
{kind=link}
{kind=link}
{kind=link}
新型香稻渝恢2103香味分子遗传特性分析
[王春萍1, *  , 张现伟
, 张现伟2, * , 白文钦1 , 蒋晓英1 , 吴红1 , 林清1 , 唐永群2 , 姚雄2 , 张巫军2 , 唐荣莉1 , 李经勇2, * , 雷开荣1, * ]
, 张现伟, 白文钦, 雷开荣]
|
|


第一作者联系方式: 王春萍, E-mail: wcp49930000@sina.com, Tel: 023-65717006; 张现伟, E-mail: ycty2006@126.com, Tel: 023-49847001
香味是优良稻米品质的重要衡量标准之一, 2-乙酰-1-吡咯啉(2AP)是最主要的香味物质, 然而2AP生物合成机理至今仍未确凿。本研究筛选了与2AP生物合成密切相关的甜菜碱脱氢酶2基因( Badh2)在30份水稻材料中的3种突变类型, 从中发现1份新的香稻材料渝恢2103, 该材料 Badh2基因序列编码区无突变, 遗传分析显示渝恢2103与 badh2-E7突变型香稻宜香1B香味基因不等位, 与非香稻杂交F2香与非香分离比接近9∶7, 与香稻杂交F2香与非香分离比接近7∶9, 表明渝恢2103的香味受多基因控制。进一步利用实时荧光定量PCR技术(qRT-PCR)比较了与2AP生物合成相关基因在日本晴、渝恢2103和宜香1B的表达情况, 结果显示, Badh2基因在日本晴和渝恢2103中表达差异不大, 但在宜香1B中表达量异常高; 多数脯氨酸与谷氨酸代谢途径相关基因在宜香1B中的表达水平显著高于日本晴和渝恢2103; 推测宜香1B的2AP合成同时受 Badh2基因以及脯氨酸与谷氨酸代谢途径相关基因的影响; 渝恢2103香味形成可能与这些基因无必然联系。渝恢2103特殊的遗传特性可能为水稻香味形成机理研究提供新的突破点。
, ZHANG Xian-Wei, BAI Wen-Qin, LEI Kai-Rong
A major component 2-acetyl-1-pyrroline (2AP) in fragrance is one of the important indices of high-quality rice, however, the biosynthetic pathway of 2AP has not been demonstrated clearly. The betaine aldehyde dehydrogenase 2 (BADH2) is considered to be closely related to 2AP biosynthesis. In this study, three mutations in Badh2 gene were screened from thirty rice varieties and a new fragrant rice variety Yuhui 2103 was discovered. The new variety has no mutation in the coding region of Badh2gene and the Badh2allele of Yuhui 2103 can complement the defect of the badh2-E7 allele in Yixiang 1B. From crosses between Yuhui 2103 and non-fragrant varieties the segregating ratios of F2 fragrant to non-fragrant individuals were 9:7, while from crosses between Yuhui 2103 and fragrant varieties the segregating ratios of F2 fragrant to non-fragrant individuals were 7:9, indicating that fragrance of Yuhui 2103 is not controlled by only one gene. Furthermore, expression patterns of genes involved in 2AP biosynthesis were examined by the quantitative real-time PCR (qRT-PCR) in Nipponbare, Yuhui 2103, and Yixiang 1B. There was no significant difference in transcription level of Badh2 in Nipponbare and Yuhui 2103, however, the transcription level of Badh2 in Yixiang 1B was unexpectedly high. The expression levels of most of the genes involved in proline and glutamatic acid metabolism were significantly higher in Yixiang 1B than that in Nipponbare and Yuhui 2103. It is proposed that 2AP biosynthesis in Yixiang 1B is both influenced by Badh2 and the genes involved in proline and glutamatic acid metabolism, however, there is no necessary relationship between these genes and the fragrance of Yuhui 2103. The novel genetic characters of Yuhui 2103 may bring new breakthrough to the study of fragrance formation mechanism in rice.
