{kind=link}
适合不同产量的环境下油菜高收获指数的产量构成因素分析
[卢坤1, *  , 申鸽子
, 申鸽子1, * , 梁颖1 , 明联2 , 贺斌3 , 琳梅3 , 张烨1 , 彭柳1 , 李加纳1, * ]
, 申鸽子, 梁颖]
|
|


第一作者联系方式: 卢坤, E-mail:drlukun@swu.edu.cn; 申鸽子, E-mail:578223874@qq.com
收获指数偏低是制约油菜籽粒产量和产油量进一步提高的瓶颈。为解析适合不同产量的环境下油菜高收获指数的构成因素及形成机制, 本研究选择在高产环境下的云南临沧和一般产量的长江流域上游主产区重庆北碚均能正常生长和成熟的321份甘蓝型油菜品种(系)为材料, 分析其产量相关性状的变异以及两地间的差异, 利用相关分析和通径分析研究籽粒产量收获指数(YHI)、产油量收获指数分别与17个产量组成性状的关系。结果表明, 云南高产环境下, 油菜高产的主要原因是光照充足、昼夜温差大, 导致生物产量高、角果多、每角粒数多, 特别是二次分枝角果对产量贡献较大。主序和一、二次分枝的角果数与产量收获指数在重庆均呈显著正相关, 而云南环境下主序角果数与产量收获指数呈显著负相关。主序和一、二次分枝的每角粒数均以云南环境下极显著高于重庆环境下, 且在两种环境下, 主序和一次分枝每角粒数都与产量收获指数和产油量收获指数呈显著正相关, 表明每角粒数需要充足的光合产物积累及高效的籽粒填充效率来保证。主序、一次和二次分枝千粒重云南均低于重庆, 表明寡日照区域的油菜会减少籽粒数, 以保证部分种子的干物质填充。云南环境下, 各部位千粒重与产量收获指数和产油量收获指数均呈显著正相关, 而重庆环境下的相关性不明显, 说明在光照充足条件下, 光合产物转运能力是导致千粒重差异的主要原因。综合分析表明, 在光照充足的环境下, 主序角果数和单株经济产量是提高YHI的关键; 而寡日照环境下YHI构成复杂, 必须将主序和一次分枝的产量组分有机结合, 且严格控制生物产量才能实现YHI的提高。
, SHEN Ge-Zi, LIANG Ying
Low harvest index is a bottleneck for further improvement of seed yield and oil production in rapeseed ( Brassica napus). A total of 321 B. napus accessions with normal growth and maturation at both the super high yield production area Lincang, Yunnan province and the major production area of the upper Yangtze River Basin, Beibei, Chongqing were chosen in this study. Variance of yield-related traits and their differences between two locations were compared, and the relationship between yield harvest index (YHI), oil production harvest index (OHI) and 17 yield component traits were also investigated by correlation and path analyses. At Yunnan, the main reasons for high yield of B. napus were adequate illumination, big diurnal temperature difference, sufficient photosynthate, more biomass, higher number of siliques per plant and seed number per silique, especially the number of siliques on the secondary branches, which might play key roles in improvement of B. napus yield. The silique number on the main inflorescence, primary and secondary branches was significantly and positively correlated with YHI at Chongqing, but the reverse relationship was observed between YHI and silique number on main inflorescence at Yunnan. The number of seeds per silique on the main inflorescence, primary and secondary branches at Yunnan were significantly higher than those at Chongqing, and the number of seeds per silique on the main inflorescence and primary branch showed significant or extremely significantly positive correlation with YHI and OHI simultaneously at both two cultivated areas, suggesting that sufficient accumulation of photosynthate and high grain filling efficiency are necessary for large number of seeds per silique. Thousand-seed weights on the main inflorescence, primary and secondary branches at Yunnan were all lower than those at Chongqing, suggesting that once the photosynthate is insufficient, the seed number per plant will decrease and partial seeds will become the priority for seed-filling in B. napus, to ensure the quality of offspring seeds. Under the high yield production condition of Yunnan, thousand-seed weights on three different parts showed significant or remarkably significantly positive correlation with YHI and OHI, while those at Chongqing were not, indicating that thousand-seed weight is determined by the transport capacity of photosynthate under the condition of sufficient illumination. In summary, the number of siliques on the main inflorescence and seed yield per plant are two key factors for increasing YHI under adequate illumination environment. But the composition of YHI is more complicated under weak illumination environment. Thus, higher YHI could be achieved when the yield components on the main inflorescence and primary branch are organic ally integrated and biological yield is restrained.
