


{kind=link}
{kind=link}
{kind=link}
{kind=link}
假禾谷镰孢侵染小麦后3种植物激素相关基因的差异表达分析
[李永辉 , 陈琳琳, 孙炳剑, 王利民, 邢小萍, 袁虹霞, 丁胜利
, 陈琳琳, 孙炳剑, 王利民, 邢小萍, 袁虹霞, 丁胜利* , 李洪连* ]
, 陈琳琳, 孙炳剑, 王利民, 邢小萍, 袁虹霞, 丁胜利, 李洪连]
|
|


第一作者联系方式:E-mail:liyonghui0823@163.com
假禾谷镰孢( Fusarium pseudograminearum)是我国近年新发现的小麦病原真菌, 本研究目的是揭示植物激素信号传导途径中相关基因表达对假禾谷镰孢侵染的响应。利用假禾谷镰孢野生菌株WZ2-8A侵染小麦品种周麦24, 对接种后5 d和15 d样品进行转录组测序, 分析差异表达基因, 并对候选基因进行qRT-PCR验证。假禾谷镰孢侵染后, 小麦幼苗生长受到明显抑制, 根长、株高、根重和地上部分鲜重均显著降低。转录组分析结果表明, 植物激素信号传导途径中有29个基因差异表达, 涉及到生长素、细胞分裂素和脱落酸3种植物激素。在接种后5 d有11个差异表达基因, 其中2个上调, 9个下调; 在接种后15 d共有25个差异表达基因, 其中8个上调, 17个下调。在生长素信号传导途径中, 生长素输入载体AUX1差异表达, 影响生长素的极性运输, 从而影响小麦根部的细胞伸长。在细胞分裂素信号传导途径中, 起正调控作用的B-ARR上调表达, 推测其促进细胞分裂素的信号传导, 从而抑制细胞分裂, 与生长素协同作用, 造成小麦的长势减弱。在脱落酸途径中, 脱落酸受体PYR/PYL下调表达; 起到负调控作用的PP2C相关基因均上调表达。脱落酸使植物对真菌和细菌的抗性起到负调控作用, 其信号传导途径与茉莉酸/乙烯途径相互拮抗, 脱落酸信号传导途径的阻遏可能会使茉莉酸/乙烯途径信号通路打开。qRT-PCR结果基本能够和转录组测序结果相拟合, 说明在假禾谷镰孢侵染胁迫下, 脱落酸的信号传导被抑制可能是中抗品种周麦24对假禾谷镰孢产生一定的抗性的生理基础。
, CHEN Lin-Lin, SUN Bing-Jian, WANG Li-Min, XING Xiao-Ping, YUAN Hong-Xia, DING Sheng-Li, LI Hong-Lian
Fusarium pseudograminearum is a pathogenic fungus in wheat newly found in China. The purpose of this study was to disclose the responses of genes involved in the hormone signals pathway after F. pseudograminearum infection. Wheat cultivar Zhoumai 24 was infected by the wild-type strain WZ2-8A, and the seedling samples at five and fifteen days after inoculation (DAI) were subject to transcriptome analysis by RNA-seq method. The selected differentially expressed genes were subsequently validated by qRT-PCR assay. The infection of F. pseudograminearum had a negative impact on wheat growth with significantly decreased root length, seedling height, and fresh weights of root and shoot. A total of 29 genes expressed differentially in the signal transduction pathways of auxin, cytokinin (CTK) and abscisic acid (ABA). At 5 DAI, 11 genes were expressed differentially compared with the control group (without WZ2-8A inoculation), in which two and nine genes were up- and down-regulated, respectively. At 15 DAI, 25 genes were expressed differentially, including eight up-regulated and seventeen down-regulated genes. In the auxin signaling pathway, auxin input transporter AUX1 was differentially expressed, influencing polar transport of auxin and disturbing cell elongation of wheat roots. In the CTK signaling pathway, the up-regulating role of B-ARR suggests that it could facilitate CTK signal transduction to inhibit cellular division, resulting in weaker wheat growth along with the interaction of the auxin signaling pathway. In the ABA signaling pathway, ABA receptor PYR/PYL was down-regulated, while the negative regulator PP2C was up-regulated. ABA plays a negative role in regulating host resistance to fungi and bacteria and its transduction has the antagonistic action against the jasmine acid (JA) / ethylene (ET) pathway, i.e., the repression of ABA signaling pathway might result in the opening of JA/ET pathway. These results of transcriptome analysis were generally validated by the qRT-PCR evidence, indicating that inhibition of ABA signal transduction after F. pseudograminearum infection might be the physiological basis of moderate resistance to the fungus in Zhoumai 24.