香米以其清香可口的特色深受消费者喜爱和国内外稻米市场的青睐, 也激励着育种研究者不断发掘香稻基因资源和培育新的香稻品种。目前大部分研究结果表明水稻香味是由单隐性基因控制, 位于第8染色体上的甜菜碱合成酶2基因(Badh2)突变导致功能丢失, 使得香味物质2-乙酰-1-吡咯啉(2AP)合成前体物质积累, 最终大量集聚2AP而形成水稻香味[1, 2, 3, 4]。
目前, 对2AP生物合成途径的研究已取得一些进展, 但仍不够确凿。Romanczyk等[5]用15N标记脯氨酸和谷氨酸, 用16C标记葡萄糖, 结果在培养的蜡样芽孢杆菌产出的2AP中发现了15N和16C, 表明脯氨酸和谷氨酸可能是2AP合成的氮源, 葡萄糖则可能是2AP合成的碳源。但在16C标记实验中还观察到大量其他同位素离子峰, 表明有其他C原子的掺入。Yoshihashi等[6]研究发现2AP的形成与脯氨酸含量相关, 认为L-脯氨酸是2AP合成的主要前体物质, 为吡咯啉环的形成提供氮源[7]。1-吡咯啉是2AP合成的前体, 多数研究认为1-吡咯啉是通过γ -氨基丁醛(GABald)自发环化形成, GABald是脯氨酸代谢的中间物质, GABald量的多少受菜碱脱氢酶2 (BADH2)活性影响, BADH2具有催化γ -氨基丁醛(GABald)的功能, 在Badh2基因发生突变的香稻品种中, BADH2功能缺失, 导致GABald的积累, 从而增加该物质转向2AP的生物合成[4, 8]。也有研究发现1-吡咯啉是吡咯啉-5-羧酸脱羧形成, 提高吡咯啉-5-羧酸水平可增加2AP的积累, 吡咯啉-5-羧酸是谷氨酸代谢的中间产物, 受谷氨酸代谢相关酶类如吡咯啉-5-羧酸合成酶 (P5CS)活性影响[9]。关于2AP乙酰基的来历, 研究认为丙酮醛能够提供乙酰基与1-吡咯啉生成2AP[10], 也有研究认为丙酮醛能够与吡咯啉-5-羧酸在室温下直接生成2AP[9]。总之, 2AP生物合成途径十分复杂, 其具体生成过程及其与Badh2基因的联系还需进一步研究验证。
在香稻Badh2基因上游调控区域和外显子区域已发现20余种突变[11], 其中频率最高的是位于第7外显子的8碱基缺失和3个SNP位点[1], 其次是位于第2外显子7碱基的缺失[12]。Shi等[13]在香稻南海138的Badh2基因5° 非编码区发现3个碱基缺失, 在启动子区域有8个碱基的缺失, 并且发现badh2-E7型突变也存在启动子区域8个碱基的缺失, 但不存在5° 非编码区3个碱基缺失。然而也有些研究发现有些水稻品种Badh2基因序列与非香稻相同, 但仍具有香味。Fitgerald等[14]分析了313份来自世界各地的香稻材料Badh2基因第7外显子突变情况, 发现279份材料存在8碱基缺失, 15份材料不存在突变, 19份材料为杂合型, 因此推断还有其他基因导致2AP的合成。但其未对这15份材料进行进一步的Badh2基因序列分析和遗传分析, 因此也不排除这15份材料包含之后发现的badh2-E2、badh2-E8等突变类型, 还有可能包含其他至今都未发现的基因类型。Amarawahi等[15]利用香稻Pusa1121与非香稻1342杂交建立重组自交系(RIL), 定位到3个分布在水稻第3、第4、第8染色体上的与香味相关的位点, 并且发现所有浓香型和82%具有中等香味或淡香型的RIL在Badh2基因第7外显子处有8碱基缺失, 难以置信的是68.5%非香型的RIL也含有8碱基缺失, 因此, 他认为水稻香味不仅受Badh2基因1个基因控制, 甚至Badh2基因可能只是真正控制香味性状基因的1个标记。