的早衰单株, 最终将PSLS1基因定位在标记ZS-3和ZS-8之间, 物理距离为89 kb的范围内(图6-A)。根据水稻基因组注释数据库(Rice Genome Annotation Project), 在精细定位的89 kb的区间中包含12个注释基因(表2)。
测序结果表明, 在psls1中, 仅有1个编码谷氨酸合酶(ferredoxin-dependent glutamate synthase)的基因LOC_Os07g46460的第2个外显子末位发生了碱基G→ A的替换(图6-B)。因此, 初步认定LOC_Os07g46460为可能的候选基因, 暂命名为PSLS1。
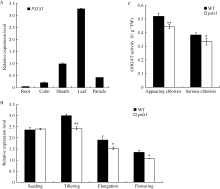
PSLS1在水稻根、茎、叶、叶鞘、穗部均有表达, 在叶片中表达量较高(图7-A); PSLS1基因的表达水平在野生型与psls1幼苗的叶片中无差异, 待分蘖期早衰表型出现后, 突变体叶片中PSLS1的表达量显著下降(图7-B), 由于PSLS1编码一个铁氧化-还原依赖的谷氨酸合酶Fd-GOGAT, PSLS1具有GOGAT活性, psls1发生早衰后谷氨酸合酶活性显著降低(图7-C), 这与PSLS1为目的基因的推测结果一致。
OsFd-GOGAT是谷氨酸合酶, 此酶可在还原型铁氧还蛋白存在的条件下, 催化谷氨酰胺生成谷氨酸, 而谷氨酸是合成其他氨基酸和含氮化合物的原料[18]。抑制谷氨酸合酶基因的表达导致叶片游离谷氨酸含量降低[24], 为了检测PSLS1基因突变与谷氨酸代谢有无关系, 我们测定了野生型和psls1抽穗期绿色叶片中游离氨基酸的含量(表3)。结果发现在突变体psls1中Glu、Ser、Gly、Ala、Val、Ile含量显著下降, 而Asn、Pser (磷酸化丝氨酸)、Thr、Cys、Phe、His、Orn、Lys、Arg含量反而增加。表明突变体中氨基酸代谢紊乱。
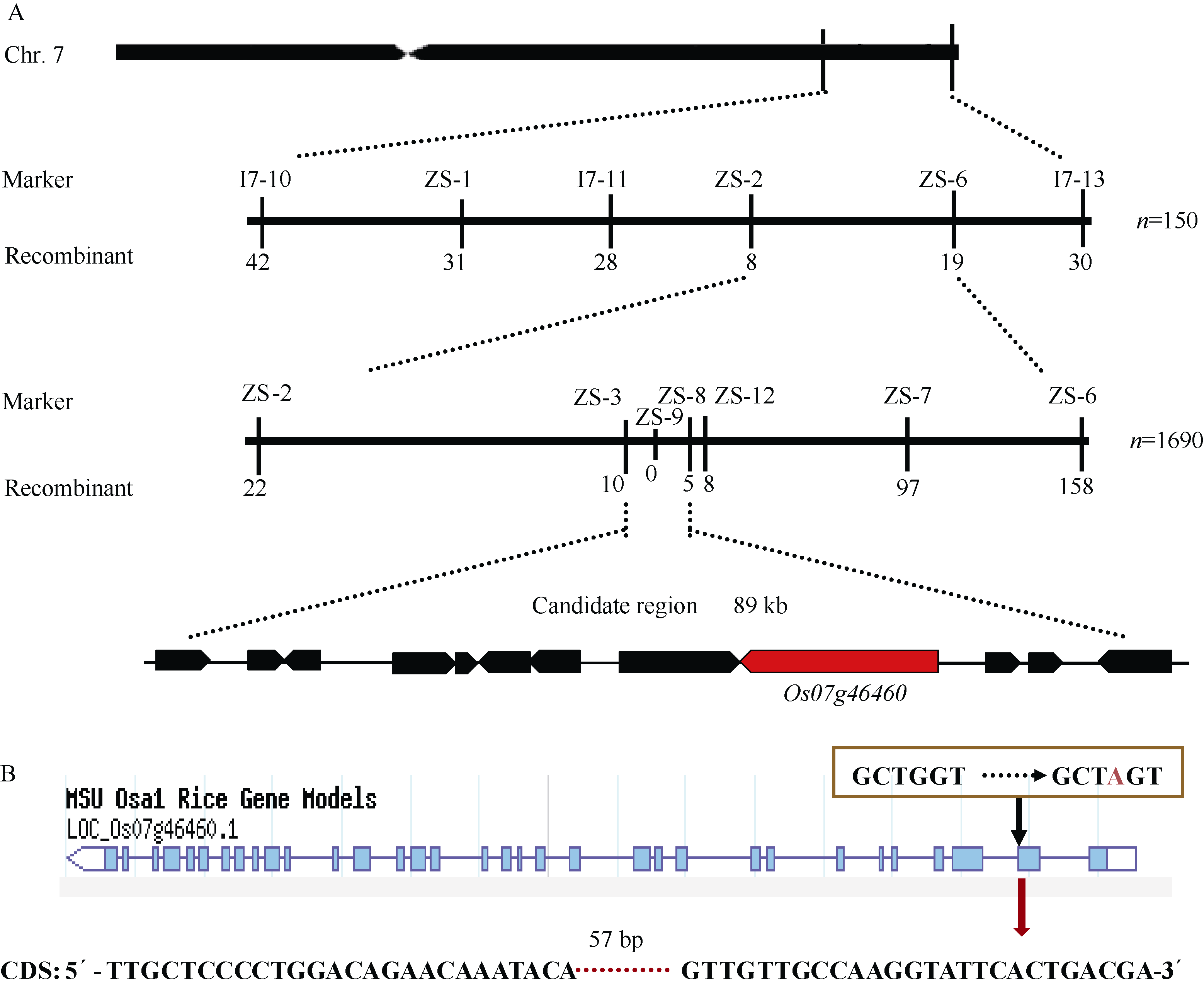 | 图6 PSLS1的图位克隆 A: PSLS1基因的精细定位(横线上方的标记表示精细定位中用的SSR和Indel标记, 下方的数字表示该标记下的重组交换株数)。B: 定位区间基因LOC_Os07g46460基因组G→ A的改变导致编码区存在57 bp碱基缺失。Fig. 6 Map-based cloning of PSLS1gene A: fine mapping of PSLS1(the numerals above bar represent SSR and Indel markers used and the numerals below bar represent recombinants); B: the sequence variations in candidate region, the mutation of single base in LOC_Os07g46460leads to base delection of 57 bp in CDS. |