假禾谷镰孢(Fusarium pseudograminearum)是我国黄淮麦区新发现的一种重要小麦土传病原真菌, 可引起小麦茎基腐病, 造成严重减产, 对小麦安全生产威胁很大。小麦茎基腐病最早于1951年在澳大利亚报道[1], 近年在我国山西、河南、河北、江苏、山东等省均有不同程度发生[2, 3, 4], 并且危害逐年加重[5]。引起小麦茎基腐病的主要病原为多种镰孢菌(Fusariumspp.), 包括假禾谷镰孢(F. pseudograminearum)、黄色镰孢(F. culmorum)、禾谷镰孢(F. graminearum)、木贼镰孢(F. equiseti)、燕麦镰孢(F. avenaceum)和三线镰孢(F. tricinctum)等[6, 7]。其中假禾谷镰孢和禾谷镰孢是我国黄淮麦区小麦茎基腐病的优势病原菌[8, 9, 10], 且假禾谷镰孢的致病力明显强于禾谷镰孢[7, 11]。
假禾谷镰孢侵染小麦幼苗, 造成小麦根部和茎基部变褐, 须根减少, 根系和植株发育受到明显抑制。植物激素作为信号分子, 在植物生长发育的各个阶段都起着重要的作用, 对根和地上器官细胞的生长和形态建成有很大影响, 参与植物对生物和非生物刺激的应答反应, 参与和调控植物对病害的抵抗[12]。比如生长素, 在调节植物与病原菌关系上发挥着重要作用, 在一些病原菌中, 生长素信号传导能削弱植物抗病防御反应, 加重病害发生; 阻断或抑制生长素信号通路能增强植物的抗病能力[13, 14]。研究表明, 生长素通过泛素连接酶复合体SCFTIR1/AFB的F-box亚基介导对Aux/IAA转录抑制因子家族的蛋白酶解来实现对基因的表达调控。生长素输入载体AUX1对IAA极性运输和原生韧皮部运输途径都是必需的[15]。根形成要求AUX1促进侧根冠和表皮组织生长素的向基运输[16, 17]。SAUR (small auxin-up RNA)基因家族受生长素诱导而快速强烈应答的一类基因, 是植物特有的、生长素响应因子中最大的一个家族[18]。两个不同研究小组报道, SAUR 主要在大豆和玉米延伸组织中表达, SAUR基因在生长素所调节的细胞伸长过程中发挥特异的作用[19, 20]。细胞分裂素(cytokinin, CTK)是促进植物细胞分裂的因子, 具有广泛的生理功能参与许多生理和发育的调控过程, 如影响根、茎的分生组织中分裂细胞的数目和分生组织的大小, 进而调控植物主根生长和侧根发育[21, 22]。Hwang等[23]发现ARR1、ARR2和ARR10三种B型ARR (Arabidopsis response regulator)过量表达时, 植物基部的细胞分裂素水平大大增加。脱落酸(abscisic acid, ABA)最初被认为是一种抑制生长的植物激素, 随着研究的深入, 发现脱落酸在非生物和生物等逆境胁迫下的调节中发挥重要作用, 如干旱胁迫下调控主根和侧根发育, 维持植物对水分的需求[24, 25]; 脱落酸也可以和水杨酸、茉莉酸、乙烯等直接参与抗病反应的信号相互作用, 从而影响植物的抗病性[26]。