随后, 该研究团队又利用转录组技术结合前期的QTL定位结果在第3、第4、第8染色体上发现10个差异表达的基因, 其中第3染色上1个基因, 第4染色体8个基因, 第8染色体1个Badh2基因[16], 再次证明水稻香味由多基因控制。Cheng等[17]研究发现香稻Bamati 370和非香稻杂交F2只有15.6%的单株香味能达到亲本Bamati 370香味程度, 表明水稻香味为多基因控制, 通过基因定位他们在水稻第8染色体上发现了4个与香味相关的位点。因此, 水稻香味产生的原因还存在多种不同研究结果, 需要不断寻找新的突破点来阐明。
本研究以30份水稻材料, 分析其在Badh2基因第2外显子、第7外显子和启动子区域的突变情况, 重点围绕其中的1份特殊香稻材料渝恢 2103, 分析其Badh2基因序列特点, 香味遗传特性, 以及2AP生物合成途径相关基因的表达特点, 为水稻香味形成机理研究提供新的参考。
本实验室保存和重庆市农业科学院重庆市再生稻研究中心提供的30份水稻材料, 即日本晴、渝恢 2103、宜香1B、渝香糯1号、中香1号、559、560、561、十里香、2014特1、2014特2、2014特3、2014特4、香软稻、黄金稻、黑丰香糯、太香2号、丰黑2号、黑香糯、高产黑米、绿米、绿玉香、天龙香、海南万里香、千里香、红米、黑香旱米、人参黑宝稻、超优30和香粘。
采用KOH浸泡法鉴定叶片香味[18]。植株孕穗前期, 取植株叶片鲜重1 g左右, 剪成碎片放入小玻璃瓶中, 加入10 mL 1.7%的KOH溶液, 立即盖紧瓶盖, 在室温下保持10 min, 然后逐一打开瓶盖迅速嗅香味有无, 每份材料3次重复, 每次重复由不同实验人员完成。
采用咀嚼法鉴定米粒香味[19]。随机选取每株一穗进行鉴定, 如果首先连续5粒种子都有香味则认为该植株所有种子都有香味; 否则要咀嚼16粒种子, 如果16粒种子全不香则认为该植株所有种子都没有香味; 如果16粒种子中香与不香的米粒同时存在则认为存在香味性状分离。
根据Shi等[12]分别合成Badh2基因第2外显子和第7外显子突变检测引物FMbadh2-E2A和FMbadh2-E7, 根据Shi等[13]合成Badh2基因启动子区域突变检测引物STS。分别以30份水稻材料基因组DNA为模板, 进行PCR扩增, PCR体系为10 μ L, 含5 μ L 2× Es Taq MasterMix (康为世纪)、0.5 μ L基因组DNA (100 ng μ L-1)、上下游引物各0.5 μ L (10 µ mol L-1), ddH2O补足体积。PCR程序为94℃预变性5 min; 94℃变性30 s、55℃退火30 s、72℃延伸30 s, 共30个循环; 最后72℃延伸10 min。扩增后, 利用6%的聚丙烯酰胺凝胶电泳检测PCR产物。
主要按照Shi等[12]的方法合成Badh2基因克隆引物, 参照日本晴Badh2基因组序列(GenBank登录号为AP004463)设计Badh2基因启动子区域引物, Badh2PRM1-1-F: 5′ -CATCGAGATAGGCAGGCACA A-3′ , Badh2PRM1-1-R: 5′ -GAACCGCCAAGATCAC TTGTCT-3′ ; Badh2PRM2-1-F: 5′ -GGTTTCGAGTTT AGTCGAGA-3′ , Badh2PRM2-1-R: 5′ -GCCTACTGT ATCCTCCTCAT-3′ 。