| 表2 候选区间内基因预测 Table 2 Putative genes in the delimited genomic region |
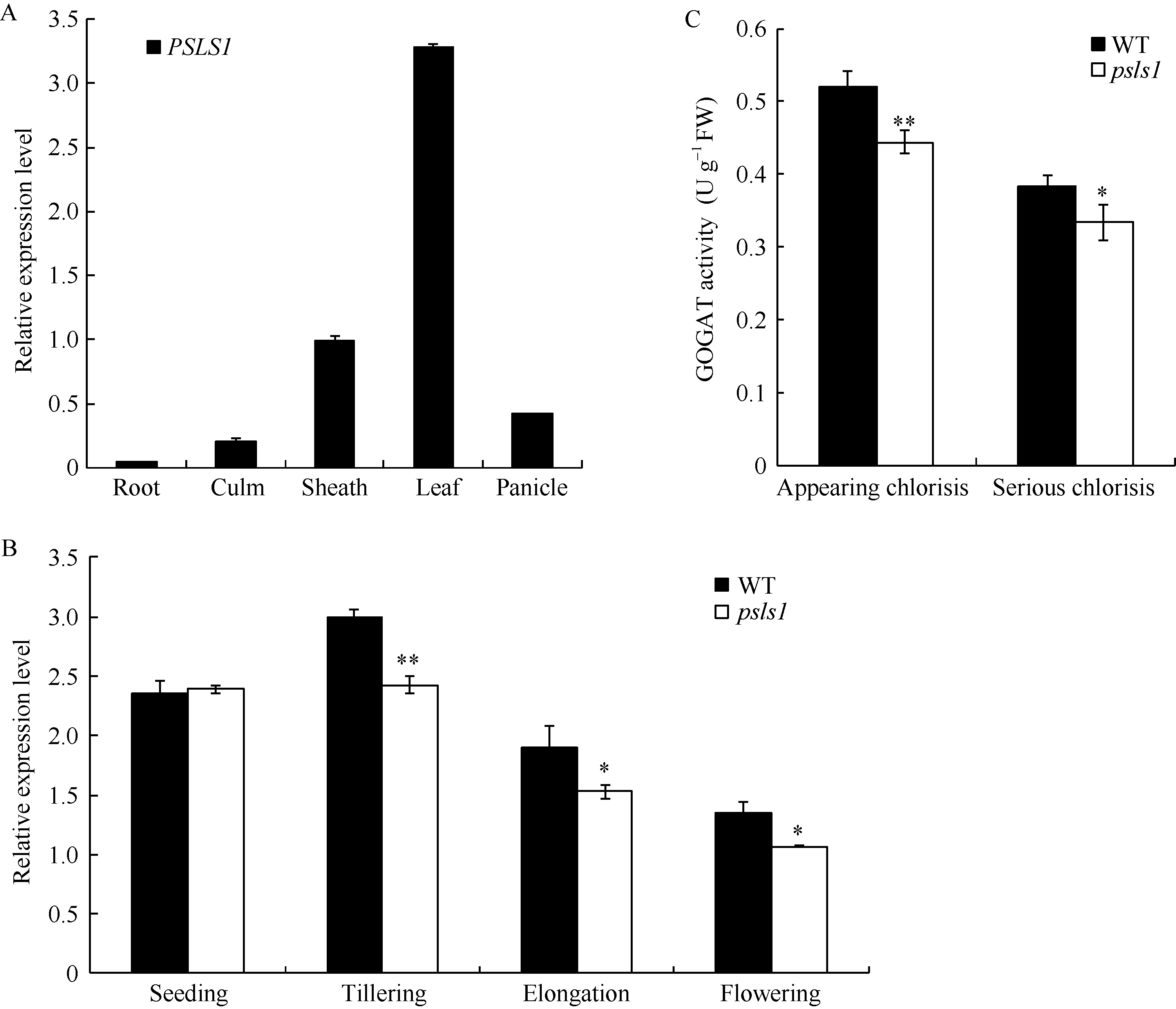 | 图7 PSLS1表达量分析和谷氨酸合酶活性测定 A: PSLS1组织特异性表达分析; B: 不同生长时期PSLS1表达量; C: psls1刚出现早衰表型(appearing chlorisis)和早衰表型严重期(serious chlorisis)谷氨酸合酶活性。Fig. 7 Relative expression level of PSLS1and GOGAT activity A: tissue-specific expression of PSLS1; B: expression of PSLS1 in different periods; C: GOGAT activity of wild type and psls1(in appearing chlorisis and serious chlorisis stages respectively). |
| 表3 野生型与psls1叶片中游离氨基酸含量 Table 3 Free amino acid content in leaves of wild-type and psls1mutant (μ g g-1) |
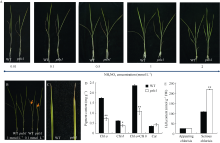
水培条件下, 不同浓度氮素外施处理3周后, 低氮处理加速psls1叶片衰老, 0.5 mmol L-1 NH4NO3处理下, psls1的第2叶完全衰老, 第3叶略微黄化, 而WT仍保持绿色; 随着氮浓度的下降, 在0.1 mmol L-1 NH4NO3以及更低的浓度处理下, 除第2叶完全衰老外, psls1的第3叶片褪绿黄化现象较0.5 mmol L-1 NH4NO3处理更加明显, 野生型的第2叶片也出现了黄化, 这些结果表明低氮处理能诱发早衰, 并加速psls1早衰的发生。在0.1 mmol L-1 NH4NO3处理下, 突变体植株叶绿素含量显著下降、H2O2积累高(图8)。
 | 图8 低氮处理下野生型和psls1表型及生理变化 A: 不同浓度梯度氮素处理下野生型WT和突变体psls1表型(从左到右NH4NO3浓度依次为0.01 mmol L-1、0.1 mmol L-1、0.5 mmol L-1、1 mmol L-1、2 mmol L-1); B: 1 mmol L-1 (左)和0.1 mmol L-1(右) NH4NO3条件下野生型WT和突变体psls1表型(红色箭头处见图C); D: 低氮处理(0.1 mmol L-1 NH4NO3)下四叶期(第3叶)色素含量; E: 低氮处理(0.1 mmol L-1 NH4NO3)三叶期(appearing chlorisis)和四叶期(serious chlorisis)过氧化氢含量。