


{kind=link}
{kind=link}
{kind=link}
{kind=link}
小麦茎秆实心度对茎秆强度的影响及相关性状QTL分析
[潘婷1, *  , 胡文静
, 胡文静2, * , 李东升2 , 程晓明2 , 吴荣林2 , 程顺和1, 2, * ]
, 胡文静]
|
|


第一作者联系方式: E-mail:862430412@qq.com, Tel: 18550707316
在QTL水平上研究茎秆实心度与强度的遗传关系及实心度对茎秆强度的影响, 为小麦抗倒伏育种提供依据。利用普通小麦宁麦18 与实秆小麦种质“武云实秆”的F2群体和F2:3家系, 对小麦茎秆强度、实心度及影响实心度的相关性状包括厚径比、壁厚、茎粗和髓腔直径进行了相关分析, 并对茎秆强度相关性状QTL进行分子标记定位及遗传效应分析。结果表明小麦茎秆强度与厚径比、壁厚均呈极显著正相关, 与髓腔直径呈极显著负相关。基于复合区间作图法进行QTL定位, 检测到与茎秆强度、厚径比、壁厚、茎粗和髓腔直径相关QTL共23个, 分布在1B、3B、4A、4B、5A上, 表型贡献率3.5%~44.0%。在染色体3B、4A和5A上的标记区间 gwm547-gwm247、 wmc718-wmc468和 gwm156-gwm443均检测到贡献率很高的茎秆实心度相关QTL, 说明在这3条染色体上可能存在控制茎秆强度的主效QTL。用普通小麦宁麦13 (N13)×武云实秆的24个F7家系检验分子标记 gwm247的可靠性表明利用标记 gwm247选育茎秆实心度优于宁麦13的概率较大。研究结果为进一步精细定位相关主效QTL以及分子标记辅助改良小麦茎秆强度奠定了基础。
Improving stem strength is an important way to increase lodging resistance of wheat. Solid stem traits, including thickness-diameter ratio of culm, stem diameter, culm wall thickness and pith diameter, are the main targets for improvement of stem strength in wheat. In the present study, an F2 population and F2:3 families derived from the cross N18 × WYSG were used to study the influence of stem solidness on stem strength and to identify QTLs for solidness related traits. The thickness-diameter ratio of culm, stem diameter, culm wall thickness and pith diameter were significantly correlated with stem strength. By using composite interval mapping (CIM), a total of 23 QTLs for the five traits were detected on chromosomes 1B, 3B, 4A, 4B, and 5A. Phenotypic variations explained by individual QTL ranged from 3.5% to 44.0%. Pleiotropic QTLs were found on chromosomes 3B ( gwm547-gwm247), 4A ( wmc718-wmc468), and 5A ( gwm156-gwm443), all contributing very high phenotypic variations. The reliability of the marker gwm247 was confirmed using 24 F7 lines derived from the N13 (hollow stem) × WYSG (solid stem) cross. The markers linked to major QTLs could be valuable in wheat breeding for lodging resistance with marker assisted selection technology.