分别以非香水稻日本晴, 香型水稻渝恢2103、宜香1B和黑香糯基因组DNA为模板, 进行PCR扩增, PCR体系为20 μ L, 含10 μ L 2× Es Taq MasterMix (康为世纪)、1 μ L基因组DNA (100 ng μ L-1)、上下游引物各1 μ L (10 µ mol L-1), ddH2O补足体积。PCR程序为 94℃预变性5 min; 94℃变性30 s、58~60℃退火30 s、72℃延伸2 min, 共30个循环; 最后72℃延伸10 min。扩增后, PCR产物经1% 琼脂糖凝胶电泳, 分析得到目的条带, 用普通DNA产物纯化试剂盒(OMEGA)纯化。将纯化产物链接pMD19-T载体(TaKaRa)并转入JM109感受态细胞(TRANCE), 筛选阳性菌落送立菲生物技术有限公司测序。利用NCBI在线数据库、EditSeq和MegAlign软件进行基因序列分析。
配制杂交组合渝恢2103(香)/宜香1B(香)、渝恢2103(香)/日本晴(非香)和渝恢2103(香)/R838(非香), 获得F1种子, 用F1植株叶片鉴定香味。进一步通过F2种子及叶片香味性状确定香味分离比, 用χ 2进行适合性检验。
分别参照Chen等[4]和Forlani等[20]的方法合成Badh2基因、内参Actin基因和谷氨酸、脯氨酸代谢途径相关基因实时荧光定量PCR引物。用RNA提取试剂盒(艾德莱生物)分别提取日本晴、渝恢2103和宜香1B茎段、叶片和扬花后10 d幼穗RNA。用反转录试剂盒(Bio-Rad)对RNA进行去基因组DNA和反转录得到cDNA。用Bio-Rad CFX96 Real-time System荧光定量PCR仪进行荧光定量PCR, 反应体系20 μ L, 含iQ SYBR Green Supermix (Bio-Rad) 10 μ L、上下游引物各0.5 μ L、cDNA模板1.0 μ L, ddH2O补足体积。每个样品重复3次。
利用KOH浸泡法鉴定叶片和咀嚼法鉴定籽粒, 发现有12份材料为香稻, 包括宜香1B、渝恢 2103、渝香糯1号、561、中香1号、2014特3、香软稻、黄金稻、太香2号、丰黑2号、黑香糯和黑丰香糯, 其余18份材料为非香稻。
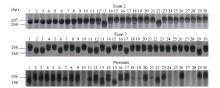
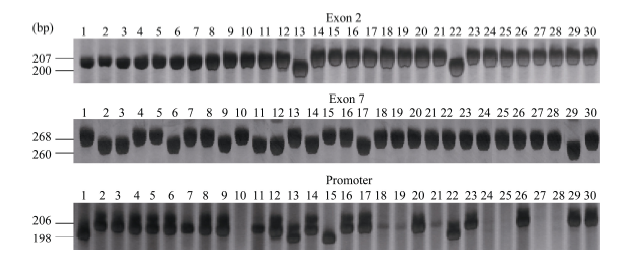 | 图1 30份水稻材料Badh2基因突变位点 泳道1~30分别为日本晴、宜香1B、渝香糯1号、渝恢 2103、十里香、中香1号、2014特1、2014特2、2014特3、2014特4、香软稻、黄金稻、黑丰香糯、太香2号、高产黑米、绿米、丰黑2号、绿玉香、天龙香、海南万里香、千里香、黑香糯、红米、黑香旱米、人参黑宝稻、超优30、559、560、561和香粘。Fig. 1 Mutation sites of Badh2gene in 30 rice varieties Lanes from 1 to 30 are Nipponbare, Yixiang 1B, Yuxiangnuo 1, Yuhui 2103, Shilixiang, Zhongxiang 1, 2014 Te 1, 2014 Te 2, 2014 Te 3, 2014 Te 4, Xiangruandao, Huangjindao, Heifengxiangnuo, Taixiang 2, Gaochanheimi, Lyumi, Fenghei 2, Lyuyuxiang, Tianlongxiang, Hainanwanlixiang, Qianlixiang, Heixiangnuo, Hongmi, Heixianghanmi, Renshenheibaodao, Chaoyou 30, 559, 560, 561, and Xiangzhan, respectively. |