作物产量提高主要有两种途径, 一是在生物学产量不变的情况下提高收获指数(harvest index); 二是在收获指数一定的情况下提高生物学产量[1, 2]。众多研究表明, 近年来世界水稻(Oryza sativa)、小麦(Triticum aestivum)和大麦(Hordeum vulgare)等主要作物产量的增长主要得益于收获指数的改良, 收获指数已成为评价作物品种选育效果的重要内容[3, 4, 5]。相当多的学者研究了水稻、小麦收获指数的遗传、改良方式和在育种中的应用[6, 7, 8, 9, 10], 但大部分学者仍集中在对作物产量的研究上。
甘蓝型油菜(Brassica napus)是高油分的油料作物和高蛋白的饲料作物, 也是潜在的能源作物。培育高产优质新品种一直是油菜育种的首要目标。与其他作物相比, 油菜的生物产量很高, 但收获指数显著偏低[5], 因此提高收获指数是实现油菜高产的有效途径。到目前为止, 关于油菜收获指数的系统研究较少。黄露等[11]对四川29个油菜杂交种的收获指数、生物产量、经济产量、粒壳重比、粒枝重比、粒茎重比等性状的相关分析发现, 粒茎重比对收获指数影响最大, 粒枝重比次之, 粒壳重比最小, 在生物产量相当的情况下, 适当控制茎秆重有利于油菜产量的提高。袁婺洲等[12]研究发现甘蓝型油菜角果皮内的淀粉酶活性, 可溶性糖含量及32P向籽粒的运输强度等均与油菜的收获指数呈显著正相关, 而角果皮内的淀粉含量则与收获指数呈负相关, 油菜粒壳比, 角杆比及千粒重与收获指数呈正相关, 杆重与收获指数呈负相关[13]。Luo等[14]利用155份材料的SNPs芯片数据, 对油菜株高、分枝数、生物产量、籽粒产量和收获指数进行了全基因组关联分析, 在C亚基因组上找到9个与收获指数紧密关联的SNPs标记, 并发现株高与收获指数呈显著负相关。
云南临沧独特的生态环境有利于油菜高产, 主推品种云油杂2号大面积单产可达4500 kg hm-2以上, 远高于长江流域油菜主产区。经前期调查, 云南临沧高产油菜的产量构成特点主要为有效分枝、单株角果和每角粒数多, 特别是二次有效分枝和二次分枝角果多、籽粒多且饱满。因此, 研究相同油菜品种(系)在高产环境(云南临沧)和一般环境(重庆北碚)下各产量组成性状差异和不同环境下收获指数与各产量组成性状的相关性, 可解析收获指数及构成因素受基因型和环境影响的规律与原因, 为油菜高收获指数育种提供理论依据。
从国内外广泛收集各种资源材料600多份, 通过预备试验, 选择在云南和重庆均能正常生长和成熟的321份甘蓝型油菜品种, 于2013年9月至2014年5月分别在重庆市油菜工程技术研究中心歇马实验种植基地(29º 45'39.99" N, 106º 22'38.47" E, 海拔238.57 m)和云南省临沧市农业技术推广站临翔区博尚镇勐准村实验种植基地(23º 43'56.69" N, 100º 02'4.79" E, 海拔1819.50 m), 采用育苗移栽方式, 参照卢坤等[15]的方法按完全随机区组设计三重复同时种植, 每小区2行, 每行15株, 行距40 cm, 株距20 cm。田间管理同常规生产, 确保同一地区所有样本的生长环境一致。
1.2.1 产量组成性状 成熟期从每小区选择生长正常、无病害、无机械损伤的10个样株, 分别调查每株生物学产量(biomass per plant, PB)、主序角果数(number of effective siliques on the main inflorescence, MIS)、一次有效分枝角果数(number of effective siliques on the primary branch, PBS, 简称一次角果数)和二次有效分枝角果数(number of effective siliques on the secondary branch, SBS, 简称二次角果数)。