Fig. 8 Phenotype and physiological changes of wild type and psls1 mutant under low nitrogen condition A: phenotype of wild type and psls1 mutant under different concentrations of nitrogen (NH4NO3) (concentration from left to right in turn are 0.01 mmol L-1, 0.1 mmol L-1, 0.5 mmol L-1, 1 mmol L-1, and 2 mmol L-1); B: phenotype of wild type and psls1 mutant under 1 mmol L-1 and 0.1 mmol L-1 NH4NO3 (the red arrow mark the leaf magnified in C); D: pigment content of wild type and psls1at four-leaf stage (the third leaf) under low nitrogen treatment (0.1 mmol L-1); E: H2O2 content of wild type and psls1 at three-leaf stage (appearing chlorisis) and four-leaf stage (serious chlorisis) under low nitrogen solution condition (0.1 mmol L-1). |
本文报道了一个水稻早衰黄化突变体psls1表型分析及精细定位结果。在田间条件下, 突变体在七叶期后叶片自下而上逐渐褪绿黄化至枯萎, 叶绿素降解、活性氧大量积累、氨基酸代谢紊乱、细胞死亡, 导致株高、分蘖、穗粒数、千粒重都显著下降, 严重影响水稻的品质和产量。遗传分析表明该早衰表型由单隐性基因控制。利用杂交组合psls1× IRAT129的F2群体, 将候选基因定位在第7染色体长臂末端物理距离89 kb的范围内。测序发现psls1中基因LOC_Os07g46460第2个外显子末位发生了一个由G到A的替换, 导致转录本错误剪切, cDNA缺失了57 bp碱基片段, 初步判断LOC_ Os07g46460为候选基因。LOC_Os07g46460编码一个铁氧还蛋白依赖的谷氨酸合酶(Fd-GOGAT)。