在30份水稻材料中共检测到2份材料为第2外显子突变, 9份材料为第7外显子突变, 这11份材料全部为香稻。香稻渝恢2103的Badh2基因在第2外显子和第7外显子未发生任何突变。进一步检测发现有21份材料在Badh2基因启动子区域存在8碱基插入, 其中包括渝恢2103在内的10份香稻和11份非香稻材料, 日本晴、高产黑米和第2外显子发生突变的黑丰香糯和黑香糯4份材料未发生突变, 另外5份材料未扩增出目标条带(表1和图1)。
| 表1 30份水稻材料Badh2基因突变位点 Table 1 Mutation sites of Badh2gene in 30 rice varieties |
使用12对引物分别对渝恢2103、宜香1B和黑香糯的Badh2基因序列进行分段克隆, 最后拼接, 获得该基因包括起始密码子上游2500 bp在内的约9300 bp全基因序列。通过与日本晴Badh2基因序列比对发现, 3份材料与日本晴相似度可达99%, 宜香1B在第7外显子处存在8碱基缺失和3个单碱基突变, 黑香糯在第2外显子存在7碱基缺失。渝恢2103在编码区没有突变, 只在启动子区域与宜香1B都有8碱基插入, 黑香糯不存在这一突变, 这与2.2中的结果一致。3份材料在第2内含子都存在2个碱基缺失, 渝恢2103第4内含子出现了10个碱基插入, 宜香1B和黑香糯在该处出现了8个碱基插入。
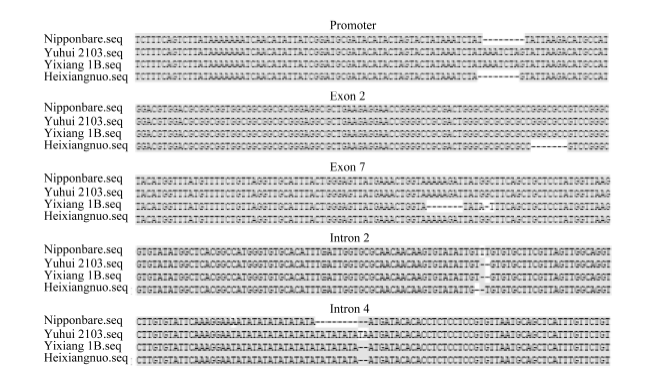 | 图2 Badh2基因主要突变位点分布Fig. 2 Distribution of main mutation sites of Badh2gene |
渝恢2103/宜香1B、渝恢2103/R838 (非香)以及渝恢2103/日本晴组合F1单株都不香(表2)。渝恢2103/宜香1B组合F2香与非香分离比接近7∶ 9 (χ 2=0.203), 渝恢 2103/R838和渝恢2103/日本晴两个组合F2香与非香分离比接近9∶ 7 (χ 2=3.80, 1.06)。
| 表2 渝恢2103杂交后代香味分离情况 Table 2 Segregation of fragrance in hybrid offspring of Yuhui 2103 |
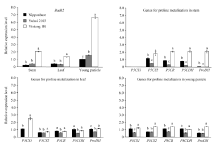
在3份材料日本晴、渝恢 2103和宜香1B中, Badh2基因在茎、叶和幼穗中的表达趋势基本相同, 即在宜香1B中的表达量显著大于渝恢2103和日本晴, 尤其在宜香1B幼穗中表达量异常高, 渝恢2103和日本晴差异不大(图3)。谷氨酸和脯氨酸代谢相关基因P5CS1、P5CS2、P5CR、ProDH和P5CDH在几份材料茎、叶和幼穗中的表达趋势基本相同, 即在宜香1B中表达量最高, 在日本晴中其次, 在渝恢2103中最低。