然后按主序、一次有效分枝、二次有效分枝分别装袋, 自然风干后考察主序粒重(seed weight on the main inflorescence, SWMI)、一次有效分枝粒重(seed weight on the primary branch, SWPB, 简称一次粒重)、二次有效分枝粒重(seed weight on the secondary branch, SWSB, 简称二次粒重)和3个部位的千粒重, 分别标识为TSWMI (thousand-seed weight on the main inflorescence)、TSWPB (thousand- seed weight on the primary branch)、TSWSB (thousand-seed weight on the secondary branch); 单株产量(seed yield per plant, PY)为主序粒重、一次粒重和二次粒重之和。最后, 分别计算主序每角粒数(number of seeds per silique on the main inflorescence, SSMI)、一次有效分枝每角粒数(number of seeds per silique on the primary branch, SSPB, 简称一次每角粒数)、二次有效分枝每角粒数(number of seeds per silique on the secondary branch, SSSB, 简称二次每角粒数), 不同部位的每角粒数计算公式为(粒重/千粒重× 1000)/角果数。根据上述性状考察结果, 计算单株角果数(silique number per plant, SP)、单株平均每角粒数(mean seed number per silique, MSP)和单株平均千粒重(mean thousand-seed weight per plant, MTSWP)。
1.2.2 产量和产油量收获指数 成熟期从每小区另外收获5株正常植株, 将所有分枝齐茎秆处剪断装袋, 晒干后称重作为上部干重; 再将所有角果脱粒, 种子晒干后获得单株经济产量; 使用NIRSystems.5000近红外分析仪测定各材料的含油量(oil content, OC); 将5株茎杆齐子叶节切断, 晒干后称重得到单株平均茎杆干重; 将上部干重与茎杆干重相加得到全株生物产量。按下述公式计算各材料的产量收获指数和产油量收获指数。产量收获指数(yield harvest index, YHI) = 经济产量/生物产量 = 5株种子总重/(5株上部干重+5株茎秆干重); 产油量收获指数(oil production harvest index, OHI) = 产油量/生物产量=含油量× 产量/生物产量=含油量× 产量收获指数。
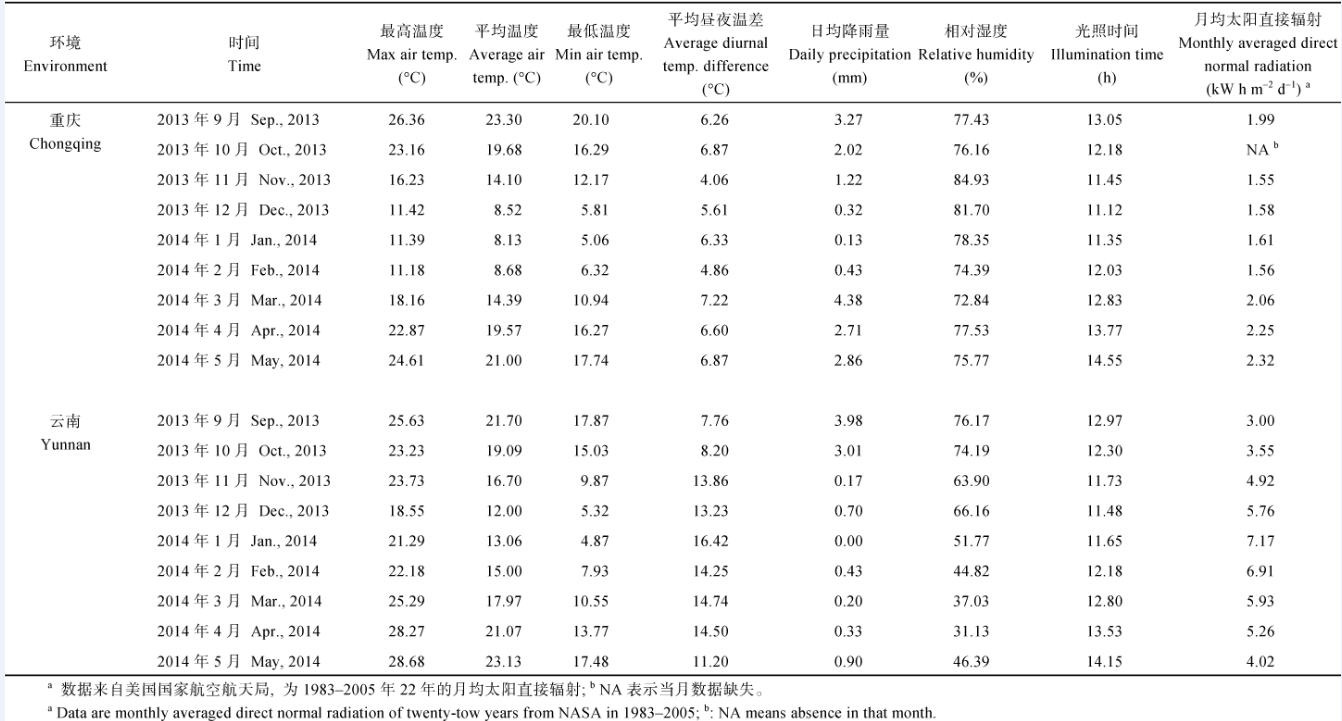
材料种植期间两地的温度、降雨、湿度、风速和光照时长数据来自美国国家海洋与大气管理局(NOAA)地球系统研究实验室物理科学部(http:// www.esrl.noaa.gov/psd/), 并以ncBrowse工具从NetCDF文件中获取相关气象数据。太阳辐射数据来自美国国家航空航天局(NASA)大气科学数据中心(https://eosweb.larc.nasa.gov/), 为两地22年(1983— 2005年)的月平均数。
利用Microsoft Excel 2010进行性状数据描述性统计分析和频率分布图制作。采用DPS7.05统计分析软件对两地各性状进行方差分析和Duncan’ s新复极差检验, 将方差组分分解为区组、环境、基因型、环境和基因型互作。用DPS7.05统计分析软件进行两地17个油菜产量组成性状与产量收获指数和产油量收获指数的相关分析和通径分析。