随着小麦产量不断提高同时伴随施肥量的增加, 倒伏成为威胁我国小麦高产稳产的重要因素之一[1, 2]。植株矮化可以有效缓解小麦倒伏危害的发生[3, 4], 但一般会导致生物量减少, 群体郁闭, 通风透光差, 病害加重, 影响产量[5], 并且植株矮化后茎秆强度可能减弱, 仍然会发生倒伏。因此, 改良茎秆强度提高抗倒性是解决小麦倒伏的理想途径[6]。
目前, 抗倒性相关研究, 包括茎秆形态结构[7, 8, 9, 10, 11]、解剖结构[12, 13]、化学成分[7]等对抗倒性的影响。汪灿等[12]关于荞麦茎秆解剖结构及木质素代谢与抗倒性关系的研究指出茎秆中木质素含量、机械组织厚度、茎壁厚度、大维管束数目和维管束面积可以作为禾本科茎秆抗倒伏育种的重要指标。有学者认为小麦茎秆强度与中下部茎粗及鲜重呈显著正相关, 而与各节间长度呈显著负相关[14, 15, 16]。但是也有一些学者认为茎秆粗度与秆强之间没有显著相关性[17]。实秆小麦在抗倒伏育种中起很重要的作用。陈华华等[18]和Kong等[19]研究发现, 实秆小麦较普通小麦具有极小的髓腔, 腔中有髓, 其机械组织含量显著高于普通小麦, 因而实秆小麦茎秆强度较高。
在20世纪40年代, 为了预防麦茎蜂(Cephus cinctus Norton)的危害, 加拿大推广了实秆小麦Rescue和Chinook[20, 21]。McNeal等[22]和Wellace等[23]提出实秆性状可能对小麦产量造成影响。Adrianna等[24]认为实秆麦是到目前为止预防麦茎蜂危害最有效的方法, 其实验表明实秆基因不会影响籽粒品质, 但对产量有影响, 当麦茎蜂数量很少时, 实秆麦的产量显著低于空秆麦, 而当麦茎蜂数量大到足以影响小麦产量时, 实秆麦的产量与空秆麦之间没有显著差别。但是也有实验表明, 实秆基因与产量性状之间没有显著关系[17]。McNeal等[25]以6个实秆小麦与3个普通小麦杂交得到的17个系为材料, 证明实秆基因并不是影响小麦高产的因素。
国内、外学者利用分子标记技术对小麦茎秆强度相关性状进行了基因定位。Hai等[14]利用CA9613 × H1488创建的DH群体对控制茎秆强度、茎壁厚度、髓腔直径、茎粗进行基因定位, 其中控制茎秆强度的基因位于3A和3B上, 并分别解释10.6%和16.6%的表型变异。郭会君等[26]研究发现控制小麦茎秆强度的基因位于2D和5B上, 共同解释24%的表型变异。Verm等[27]利用Milan × Catbird产生的DH群体将控制茎秆抗折力的基因定位在4B和2D上, 表型贡献率分别为11%和13%。
国内研究茎秆强度与茎壁厚、茎粗、节间长度等关系较多, 较少研究实秆小麦与茎秆强度的关系。本实验旨在研究小麦茎秆实心度对茎秆强度的影响, 并定位相关基因进行, 为培育小麦抗倒伏新品种奠定基础。
武云实秆是四川省农业科学院采用实秆小麦86-741[28]与川麦42 [29]杂交、回交育成, 其茎秆基部有极小髓腔, 髓腔中有髓。以空心高秆种质“ 宁麦18” 为母本, 矮秆实心种质“ 武云实秆” 为父本。于江苏里下河地区农业科学研究所万福基地试验田, 2013— 2014年生长季获得F2群体, 2014— 2015年生长季获得的杂交F1代和F2:3家系。验证群体是宁麦13和武云实秆的杂交F7代高代系。
2014年和2015年, 分别选取成熟期小麦武云实秆、宁麦18和F1植株各20株, 取F2群体184个单株及对应F2:3家系每行20株。用手术刀切开基部第1节间和第2节间, 用游标卡尺测量外茎及内径的长度, 每个单株成“ 米” 字形测3次取均值。外径记为茎粗, 内径记为髓腔直径, 秆壁厚度为外径与内径差值的一半。实心度用厚径比来表示, 厚径比为秆壁厚度与茎粗的比值[24, 30]。
2015年度, 分别选取成熟期小麦武云实秆、宁麦18及F1植株各20株, 取对应184个F2: 3家系每行20株采用茎秆强度检测仪YY1-a测力计(浙江托普仪器有限公司), 垂直作用于茎秆基部第1和第2节间的中间位置, 缓缓用力, 茎秆折断时的最大读数即为该茎的茎秆强度。
从1872对SSR引物中筛选出221对具有多态性的引物。利用JoinMap4.0构建遗传图谱, 其中170个SSR标记基本均匀分布在18条染色体上(1D、4D、7D未覆盖到), 覆盖小麦染色体组1482.15 cM, 标记间平均遗传距离为8.72 cM。利用WinQTLCart 2.5软件在全基因组范围对控制小麦茎秆强度及实心度相关性状的QTL进行检测, 采用复合区间作图法进行QTL定位[31], 当阈值(LOD)大于2.5时, 就认为可能存在一个QTL, 同时计算每个QTL的贡献率和加性效应。