 | 图3 2AP生物合成相关基因表达情况 P5CS1: 吡咯啉-5-羧酸合成酶1; P5CS2: 吡咯啉-5-羧酸合成酶2; P5CR: 吡咯啉-5-羧酸还原酶; P5CDH: 吡咯啉-5-羧酸脱氢酶; ProDH: 脯氨酸脱氢酶。图柱上不同小写字母表示同一部位或同一基因的差异达0.01显著水平。Fig. 3 Expression profile of genes involved in 2AP biosynthesis P5CS1: δ 1-pyrroline-5-carboxylate synthetase 1; P5CS2: δ 1-pyrroline-5-carboxylate synthetase 1; P5CR: δ 1-pyrroline-5-carboxylate reductase; P5CDH: δ 1-pyrroline-5-carboxylate dehydrogenase; ProDH: proline dehydrogenase. Bars superscribed by different letters within the same part or the same gene are significantly different at P< 0.01 level. |
近年来, 大多数水稻香味研究是建立在承认Badh2单基因控制香味的思维模式上, 导致寻找香味新品种时也多是围绕该基因的突变进行的, 然而进一步对突变位点与基因功能之间联系的研究很少。在目前发现的Badh2基因突变类型中, 除了最普遍的第2外显子和第7外显子突变外, Shi等[13]发现在香稻南海138以及badh2-E7型突变的Badh2基因启动子区域有8个碱基的插入, 本研究发现除了badh2-E7型突变外, 有包括渝恢2103在内的10份香稻和另外11份非香稻材料都能检测到这种突变, 说明该启动子区域的突变与水稻香味没有必然的联系。另外, 还有5份材料不能扩增出目标条带, 推测在该区域还有其他突变位点。因此, 在Badh2基因中的一些突变尽管位于基因表达调控区域甚至是编码区域, 但可能只是其遗传进化中的一个简单事件, 在香味的形成中并未起到太大作用。目前针对Badh2基因突变开发的一些分子标记, 在实际辅助育种中的有效性还有待于证实。Badh2基因内含子区域突变与香味形成的相关性还未见报道, 本研究3份香稻材料Badh2基因序列第2和第4内含子出现了突变, 在GenBank公布的苏御糯(EU770320.1)和武香粳(EU770321.1) 2份香稻的Badh2基因序列第2内含子也出现了相同突变, 在第4内含子分别出现了10碱基和6碱基插入。这两处突变是否存在于所有香稻的Badh2基因序列中, 其与水稻香味形成是否存在相关性, 有待进一步研究验证。继续开展对已有突变类型的深入研究和发掘香味形成相关新基因将对香味形成的机制阐明具有推动作用。
非香稻中并不是不合成2AP, 而是合成的量少[21, 22], 因此, 我们可以假设, 当与2AP合成相关的不同遗传位点聚集在一起相互作用的时候就导致不同量2AP生成, 从而形成不同程度香味的水稻材料, 但由于目前对香味程度鉴定技术方法的不成熟, 绝大多数现有的研究报道都只能对香味的有无进行判断。位于第8染色体上的Badh2基因是一个比较重要或者比较普遍存在的控制水稻香味的基因, 因为前人的定位研究都发现了这个位点[3, 15, 17, 21, 23]。本研究发现渝恢 2103与badh2-E7突变型香稻宜香1B杂交F1为非香, 说明渝恢2103香味基因位点与badh2-E7突变型不等位, 证明了其他控制香味的基因位点的存在。渝恢2103与非香稻日本晴和R838杂交F2香与非香都基本呈9∶ 7分离, 与香稻宜香1B杂交F2香与非香分离比接近7∶ 9, 表明当有了宜香1B的遗传物质掺入后, 其遗传后代具有香味的群体反而变小了, 证明存在多基因间的互作, 这其中包括结构基因和转录因子。