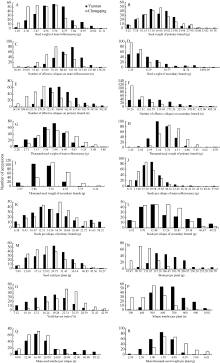
总体来看, 两地各性状数据分布情况基本相同(图1); 除二次有效分枝的粒重和有效角果数为偏态分布外, 其余性状均为正态分布, 说明这些性状均为受多基因控制的数量性状。
表1表明, 321份材料在重庆的YHI、一次粒重、主序千粒重、一次千粒重和二次千粒重均值均高于云南, 其余性状则云南高于重庆, 尤其是单株生物产量、二次分枝角果数、每角粒数和千粒重显著高于重庆, 表明云南高产环境下, 光合产物充沛、生物产量高、角果多、每角粒数多, 特别是有效的二次角果数多使产量结构更均衡, 更容易实现高产。重庆环境下YHI均值比云南高5.29%, 达到极显著差异, 主要原因是单株生物产量显著低于云南, 但相同材料在云南的含油量偏高, 所以云南环境的OHI仍然较高。
YHI和每角粒数(主序、一次和二次)在云南环境的变幅和变异系数远高于重庆, 说明在高产环境诱导下, 控制这几个性状的基因群更能表现出遗传差异。单株生物产量在重庆的变幅和变异系数远高于云南, 说明在重庆光照不足、昼夜温差小的生态环境下, 各基因型的光合效能差异能够得到更充分的体现, 有利于选择鉴定高光效品种材料。从性状稳定性看, 在云南和重庆两地, 粒重(含主序、一次和二次)和二次角果数、二次每角粒数的变异系数都较大, 说明这几个性状有较大的改良空间。
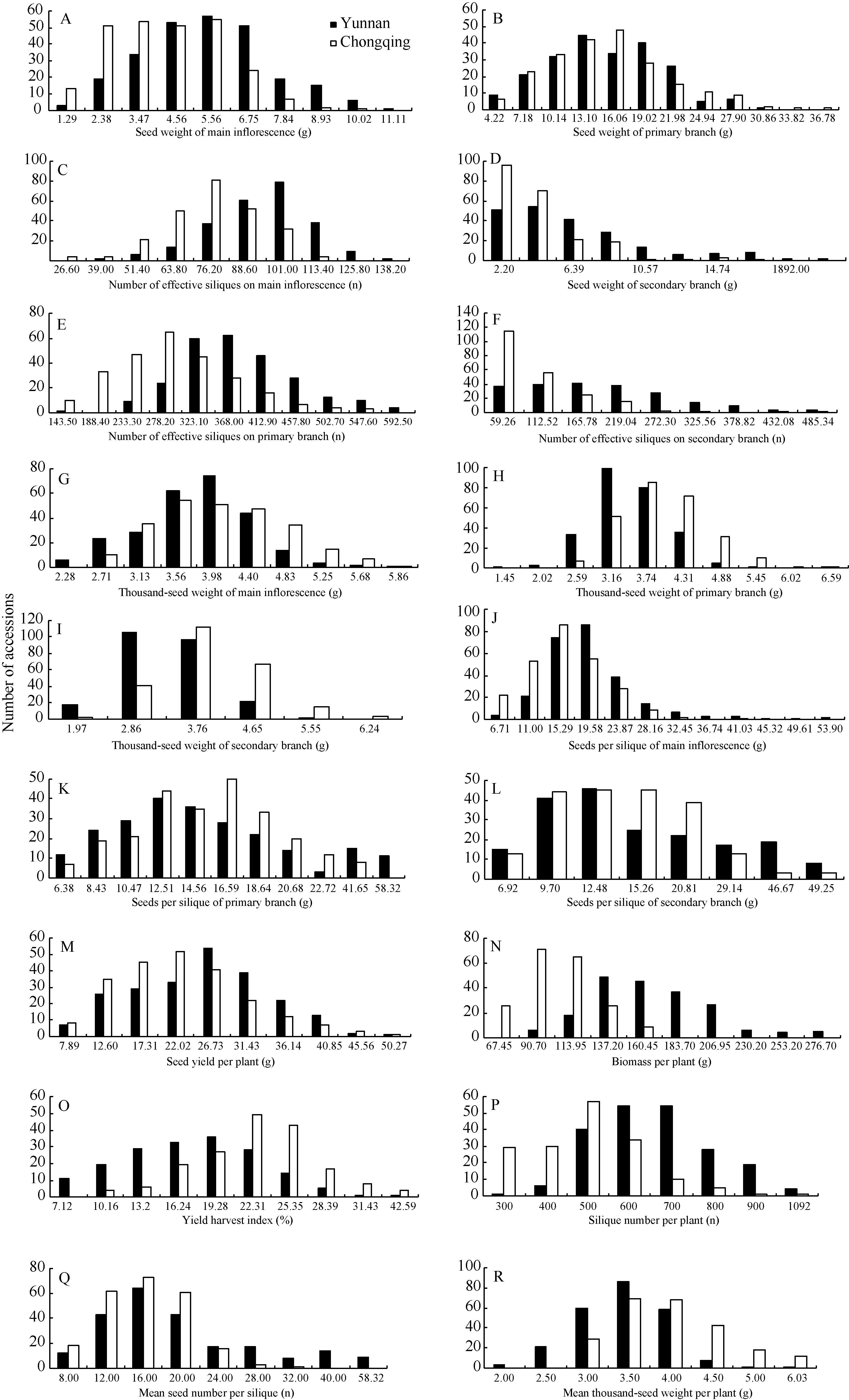 | 图1 云南和重庆321份甘蓝型油菜材料18个性状的频率分布直方图Fig. 1 Frequency distribution of 18 traits in 321 B. napus accessions cultivated at Yunnan and Chongqing |
表2表明, 除OHI、一次粒重和单株平均千粒重外, 其余17个性状在两地间均有极显著差异; 除YHI的环境与基因型互作不显著外, 其他性状的环境与基因型互作均极显著, 说明有必要对两地数据分别分析。
| 表1 重庆和云南321份材料20个性状的统计分析 Table 1 Statistical analysis of 20 traits in 321 B. napus accessions cultivated at Yunnan and Chongqing |
| 表2 云南和重庆321份甘蓝型油菜20个性状的方差分析 Table 2 Analysis of variance of 20 traits in 321B. napusaccessions cultivated at Chongqing and Yunnan |
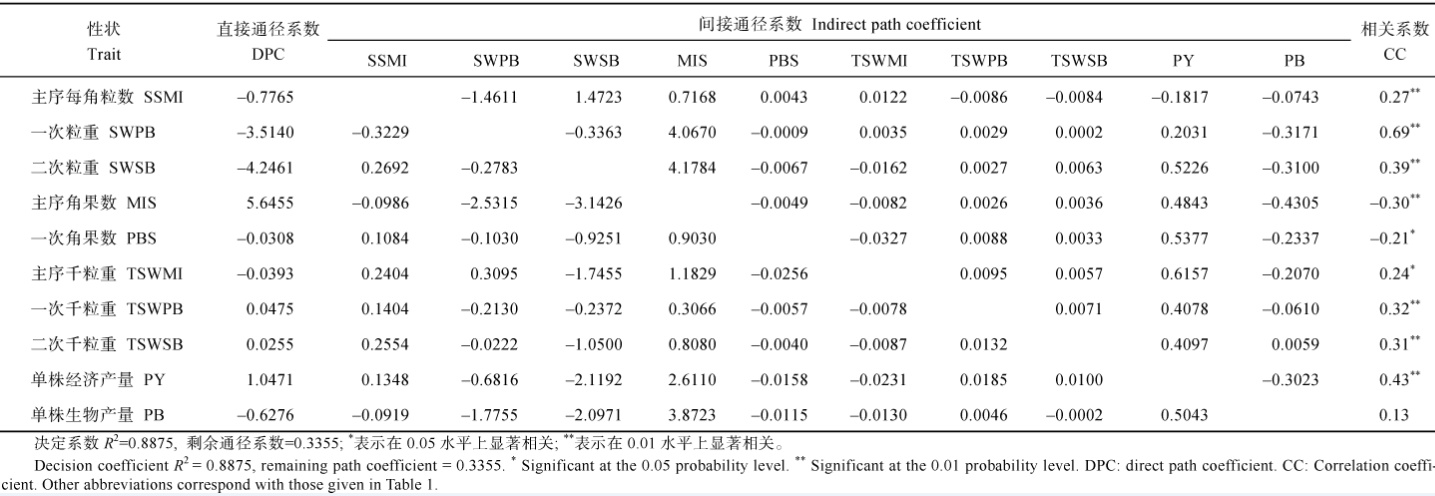
2.2.1 相关分析 表3表明, 在云南环境下, YHI与一次粒重、单株经济产量、单株平均每角粒数、一次每角粒数、二次粒重、单株平均千粒重、主序每角粒数、主序千粒重和二次千粒重呈极显著正相关, 且相关性依次递减; 与单株生物产量和主序角果数呈极显著负相关, 相关性递减。在重庆环境下, YHI与一次粒重、主序粒重、一次每角粒数、主序每角粒数、一次角果数、二次粒重、单株角果数和单株经济产量呈极显著正相关, 相关性依次递减; 与二次角果数和主序角果数呈显著正相关, 相关性递减; 与单株生物产量呈显著负相关。