对脱落素信号传导途径的脱落酸受体, 一度存在争议, 目前研究最系统的脱落酸受体是PYR/PYL (pyrabactin resistance/PYR-like), SNF1 (sucrose non-fermenting 1)相关的蛋白激酶SnRK2 (Snf1-related kinase 2)家族起正调节因子作用和蛋白磷酸酶PP2C起负调节因子作用, 从而调控ABA信号传导及其下游反应[27, 28, 29]。Di等[30]在研究土壤习居菌尖孢镰孢侵染发病过程中植物激素的变化时发现, 生长素、乙烯、脱落酸等植物激素影响尖孢镰孢侵染定殖和寄主植物的抗感病性。同样为土壤习居菌的假禾谷镰孢胁迫影响小麦植物激素的研究未见有报道。本研究通过转录组测序分析小麦在假禾谷镰孢侵染后植物激素信号传导途径差异表达的基因, 并用qRT-PCR方法进一步验证, 目的是了解假禾谷镰孢侵染发病过程中寄主小麦植物激素的应答调控, 为该病害的防治提供新的思路。
供试小麦品种为周麦24, 对假禾谷镰孢(F. pseudograminearum)在田间表现中抗。接种体为假禾谷镰孢强致病力菌株WZ2-8A, 由本课题组从河南武陟县田间病株上分离并保存。将小米煮1~2 min后, 装入三角瓶, 灭菌, 接入新鲜的WZ2-8A菌饼, 25° C温箱培养7 d, 待菌丝布满培养基后备用。将病菌培养物以0.5%接菌量与灭菌土混匀, 装入育苗钵, 作为处理组, 以不接菌的灭菌土作为对照组。以3%次氯酸钠处理种子5 min, 用无菌水冲洗后, 放入带滤纸片灭菌的培养皿内, 置25° C温箱保湿24 h, 然后播入上述育苗钵, 每钵10~15粒。每个取样时间点随机取处理组和对照组各9钵, 每3钵收取的小麦根和茎基部样品作为一个重复, 每处理3次重复。
取处理组和对照组接种后3、5、10、15和20 d的小麦幼苗, 分别测量其根长、株高及根部和地上部鲜重, 3次重复, 每重复7~10株。使用Microsoft Excel 2013软件统计小麦生长指标进行, 使用t测验检验显著性。
取接种后5 d和15 d的处理组以及5 d的对照组麦苗, 用水冲去泥土, 用滤纸吸掉多余的水分, 剪取小麦根部和茎基部, 称取每处理2个重复的样品(每份至少1 g), 送广州基迪奥公司测序, 测序深度为10 G; 将余下样品装入自封袋中用液氮处理, -80 ° C保存, 用于后续验证实验。使用OmicShare Tools云平台(http://www.omicshare.com/tools)上的应用程序对测序数据作进一步的个性化分析。
按照TRIzol试剂盒(Thermo Fisher)说明书提取每个样品的总RNA, 之后从每个样品取0.5 μ g RNA用于cDNA合成, 用反转录试剂盒PrimeScript RT reagent Kit with gDNA Eraser (RR047a, TaKaRa), 按说明书首先去除基因组DNA污染, 反应体系包括5× gDNA Eraser buffer 2 μ L, gDNA Eraser 1 μ L, Total RNA 0.5 μ g, 加RNase Free ddH2O至10 μ L, 混匀反应液, 置反应管于PCR仪上, 42° C 2 min; 然后取出反应管, 冰浴, 再向反应管中加入反转录试剂(PrimeScript RT Enzyme Mix I 1 μ L, RT Primer Mix 1 μ L, 5× PrimeScript buffer II 4 μ L, RNase Free ddH2O 4 μ L), 混匀, 置反应管于PCR仪上, 37° C 15 min, 85 ° C 5 s, 反转录成20 μ L cDNA。