武云实秆和F1植株基部第1和第2节间的秆强、厚径比、壁厚、髓腔直径均与宁18之间存在极显著差异, 武云实秆的茎秆强度和厚径比分别是宁麦18的2.20倍和2.97倍(表1)。而武云实秆基部第1节间的茎粗与宁麦18之间差异并不显著。在这些性状中, 茎秆强度的变异系数都高于10%, 基部第2节间变异系数较大, 说明茎秆强度在品种间有较高的遗传变异。
| 表1 亲本宁麦18、武云实秆及其F1代植株基部第1和第2节间茎秆强度相关性状 Table 1 Stem strength related traits for the first and second stem internodes of Ningmai 18, Wuyunshigan, and their F1 |
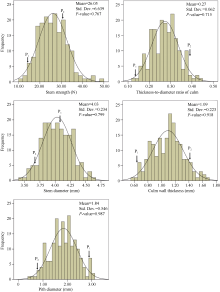
F2:3家系茎秆第2节间的强度、厚径比、茎粗、秆壁厚度和髓腔直径均呈现连续分布(图1), 表明这些性状均属多基因控制的数量性状。其中茎秆强度变幅为11.0~49.4 N, 厚径比的变幅为0.13~0.44, 茎粗的变幅为3.43~4.65 mm, 壁厚的变幅为0.58~1.69 mm, 髓腔直径的变幅为0.45~3.09 mm。这几个性状均表现超亲分离, 其中茎秆强度和茎粗表现更明显, 表明实秆的遗传基础更复杂。
 | 图1 宁麦18 (P1)与武云实秆(P2)杂交F2:3家系的表型分布Fig. 1 Phenotypic variations in the F2:3 lines derived from the cross between Ning 18 (P1) and Wuyunshigan (P2) |
F2:3家系基部第2节间茎秆强度与厚径比、壁厚呈极显著正相关(表2), 而与髓腔直径呈极显著负相关, 即小麦秆壁越厚、髓腔越小, 其实心度越高, 茎秆强度也就越大。而茎秆粗度与茎秆强度并没有显著相关性。
茎秆基部第2节间的实心度与茎壁厚度呈极显著正相关, 与茎粗和髓腔直径呈显著负相关, 即茎秆越细、茎壁越厚髓腔直径越小, 茎秆的实心度就越高。
| 表2 小麦茎秆强度与性状的相关系数 Table 2 Correlation coefficients between stem strength and culm traits in wheat |
共检测到4个控制茎秆强度的QTL (表3和图2), 位于3B、4A、4B和5A染色体上, 分别解释表型变异的20.7%、4.9%、6.9%和5.0%。其中QSs-3B、QSs-4B和QSs-5A-2增效基因来自武云实秆, 而QSs-4A增效基因来自宁麦18。
在3B、4A和5A染色体上共检测到4个控制小麦厚径比的QTL (表3和图2)。其中, QTdr-3B-2在两年中均被检测到, 且分别解释表型贡献率为28.9%和43.9%, 遗传效应很大, 增效基因均来自为武云实秆。
与茎秆粗度相关的2个QTL分别位于1B和3B染色体上, 其中QSd-1B在两年中均被检测到, 分别可解释表型贡献率的20.7%和9.1%。
在3B、4A和5A染色体上检测到4个控制小麦髓腔直径的QTL, 单个QTL可解释表型变异的4.0%~41.9%, 其中QPd-3B-2和QPd-4A加性效应为正, 增效基因均来自宁麦18。
在3B、4A、4B和5A染色体上检测到5个控制茎壁厚度的QTL, 加性效应均来自武云实杆。位于3B上的QCwt-3B-2在两年度均被检测到, 表型贡献率分别为22.3%和44.0%; 而位于4A和5A上的QTL对表型性状的贡献率分别为24.5%、10.9%和10.3%。
 | 图2 小麦茎秆强度相关性状QTL在染色体上的分布Fig. 2 Chromosome location of putative QTL associated with traits related to stem strength in wheat |
| 表3 小麦茎秆强度相关性状的QTL Table 3 Putative QTLs for traits related to stem strength |
用24个宁麦13× 武云实秆F7代高代系验证分子标记gwm247的可靠性, SSR标记gwm247在宁麦13× 武云实秆F7家系中扩增结果显示, 除了23号家系, 其余家系在此位点的基因型均与武云实秆相同(图3), 其茎秆厚径比除21号和23号家系外, 其余家系均比宁麦13显著较大(图4和表4), 表明利用标记gwm247选育出茎秆实心度优于宁麦13的茎秆类型概率较大。