比较两地YHI与各性状的相关性可知, 一次和二次角果数与云南YHI无显著相关性, 但与重庆YHI呈显著或极显著正相关; 主序角果数与云南YHI呈极显著负相关, 但与重庆YHI呈显著正相关; 各部位千粒重与云南YHI呈显著或极显著正相关, 但与重庆YHI无显著相关性。角果数、每角粒数和千粒重之间有代偿互补关系, 由上述数据可以看出云南和重庆收获指数的构成特点差异: 在云南高产环境下, 分枝角果数是可代偿性状, 而各部位千粒重为必需的正效应性状, 在重庆环境下却刚好相反。
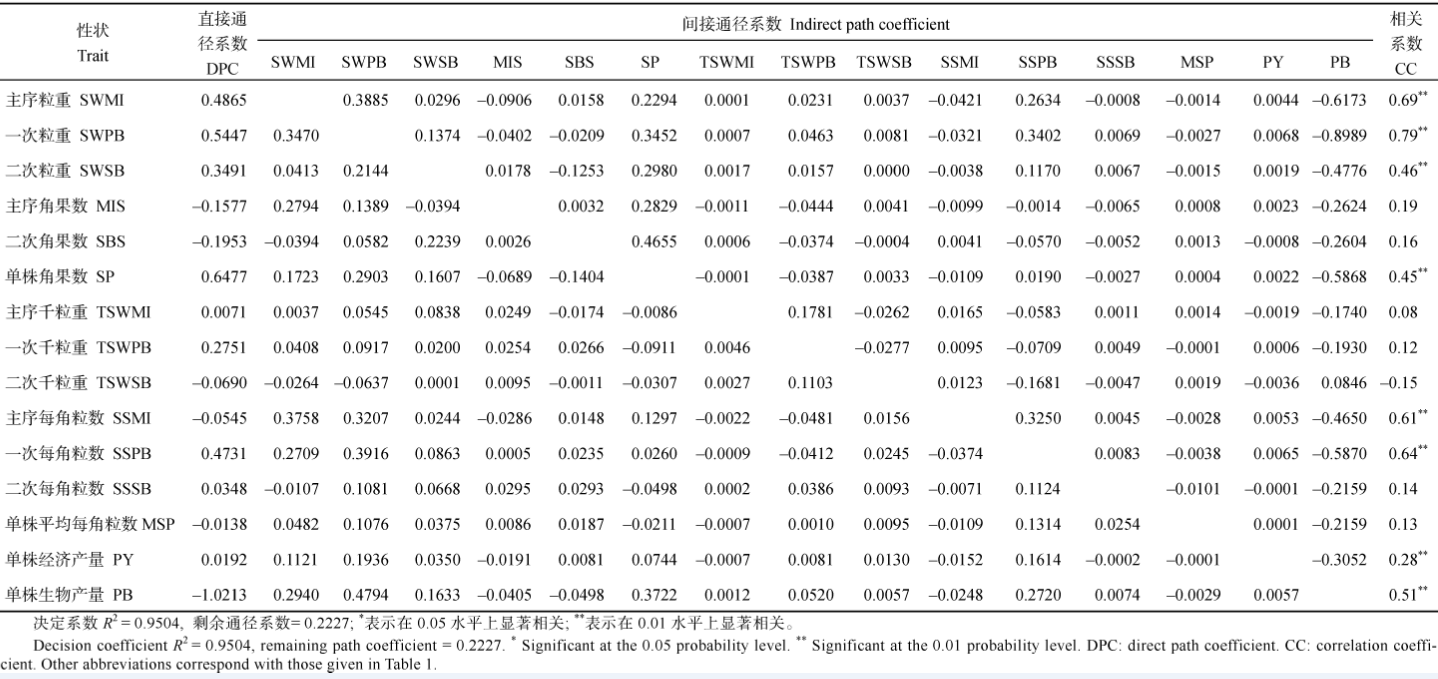
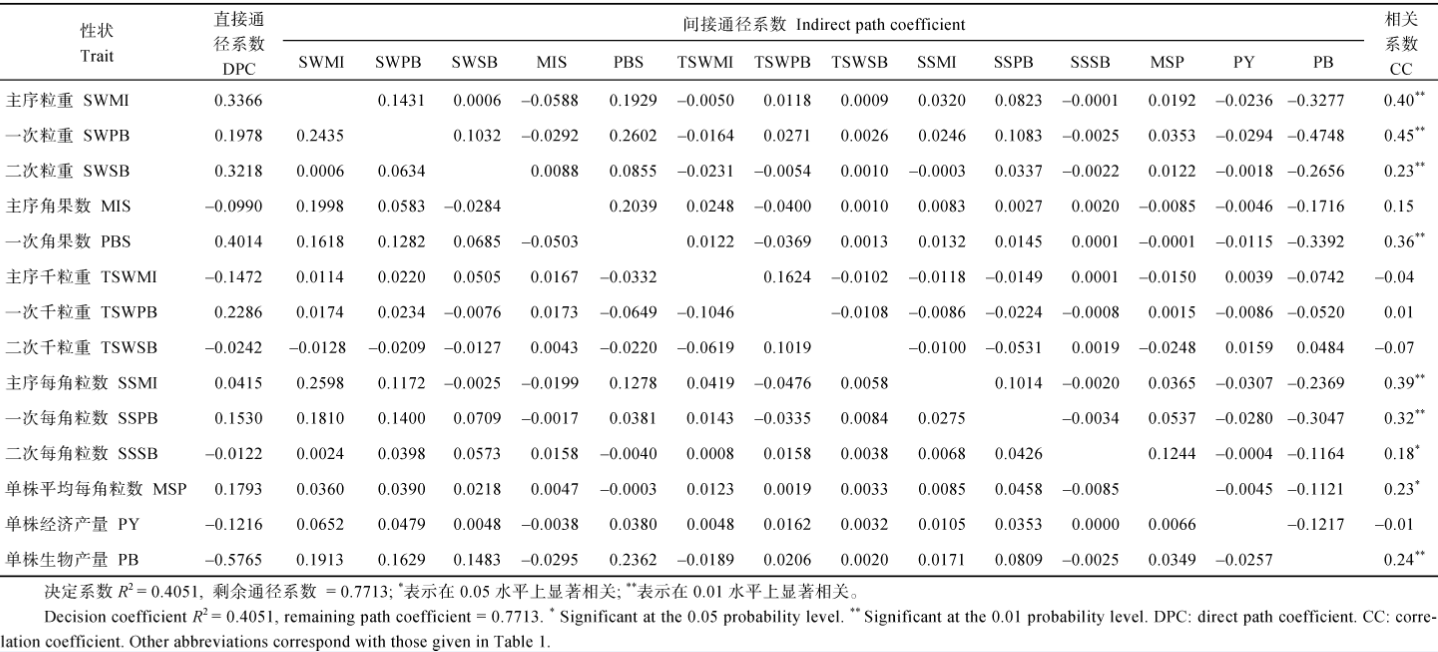
2.2.2 YHI与各农艺性状的通径分析 由表4看出: (1)主序角果数和单株经济产量对YHI的直接正效应很大, 一次和二次粒重、主序每角粒数和生物产量对YHI有较大的直接负效应, 其他性状对YHI的直接效应很小; (2)主序和二次千粒重均通过主序角果数对YHI有较大的间接正效应, 而通过二次粒重对YHI有较大的间接负效应; (3)由于各产量性状也是生物产量的一部分, 它们一般会通过生物产量形成对YHI的负效应, 特别是主序角果数、一次粒重、二次粒重、主序角果数和二次分枝角果数等性状的间接负效应更突出。上述情况说明, 在云南环境下, 各产量组成性状的改良均能通过提高主序角果数和单株经济产量而提高YHI, 各性状之间相互抑制抵消的现象不明显, 但对生物产量要适当控制。重庆环境的通径分析(表5)表明, 各产量组成性状对YHI的贡献与云南不同: (1)各产量组成性状中, 单株角果数、主序、一次分枝和二次分枝的粒重和一次分枝每角粒数均对YHI有很大的直接效应, 单株经济产量对YHI的直接效应不明显, 但单株生物产量对YHI有很大的直接负效应; (2)各产量组成性状对YHI的间接效应都很低, 但由于它们提高了生物产量而对YHI有较大的负效应; (3)主序粒重、一次分枝粒重、单株角果数和一次分枝每角粒数这4个性状有正向连带功效, 它们相互间均能对YHI产生较大的间接正效应; (4)主序每角粒数直接效应不明显, 但通过主序粒重、一次分枝粒重和一次分枝每角粒数对YHI有较大的正向间接效应, 最终与YHI极显著正相关; (5)其他各产量组成性状之间相互间接作用对YHI的效应都很小。上述情况说明, 重庆环境下油菜YHI构成更复杂, 单纯提高单株经济产量不能直接提高YHI, 单纯改善某一个产量组成性状也不能明显提高单株经济产量, 进而提高YHI; 高YHI必须将主序和一次分枝的产量组分有机结合, 且严格控制生物产量才能实现。
| 表3 云南和重庆321份甘蓝型油菜产量收获指数和产油量收获指数与17个产量组成性状的相关系数 Table 3 Correlation coefficients between YHI or OHI and 17 yield component traits in 321B. napusaccessions cultivated at Chongqing and Yunnan |
2.3.1 OHI与各产量性状的相关分析 从表3可以看出, 与YHI相比, OHI与各产量组成性状的相关性有所降低, 说明含油量的引入使收获指数与产量相关性状的关系变得更为复杂。在云南环境下, YHI和OHI与各产量组成性状的相关性方向和显著程度基本一致, 只有主序粒重和二次分枝每角粒数两个性状与YHI的关系为显著或极显著正相关, 而与OHI的相关性却不显著; 值得注意的是生物产量与OHI的相关性不显著, 说明在高产环境下, 光合产物与产油量的关系比较复杂。在重庆环境下, 除主序角果数、二次角果数、单株平均每角粒数和单株经济产量外, 其他产量组成性状与YHI和OHI的相关性方向和显著程度基本一致; 比较意外的是单株经济产量与OHI没有相关性, 说明含油量性状的导入抵消了单株经济产量与YHI的关系; 单株生物产量与OHI呈极显著正相关, 说明在光合产物普遍不足的重庆, 生物产量的提高有利于产油量的提高。