之后通过PCR扩增验证cDNA中是否有基因组DNA污染, 扩增引物序列为F:5° -CACGCCGTTCATTGAATCCT-3° 和R:5° -GCAAGCCGAGTGTAGAAGAG-3° 。cDNA扩增的目标片段大小为205 bp, 以小麦基因组DNA为模板的扩增产物为1716 bp的片段, 通过条带大小的不同可证明cDNA中没有基因组DNA污染, 可以用作模板进行候选基因的实时定量PCR验证。PCR验证使用2× Taq PCR MasterMix (KT201, 天根生物科技公司), 该混合体系包含Taq DNA聚合酶、dNTPs、MgCl2和PCR反应的增强剂和优化剂以及稳定剂, 实际扩增体系为25 μ L, 其中包括12.5 μ L 2× MasterMix, 上下游引物各0.5 μ L (终浓度0.2 μ mol μ L-1), cDNA 0.5 μ L, ddH2O 11 μ L。扩增程序为95 ° C 预变性5 min, 然后按95° C 30 s、62° C 40 s和72° C 90 s进行30个循环, 最后72 ° C延伸7 min。
测序数据经分析后, 将选取的候选基因进行实时荧光定量PCR验证, 以actin为内参基因来评价候选基因的相对表达量。使用Primer5软件为内参基因和候选基因设计实时荧光定量PCR引物(表1), 使用MasterCycler RrealPlex4实时荧光定量PCR仪(Eppendorf)和SYBR Premix ExTaq (RR420a, TaKaRa)荧光定量试剂盒对候选目标基因进行定量PCR验证。反应体系为12 μ L, 包括2× SYBR Premix ExTaq6 μ L, 上下游引物各0.24 μ L (终浓度0.2 U μ L-1), cDNA 1 μ L, ddH2O 4.52 μ L; 反应条件为95° C 30 s; 95° C 5 s, 60° C 15 s, 68° C 20 s, 40个循环; 95° C 15 s, 60° C 15 s, 20 min升至95 ° C, 95° C 15 s, 以检测扩增产物的熔解曲线。每个样品3次重复。采用2-∆ ∆ Ct法[31]计算基因的相对表达量。在鉴定内参基因和候选基因具有相似扩增效率后进行上述检测反应, 如果扩增效率达不到要求, 重新设计引物, 再评价。
| 表1 qRT-PCR验证所用引物 Table 1 Primers used in qRT-PCR validation |
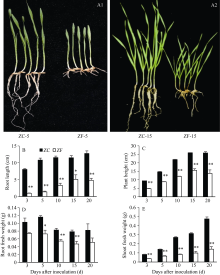
根据转录组测序分析结果, 重复进行室内假禾谷镰孢土壤接种侵染小麦实验。调查数据显示, 本次接种实验重复出了测序样品采集时的相似结果。与不接菌的对照相比, 接种后5 d的小麦根部开始变褐, 10 d和15 d的多变为褐色, 茎基部有褐色病斑和菌丝层, 个别小麦幼苗死亡。接种的小麦苗显著矮化, 这也和最初供试菌株致病性测定的症状相似, 说明假禾谷镰孢能够明显抑制小麦的生长(图1-A1, A2)。测量发现, 假禾谷镰孢能够明显抑制小麦的生长。受到假禾谷镰孢侵染后, 小麦根长(图1-B)、株高(图1-C)、根部鲜重(图1-D)和地上部鲜重(图1-E)均与对照组均有显著差异。