植物主要通过GS/GOGAT循环将无机氮转化为有机氮, 水稻谷氨酸合酶被认为具有提高水稻氮素利用率的潜力, 抑制NADH-GOGAT1和NADH- GOGAT2编码基因的表达均可以导致地上部干重、氮总含量降低[24, 32]。Lam等[18]研究认为在非豆科植物中, NADH-GOGAT定位于非光合组织如根的质体中, 功能主要是氮素的初吸收以及氨基酸代谢产生氮的再吸收。Fd-GOGAT则定位于叶绿体中, 参与植物光呼吸过程中氨的再吸收。本研究结果表明, 候选基因PSLS1主要在叶片中表达。相对于野生型, 突变体叶片中PSLS1的表达量明显偏低, 谷氨酸合酶活性下降, 叶绿素含量降低, 活性氧含量增加, 氨基酸代谢紊乱, 衰老提前发生。低氮条件能导致突变体的衰老提前发生。这些证据支持了PSLS1可能为目的基因的结论。
植物从土壤中吸收的硝态氮首先被硝酸还原酶(NR)和亚硝酸还原酶(NiR)还原为氨(NH4+), 然后在谷氨酰胺合酶的催化下结合谷氨酸生成谷氨酰胺, 进一步在谷氨酸合酶的作用下与α -酮戊二酸结合形成谷氨酸[33]。谷氨酸是植物体内其他氨基酸和酰胺的合成前体, 通过氨基交换作用最终形成植物可以直接利用的氮素化合物[34]。Fd-GOAGT/GS2是植物光呼吸NH4+再吸收的主要机制[35], 在拟南芥和大麦中, Fd-GOAGT缺陷植株均严重黄化、光呼吸受阻[21, 36, 37]。在拟南芥中, 虽然Fd-GOAGT对植株的氮素利用和吸收起到至关重要的作用[18, 20, 21, 24], 但是到目前为止, 尚未有Fd-GOAGT缺乏引起水稻早衰的报道。近年来水稻中也已经鉴定了大量的SAGs, 分别涉及叶绿素合成降解、蛋白合成降解、激素等途径, 但是有关氮素吸收与水稻衰老关系的分子机制仍有待阐明。Fd-GOGAT在光呼吸氨的再吸收过程中发挥着重要作用, 在本研究中, 突变体PSLS1基因的突变引起水稻叶片黄化早衰, 但是, Fd-GOGAT如何在光呼吸GS/GOGAT循环中起作用, 如何通过提高Fd-GOGAT酶活性来提高水稻氮素利用率进而减缓植株生长后期叶片过早衰老对水稻产量造成的损失有待进一步研究。
水稻突变体psls1是一个新的早衰黄化突变体。该突变体过氧化氢积累升高、叶绿体降解、细胞坏死, 谷氨酸合酶活性降低, 谷氨酸含量下降, 氨基酸代谢紊乱。突变体LOC_Os07g46460基因的第2个外显子末位一个G到A的替换导致转录本错误剪切, cDNA缺失57 bp碱基片段, 进而引起PSLS1表达量下降, 是造成突变体psls1黄化早衰的直接原因。
| 附表1 本研究所使用的定位引物 Supplementary table 1 Markers used in mapping |
| 附表2 本研究所用荧光定量引物 Supplementary table 2 Markers for Real-time PCR in this study |
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|