两地数据比较分析表明, 单株平均千粒重、主序粒重、单株角果数、主序角果数、一次分枝角果数、3个部位的千粒重、单株经济产量和生物产量在两地与OHI的相关性均不相同, 说明不同环境下, OHI的构成方式不同, 有必要分别进行通径分析。
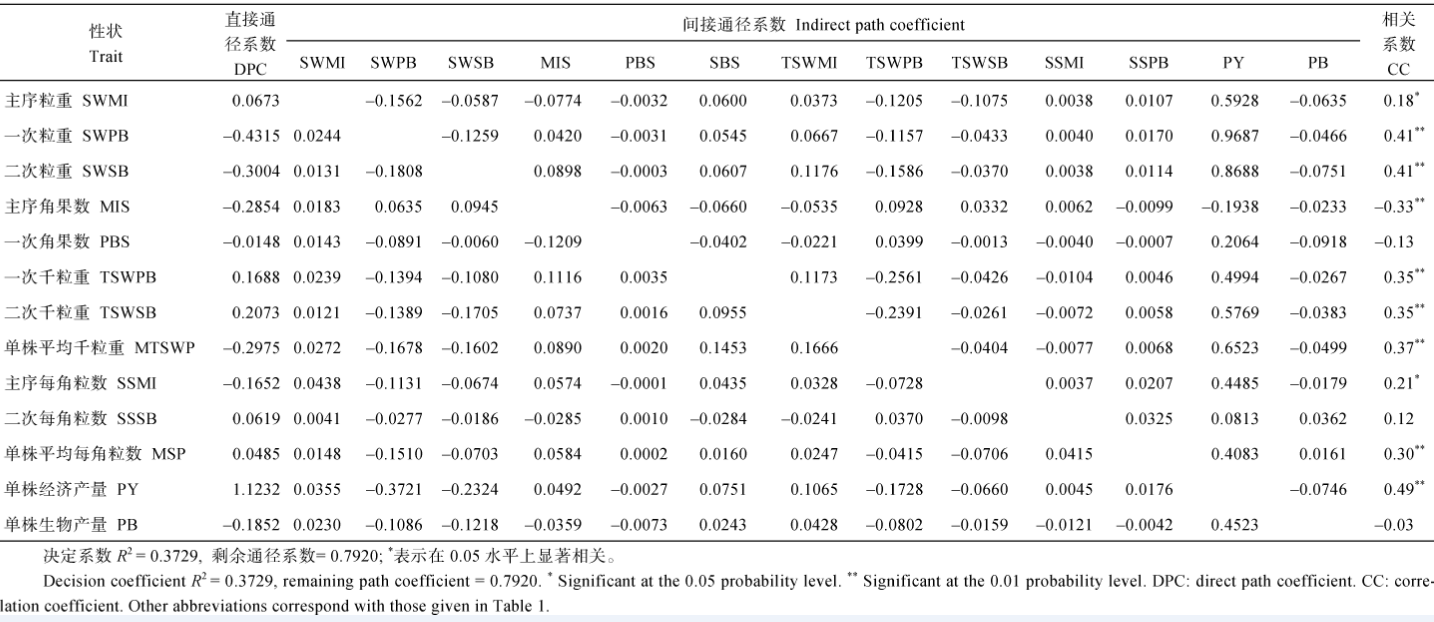
2.3.2 OHI与各产量组成性状的通径分析 云南环境下的通径分析(表6)表明: (1)单株经济产量的直接通径系数最大, 达到1.1232, 且与OHI呈极显著正相关, 说明单株经济产量对OHI起直接正向作用; (2)一次和二次粒重与OHI呈极显著正相关, 但其直接效应为负值, 主要通过提高单株产量实现对OHI的提高; (3)主序角果数与OHI的极显著负相关主要由直接效应来实现, 从表1可以看出, 云南环境下各材料的主序发达, 变异幅度较大, 适当减少主序角果数有助于提高OHI; (4) 3个部位的千粒重和每角粒数均与OHI呈显著或极显著正相关, 但其直接作用不大, 主要通过提高单株产量来实现。重庆环境下的通径分析(表7)表明: (1) 3个部位的粒重对OHI均有较大的正效应, 它们还通过一次分枝角果数对OHI有较大的间接正效应, 但它们通过提高生物产量又对OHI有较大的负效应, 正负效应抵消后, 仍与OHI保持极显著正相关; (2)一次分枝角果数和每角粒数对OHI均有较大的正效应, 它们通过增加各部位的粒重对OHI有一定的间接正效应, 但它们又通过增加生物产量对OHI有较大的间接负效应, 正负效应抵消后, 仍与OHI保持极显著正相关; (3)单株经济产量对OHI有直接负效应, 同时还通过增加生物产量对OHI有间接负效应, 但它通过大多数产量组成性状对OHI有一定正效应, 正负效应抵消后, 与OHI的相关性不明显; (4)单株生物产量对OHI有较大的直接负效应, 但它通过增加各部位粒重和一次角果数对OHI产生较大的间接正效应, 最后表现与OHI的极显著正相关。
| 表4 云南321份甘蓝型油菜17个产量组成性状与产量收获指数的通径分析 Table 4 Path analysis between YHI and 17 yield compoment traits in 321 B.napus accessions cultivated at Yunan |
| 表5 重庆321份甘蓝型油菜17个产量组成性状与产量收获指数的通径分析 Table 5 Path analysis between YHI and 17 yield component traits in 321 B.napus accessions cultivated at ChongQing |
| 表6 云南321份甘蓝型油菜17个产量组成性状与产量收获指数的通径分析 Table 6 Path analysis between YHI and 17 yield component traits in 321 B.napus accessions cultivated at Yunnan |
比较云南和重庆两地各性状与OHI的通径分析结果, 最大的差别是单株经济产量对OHI的直接效应值, 云南的直接正效应非常大, 其他产量性状基本上都是通过提高单株经济产量来提高OHI, 而重庆的直接作用不明显, 与OHI的相关性也不明显。重庆环境下, 粒重、一次分枝角果数和每角粒数几个性状相互协同对OHI产生正效应; 两地单株生物产量对OHI均为负效应, 但在云南环境下直接、间接效应都不强, 相关性不明显, 重庆环境下直接负效应较大, 但又可通过提高多个产量组成性状而与OHI呈极显著正相关。上述情况表明, 在云南选育高OHI材料可直接选单株经济产量较高的材料, 在重庆却要同时考虑各部位粒重及一次分枝角果数和每角粒数, 此外还要选择光合效率较高、生物产量较高的材料。