 | 图1 小麦受假禾谷镰孢侵染后的症状及对植株生长的影响 A1和A2分别为接种后5 d和15 d的小麦。* 和* * 分别表示处理组(ZF)与对照组(ZC)之间有显著(P < 0.05)和极显著差异(P< 0.01)。Fig. 1 Symptom and effect on growth of wheat plant infected by F. pseudograminearum A1 and A2 show wheat growth at 5 days and 15 days after inoculation, respectively. * and * * indicate significant difference between the treatment group (ZF) and the control group (ZC) atP< 0.05 and P< 0.01, respectively. |
转录组测序结果表明, 只有生长素、细胞分裂素和脱落酸这3种激素的信号传导途径有基因显著性差异表达。在接种5 d后, 共有11个相关基因差异表达, 其中2个上调表达, 9个下调表达; 在接种15 d后, 共有25个相关基因差异表达, 其中8个上调表达, 17个下调表达。生长素中差异表达基因主要为下调表达, 其中AUX1基因只在接种后5 d差异表达, 其中1个上调表达, 1个下调表达; SAUR基因在侵染后5 d有6个下调表达, 在接种后15 d有8个下调表达; 细胞分裂素途径中的B-ARR基因在接种后5 d有1个上调表达, 1个下调表达, 接菌后15 d有2个上调表达; 脱落酸受体PYR/PYL基因仅在接种后15 d差异表达, 且全部为下调表达, 负调控因子蛋白磷酸酶PP2C的基因仅在接种后15 d差异表达, 且全部为上调表达(表2)。
| 表2 假禾谷镰孢侵染诱导后植物激素差异表达基因数 Table 2 Number of differentially expressed genes related to plant hormones after inoculation with F. pseudograminearum |
细胞分裂素途径中的一个B-ARR基因Traes_7AS_503B57D77和脱落酸途径中的一个PP2C基因Traes_2AS_048E13951在接种后15 d差异表达倍数最大, 其log2FC可达到10以上; 而下调表达最明显的是1个SAUR基因Traes_5BL_80B9C8E10, 其log2FC值为-5.9, 其次是1个SAUR基因Traes_5DL_677150034和1个PYR/PYL基因Traes_1AL_B82F1573F, 它们的log2FC值均为-4.6 (表3)。
将各个差异表达基因表达量的FPKM (fragments per kilobase of transcript per million mapped reads)值取以10为底的对数(lg FPKM)制作热图能够直观看出这些基因的差异表达情况。图2显示, SAUR和PYR/PTL差异表达基因最多, 大部分为在接种5 d后下调表达的, 而在接种5 d和15 d后基因表达量变化不大。而B-ARR和PP2C的基因大部分上调表达。其中, B-ARR在接种后5 d至接种后15 d逐渐上调表达, PP2C的基因在接种后5 d不差异表达, 而在接种后15 d明显上调表达。