 | 图3 SSR标记gwm247在宁麦13 (P3) × 武云实秆(P2) 24个F7家系中的扩增结果Fig. 3 Amplification patterns of the SSR marker gwm247 in 24 F7 lines derived from the cross of Ningmai 13 (P3)× Wuyunshigan (P2) |
 | 图4 宁麦13 (P3) × 武云实秆(P2) F7家系茎秆强度与厚径比分布图Fig. 4 Histogram of stem strength and thickness-diameter ratio of culm for 24 F7 lines derived from Ningmai 13 (P3) and Wuyunshigan (P2) |
| 表4 各家系与宁麦13茎秆强度与厚径比的差异显著性比较 Table 4 Difference in stem strength and thickness-diameter ratio of culm between each lines and Ningmai 13 |
植株高度[32, 33]、重心高度[34]、茎秆节间长度[17]、髓腔直径[18]、茎壁厚度[35]、节间干重[11]、维管束量[36]、木质素、纤维素含量[37]等都是影响植株抗倒性的重要因素。本研究发现茎秆强度与基部节间第1和第2节间实心度、秆壁厚度和髓腔直径均呈极显著相关。对于茎秆粗度与抗倒性的关系, 不同学者观点分歧。部分学者认为茎秆越粗, 植株抗倒伏能力越强[5, 11], 也有不少学者认为茎秆粗度与抗倒伏能力关系不密切[17]。而本研究表明, 小麦茎秆强度与基部第1节间茎秆粗度关系不显著, 与基部第2节间关系显著, 这说明基部第2节间粗度可作为衡量茎秆强度的一个指标。因此在小麦育种过程中选择茎秆粗、秆壁厚、髓腔小、实心度高的小麦品种有助于提高小麦茎秆强度从而提高抗倒性以达到稳产的目的。
与Hai等[14]的报道一致, 本研究发现小麦茎秆基部第2节间强度、实心度、壁厚、茎粗和髓腔直径在F2:3家系中均呈现连续分布, 表明这些性状均属多基因控制的数量性状。共检测到23个与这5个性状关联的QTL, 分布于5条染色体上, 其中在3B染色体标记区间gwm547-gwm247同时检测到控制5个性状的QTL, 解释的表型变异率为8.14%~44.00%, 说明该区域在控制小麦抗倒伏方面起着重要作用。茎秆强度基因被定位在3B、4A、4B和5A染色体上, 而位于3B上的QSs-3B-2解释表型变异率较大, 该位点与Hai等[14]将控制秆强的基因QSs-3B定位在标记gwm108-wmc291之间结果相近, 可能是控制茎秆强度的主效QTL。
茎秆实心性状受到复杂的数量性状控制, 在回交育种过程中常常丢失实心性状, 所以仅通过观察表型来改良小麦品种的茎秆实心度很困难, 分子标记辅助育种可以将分子标记应用于作物改良。早在1959年就有关于实秆基因的报道, Larson和MacDonald等[38, 39]利用S-615的单体系推断出3B和5A染色体上有控制小麦茎秆实心性状的位点。到目前为止, 关于小麦茎秆实心性状的QTL大多定位在3BL上, 并与标记gwm247、gwm547、gwm340和barc77紧密连锁[18, 19, 40]。本研究验证了前人的这些报道, 而且还发现了一些茎秆实心度相关的新位点(表3和图2), 例如在4A染色体标记wmc718- wmc468之间同时定位了控制茎秆实心度、壁厚和髓腔直径的QTL, 在一定程度上解释了茎秆壁厚、髓腔直径与实心度存在显著相关性。其中位于3B染色体的QTdr-3B-2表型贡献率很大, 该位点很可能是控制茎秆实心度的主效QTL。
本研究在分子标记gwm247附近检测到多个与抗倒性相关的QTL, 且对表型贡献率都很大, gwm247可用作分子标记辅助抗倒伏育种的重要标记。本研究用普通小麦宁麦13× 武云实秆的24个F7家系检验分子标记gwm247的可靠性表明利用标记gwm247选育出茎秆实心度优于宁麦13的概率较大(图3、图4和表4)。但要开发可靠分子标记就需要进一步定位或克隆该基因。本研究将为进一步精细定位实秆基因, 选育抗倒小麦新品种奠定基础。
检测到与茎秆强度、厚径比、壁厚、茎粗和髓腔直径相关QTL共23个, 分布在1B、3B、4A、4B、5A上, 其中在3B染色体标记区间gwm547-gwm247同时检测到控制5个性状的QTL, 解释的表型变异率为8.14%~44.00%。小麦茎秆强度与厚径比、壁厚均呈极显著正相关, 与髓腔直径呈极显著负相关。利用gwm247选育茎秆实心度优于宁麦13的可靠性较高。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|