云南各性状关于OHI通径分析的决定系数仅0.3729, 重庆该决定系数也仅为0.4051, 远低于两地各性状关于YHI的决定系数, 说明本研究中的产量组成性状仅是影响产油量收获指数的部分性状。
相同材料在不同环境中所表现性状变异反映了该性状基因群对环境变化的响应差异, 而不同性状差值的相关性, 反映不同性状基因群对环境的协同响应。为了研究油菜产量相关性状与YHI和OHI对环境的协同响应情况, 我们计算了参试材料YHI和OHI两地差值与各产量相关性状差值间的相关性(表8)。
YHI两地差值与三部位粒重、二次角果数、一次分枝千粒重、主序和一次分枝每角粒数、单株平均每角粒数和单株经济产量的差值呈显著正相关, 说明在环境诱导下, 各油菜材料YHI的变异与大多数产量组成性状的变异同步, 即控制各产量组成性状的基因群与YHI的基因群对环境的响应有协同趋势, 要选择环境钝感、YHI高且稳定的亲本材料, 需要考察上述各产量组成性状的遗传稳定性。
OHI两地差值与三部位粒重、主序和一次分枝千粒重、3个部位每角粒数等性状的差值呈显著或极显著正相关, 与主序角果数差值呈显著负相关, 说明控制各产量组成性状的基因群与OHI的基因群对环境的响应也有协同趋势, 但主序角果数基因群对环境的响应效应相反, 联系到主序角果数和一次分枝角果数差值与YHI差值关系不密切的现象, 可以确定, 选择在云南角果数多、在重庆角果数较少的材料, 其OHI的稳定性较好。
| 表7 重庆321份甘蓝型油菜17个产量组成性状与产量收获指数的通径分析 Table 7 Path analysis between OHI and 17 yield component traits in 321 B.napus accessions cultivated at ChongQing |
| 表8 云南和重庆YHI或OHI差值与17个产量组成性状差值的相关分析 Table 8 Correlation coefficients between difference of YHI or OHI and difference of 17 yield component traits of Chongqing and Yunnan |
| 表9 材料种植期间云南和重庆的气象资料 Table 9 Meteorological data at Chongqing and Yunnan during cultivation |
提高油菜收获指数是进一步提高油菜单产的主要途径。本文初步揭示了两种环境下, 油菜产量构成特点、油菜产量和产油量高收获指数与各产量组成性状的关系、以及两地差值的协同变化规律。
两地各性状均值和变异幅度分析表明, 尽管321份参试材料的经济产量在云南比重庆高13.99%, 但收获指数却比重庆低5.29%。Badger[16]研究表明, 在寡日照条件下, 光能转化效率主要受限于CO2固定的初级量子产率, 如较低的核酮糖-1, 5-二磷酸羧化酶/加氧酶(ribulose bisphosphate carboxylase oxygenase, Rubisco)活性; 而在光照充足区域, 固定CO2的叶绿体基质反应效率是光能转化效率的主要限制因子。与寡日照相比, 充足的光照能使大豆和水稻的CO2同化率均提高40%, 增加C3草本植物地上部生物产量约20%, 最终实现增产12%~23% [17]。从气象资料比较分析可以发现(表9), 材料种植期间云南临沧高产环境的昼夜温差、日均温度和太阳直接辐射量均显著高于重庆环境(P < 0.01), 使云南临沧各材料平均生物产量比重庆高66.23%, 高于其他作物在光照充足条件下的生物产量增加率。因此, 除光照强度外, 昼夜温差及日均温度是否对油菜生物产量有较大影响, 值得进一步研究。