| 表3 假禾谷镰孢侵染诱导后植物激素相关基因差异表达倍数 Table 3 Fold change (FC) of differentially expressed genes related to plant hormones after inoculation with F. pseudograminearum |
 | 图2 植物激素途径中差异表达基因表达量热图 不同颜色表示基因表达量(lg FPKM)。ZC:对照组; ZF-5:接种后5 d; ZF-15:接种后15 d。Fig. 2 Heatmap of differentially expressed genes involved in plant-hormone pathways Gene expression levels in lg FPKM represent in different colors. ZC:control group; ZF-5:5 days after inoculation; ZF-15:15 days after inoculation. |
根据转录组测序分析结果, 重复了室内假禾谷镰孢土壤接种侵染小麦的实验, 在相应的设计时间点取样, 提取总RNA, 反转录获得cDNA。RNA电泳检测, 28S和18S条带间隔区清晰, 而5S条带应该是电泳过程中有部分RNA降解, 提取的总RNA可用进行下一步实验图显示(电泳图未列出)。反转录后的cDNA没有基因组DNA污染, 能够用于后续的qRT-PCR实验(图3)。设计了内参基因actin和9个候选基因设计实时荧光定量PCR引物, 经过扩增效率验证, 这些引物的扩增效率均在80%~120%之间, 具有相似的扩增效率, 可以用于候选基因的qRT-PCR验证。
选择参与生长素、细胞分裂素和脱落酸途径的9个基因(Traes_1DL_F70395059、Traes_7AL_ AEC178775、Traes_5BL_80B9C8E10、Traes_5DL_ D5D75CCDF、Traes_5DL_32B83E57E、Traes_7AS_ 503B57D77、Traes_2BL_616F08780、Traes_2AL_ 2FF604DA9和Traes_3DS_D7629B48F)进行qRT- PCR验证, 将上述9个基因分别命名为AUX1-1、SAUR-2、SAUR-3、SAUR-5、SAUR-6、B-ARR-1、SnRK2-1、SnRK2-2和PYR/PYL-1。生长素信号传导途径中的AUX1-1、SAUR-2、SAUR-3、SAUR-5和SAUR-6的qRT-PCR表达趋势与转录组分析结果相吻合; 细胞分裂素传导途径中的B-ARR-1, 虽然log2FC值和转录组数据相差较大, 但都上调表达。脱落酸传导途径中的SnRK2-1, 转录组分析显示接种5 d后的表达量与对照相比没有显著变化, 接种15 d后上调表达, 但是qRT-PCR结果显示在这两个时间点均下调表达; 类似地, SnRK2-2在转录组数据中接种5 d后的基因表达量没有明显变化, 而qRT-PCR结果显示该基因在接种5 d后上调表达, 接种15 d后下调表达(图4), 说明参与脱落酸传导途径的SnRK2-1和SnRK2-2的实时定量PCR分析结果与转录组测序结果不尽一致。