Chen等[4]比较了Badh2基因在香稻与非香稻中的表达差异, 研究结果表明香稻中Badh2基因表达量与非香稻中差异不大, Badh2基因突变导致其翻译水平降低, 并不影响转录水平。本研究发现Badh2基因在宜香1B中的表达水平却显著高于非香型日本晴和渝恢2103, 尤其在幼穗中表达量异常高。这一结果与前人的研究存在差异, 可能有2个原因: (1)不同品种间遗传背景存在较大差异, 同一基因在不同材料中的本底表达本身就存在较大差异。(2)是植物的一种生理调节, 宜香1B的Badh2基因第7外显子存在8碱基的缺失, 导致有功能的BADH2蛋白质含量降低, 于是增加Badh2基因转录水平的表达以调节之, 渝恢2103与日本晴间的差异不大, 也暗示渝恢2103香味的形成与Badh2基因没有直接联系。
Huang等[9]研究香稻中较高的2AP含量与香稻中较高的吡咯啉-5-羧酸水平有关, 而后者与吡咯啉- 5-羧酸合成酶(P5CS)基因表达水平的上调有关, 因此推测P5CS催化谷氨酸生成吡咯啉-5-羧酸, 吡咯啉-5羧酸降解成1-吡咯啉后与丙酮醛作用生成2AP。其实, 谷氨酸生成吡咯啉-5-羧酸的过程也是合成脯氨酸的一个关键步骤, 吡咯啉-5-羧酸在吡咯啉- 5-羧酸还原酶(P5CR)的作用下就生成脯氨酸, 脯氨酸又能在脯氨酸脱氢酶(ProDH)催化下生成吡咯啉- 5-羧酸, 吡咯啉-5-羧酸在吡咯啉-5-羧酸脱氢酶(P5CDH)作用下生成谷氨酸。因此, 脯氨酸与谷氨酸能够通过吡咯啉-5-羧酸以及几种关键酶建立起一个代谢循环[14]。本研究发现宜香1B中这几个关键酶基因的表达量整体趋势显著高于日本晴和渝恢2103。因此, 我们推测, 在宜香1B中有两条途径限制2AP的合成, 一条是脯氨酸代谢成GABald后生成1-吡咯啉的过程, 受BADH2蛋白限制, 另一条是谷氨酸或脯氨酸代谢生成吡咯啉-5-羧酸, 后者又生成1-吡咯啉的过程, 受脯氨酸和谷氨酸代谢循环中的几种关键酶影响, 渝恢2103香味形成可能与这些蛋白酶无必然联系。然而, 这种不同材料间的比较受遗传背景差异等因素限制, 结果有待进一步验证。本研究团队正在构建渝恢2103等材料杂交后代的重组自交系和近等位基因系, 从而消除遗传背景的差异, 为进一步揭示其香味分子遗传机制奠定基础。
渝恢2103是一份新的香稻材料, 该材料Badh2基因序列编码区无突变, 与badh2-E7突变型香稻宜香1B香味基因不等位, 与非香稻杂交F2香与非香分离比接近9∶ 7, 与香稻杂交F2香与非香分离比接近7∶ 9, 表明渝恢2103的香味受多基因控制。Badh2基因在日本晴和渝恢2103中表达差异不大, 但在宜香1B中表达量异常高; 多数脯氨酸与谷氨酸代谢途径相关基因(P5CS1、P5CS2、P5CR、P5DH、ProDH)在宜香1B中的表达水平显著高于日本晴和渝恢2103; 推测宜香1B的2AP合成同时受Badh2基因以及脯氨酸与谷氨酸代谢途径相关的基因影响, 渝恢2103香味形成可能与这些基因无必然联系。渝恢2103特殊的遗传特性可能为水稻香味形成机制研究提供新的突破点。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|