在光照充足、温度适宜的最适生长条件下, 不同材料的收获指数的差异可能主要与光合产物转运能力有关。在未经过定向选择前, 多数材料并不能充分利用光合产物, 各基因型的光合转运能力差异得到充分体现, 因此, 云南环境下产量收获指数的变异系数比重庆高51.81%, 有利于对高转运效率基因型的鉴定; 而在重庆这种光照不足、昼夜温差小的生态环境下, 各基因型的光合效能差异更能够得到充分体现, 有利于选择高光效基因型。
单株角果数是油菜重要的产量性状, 但从收获指数的角度考虑, 角果数并不是越多越好。因为, 单株角果数既是籽粒产量的重要基础, 也是生物产量的主要组成部分, 当单株角果数增加到一定程度, 每角粒数、千粒重和含油量均会相应下降, 超过补偿点, 会导致YHI和OHI下降。前人研究表明, 水稻和小麦的单株穗数及油菜的单株角果数均与YHI无显著相关性[18, 19, 20, 21]。本研究将单株角果分为主序、一次和二次分枝3个部位分别与YHI进行相关分析, 结果发现重庆寡日照区域3个部位的角果数均与YHI呈极显著正相关, 而光照充足的云南临沧区域仅有主序角果数与YHI呈极显著负相关, 提示在光热资源不同的地区, 不同部位的角果数与YHI的相关性可能存在差异, 这对培育油菜高YHI品种及其他作物的类似分析均有一定的借鉴意义。
千粒重是重要的产量性状, 也是“ 库” 容量的主要指标。前人研究表明, 水稻千粒重对YHI的影响不大[18], 但在油菜中, 千粒重与YHI呈极显著正相关[20]。本研究中, 云南环境3个部位的千粒重均与
YHI呈极显著或显著正相关, 与前人研究结果一致。但重庆环境3个部位的千粒重均与YHI无相关性, 表明千粒重与YHI的关系比较复杂, 不同作物和不同环境均能影响千粒重与YHI的关系。各部位千粒重在重庆寡日照环境下高于云南光照充足的环境下, 表明油菜在“ 源” 不足的条件下, 很可能会优先保障部分籽粒充实, 以保证部分种子的质量; 而在云南环境下, 各部位千粒重与YHI和OHI均呈极显著正相关, 说明在光照充足条件下, 千粒重与光合产物转运能力密切相关, 高收获指数材料有较大的籽粒。
每角粒数也是重要的产量性状, 与YHI呈显著正相关[13]。本研究中, 云南环境下各部位每角粒数都极显著高于重庆环境下, 且在两种环境下, 主序和一次分枝每角粒数都与YHI和OHI有极显著正相关, 结果与前人在油菜中的分析一致, 但与水稻的结果存在差异, 表明每角粒数与YHI的关系较为复杂, 需要针对目标作物具体分析。同时, 也说明每角粒数性状需要充足的“ 源” 和畅通的“ 流” 来保证。两种环境下二次分枝的每角粒数有一定差别, 在云南环境下, 它与YHI有显著正相关, 在重庆环境下没有相关, 说明二次分枝每角粒数在云南产量中的重要性。在重庆环境下, 大多数二次分枝的每角粒数非常少, 在单株产量中所占份额少, 对产量收获指数的影响小。
角果数、每角粒数和千粒重之间有代偿互补关系。本试验在云南高产环境下, 分枝角果数是可代偿性状, 而各部位千粒重为必需的正效应性状, 在重庆环境下却刚好相反。
云南高产环境下, 不同部位单株角果数和每角粒数均显著高于在重庆环境下, 但主序和一次分枝千粒重显著低于在重庆环境下。在光照充足的环境下, 各产量组成性状的改良均能通过提高主序角果数和单株经济产量而提高YHI, 但要适当控制生物产量; 而寡日照环境下YHI构成复杂, 必须将主序和一次分枝的产量组分有机结合, 且严格控制生物产量才能提高YHI。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|