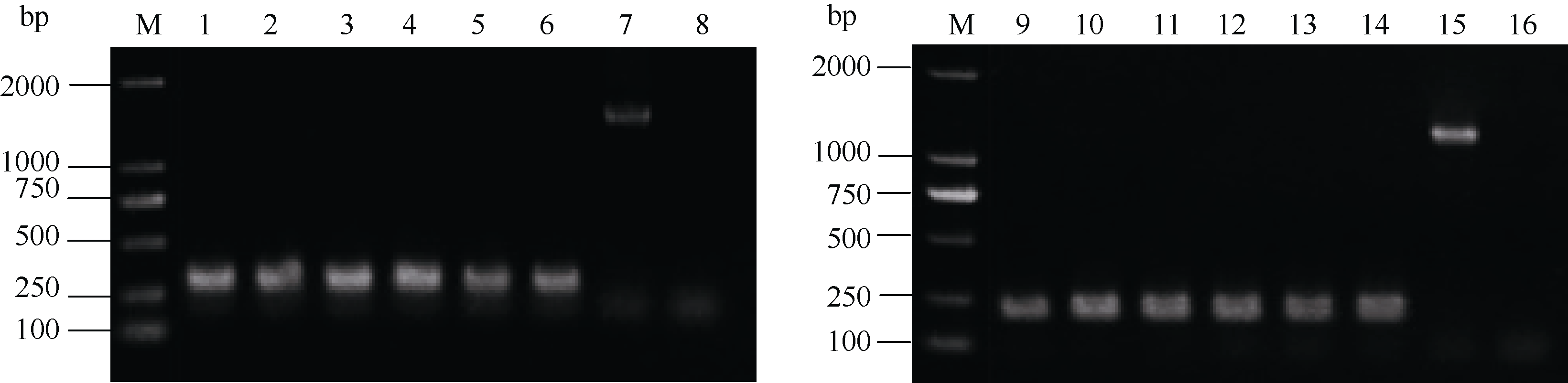 | 图3 cDNA中基因组DNA污染验证电泳图 M:DL2000 marker; 1~3:接种后5 d对照组; 4~6:接种后5 d处理组; 7和15:gDNA; 8和16:ddH2O; 9~11:接种后15 d对照组; 12~14:接种后15 d处理组。Fig. 3 Electrophoretic pattern of verification of genomic DNA contamination in cDNA pool M:DL2000 marker; 1-3:control group at 5 days after inoculation (DAI); 4-6:treatment group at 5 DAI; 7 and 15:gDNA; 8 and 16:ddH2O; 9-11:control group at 15 DAI; 12-14:treatment group at 15 DAI. |
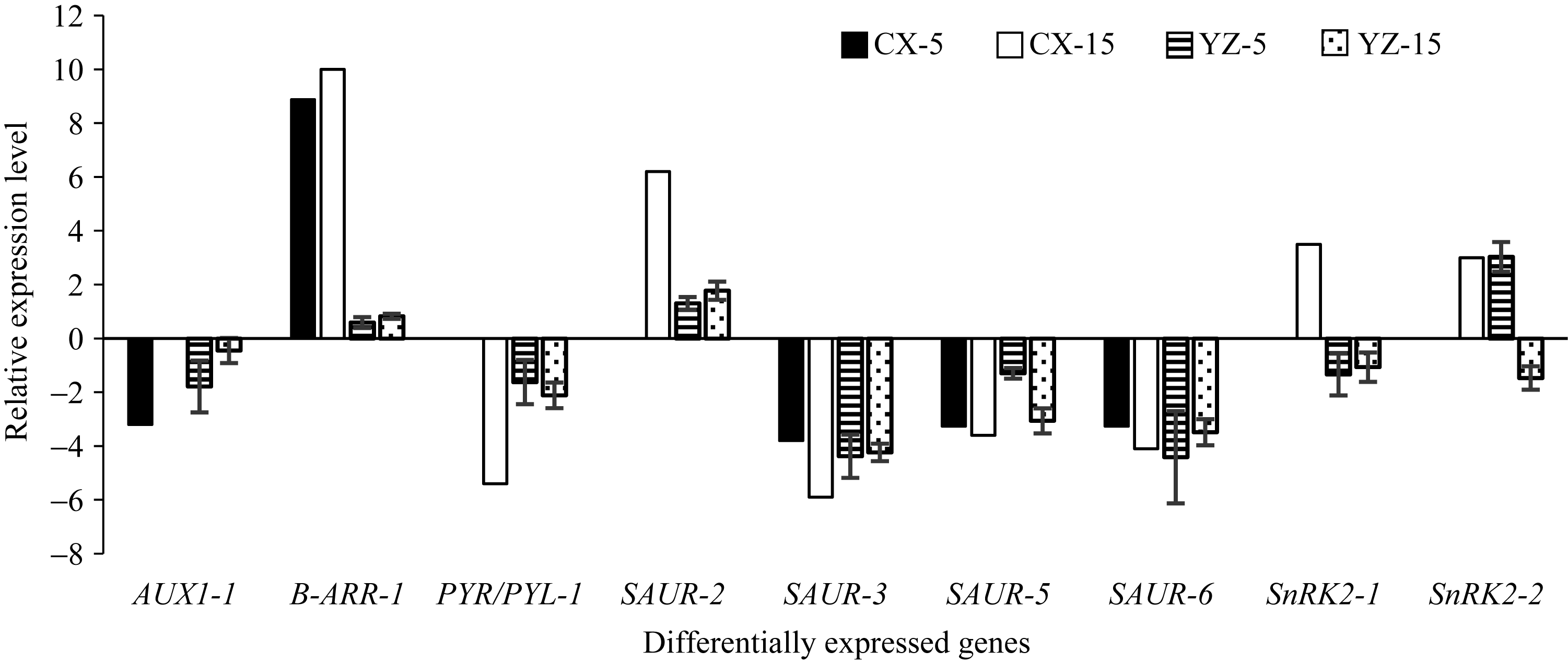 | 图4 差异表达基因qRT-PCR验证结果 CX-5和CX-15代表5 d和15 d测序样品; YZ-5和YZ-15代表5 d和15 d验证样品。Fig. 4 qRT-PCR validation for the differentially expressed genes CX-5 and CX-15 represent sequencing samples at 5 days after inoculation (DAI) and 15 DAI, respectively; YZ-5 and YZ-15 represent validation samples at 5 DAI and 15 DAI, respectively. |
植物激素调控着植物生长、发育、适应、抗逆等所有环节, 目前对非生物胁迫的激素调控机制报道较多, 而对生物胁迫, 特别是病原真菌侵染过程中的激素调控, 则研究较少[32, 33]。假禾谷镰孢是我国近年新发现的小麦病原真菌, 侵染后小麦幼苗表现为地上部矮化, 根部发育受到严重抑制。本研究发现, 该菌侵染小麦过程中, 生长素、细胞分裂素和脱落酸的信号调控相关的基因差异表达, 而且多数为显著下调表达, 推测这些差异表达基因可能与被侵染小麦的矮化和根生长发育受抑制的表型相关。AUX1是生长素输入载体, 其极性分布会影响侧根和不定根的发育以及根的向重性[34], 小麦受到假禾谷镰孢侵染后, AUX1-1基因下调表达, 然而另一个编码AUX1的基因却表现为上调表达。因此推测, 假禾谷镰孢侵染小麦后会在转录组层面对生长素输入载体AUX1造成影响, 从而影响小麦的生长, 但其作用机制还有待进一步研究。
生长素早期相应基因的3类家族(Aux/IAA、SAUR和GH3)中, SAUR家族基因受感应生长素速度最快的一类基因, 但目前大部分SAUR的功能仍不清楚, 大豆和玉米的SAUR基因的研究显示其主要在下胚轴和叶鞘伸长组织中表达, 表明SAUR基因具有特异的调控细胞伸长的作用[20, 35]。任怡怡等[36]利用原位杂交技术, 发现SAUR在茎的伸长区表达量较高, 而在根或其他器官中表达量较低或者不表达, 因此推测这类基因对生长素调节细胞伸长起到重要作用。本研究中, 假禾谷镰孢侵染小麦早期(5 d), 10个SAUR基因的表达显著被抑制, 间接说明是生长素的传导在假禾谷镰孢胁迫下受到抑制。细菌侵染拟南芥诱发IAA水平上升, 当抑制生长素信号传导, 则相应提高拟南芥对植物病原细菌的抵抗力[37]。假禾谷镰孢侵染小麦, 数个SAUR基因的表达显著下调, 推测真菌侵染后的小麦根部合成位点的生长素的转运受到抑制, 使SAUR基因转录水平显著下降, 根部总体生长素水平上升, 导致假禾谷镰孢侵染的小麦根部生长受抑制, 长势弱于对照组。但生长素信号传导受到抑制是否与周麦24对假禾谷镰孢侵染产生一定的抗性有关, 尚待进一步深入研究。
细胞分裂素(cytokinin)主要在根冠的平衡细胞合成, 经木质部向上运输到植物的各个器官和组织, 调节植物细胞的分裂和分化, 过量的细胞分裂素会减少根分生组织的大小, 根的正常发育往往是生长素和细胞分裂素协同作用的结果[38, 39, 40]。在细胞分裂素双元信号转导系统中, B型ARR是一类转录因子, 作为细胞分裂素的正调控因子转录下游靶基因。在细胞分裂素的2个差异表达基因中, 上调的量明显高于下调量, 因此推测其促进细胞分裂素的信号传导, 但其过量表达会使小麦的根部细胞变小, 长势变弱; 也有可能是因为B-ARR的过量表达对生长素合成酶基因YUCCA起负调控作用[41], 使生长素的生物合成受到抑制, 细胞分裂素和生长素信号传导途径的交互作用导致小麦长势变弱。
脱落酸在植物抗病中起非常重要的作用, 然而脱落酸在抗病反应中起正调控还是负调控作用的结论却不一致。丁新伦等[42]试验表明, 脱落酸可以调节胼胝质积累, 阻挡病菌侵入, 提高植物的抗病性, 如水稻对稻条纹病毒(Rice stripe virus, RSV)的抗性与胼胝质含量有关, 抗性强的水稻品种中胼胝质含量高。但更多学者认为, 脱落酸能够增强植物的感病性, 如玉米瘤黑粉菌(Ustilago maydis)、灰霉菌(Botrytis cinerea)和稻瘟病菌(Magnaporthe oryzae)侵染玉米、番茄和水稻的过程中发现, 真菌侵染早期植物脱落酸水平明显提高, 植物敏感性增强[43], 而脱落酸缺陷型突变体会使植物对病原菌的抗性增强[44, 45]。本研究发现, 脱落酸的8个受体基因在接种后5 d表达没有明显变化, 接种后15 d全部下调表达, 起负调控作用的PP2C基因在接种后15 d全部上调表达, 推测在周麦24受侵染早期, 脱落酸并没有起抵抗假禾谷镰孢的作用, 而在侵染后期脱落酸信号传导途径受到抑制, 从而使茉莉酸/乙烯途径信号传导途径表达, 使受到侵染的小麦植株产生一定的抗性。
候选基因的qRT-PCR和转录组测序所显示的表达趋势大部分是相一致的, 然而起到正调控作用的SnRK2的差异表达基因中的两个验证结果不符, 这一现象可能是转录组测序中样品cDNA文库PCR扩增步骤带来的某些偏好性造成的。qRT-PCR验证结果显示, 这两个基因在小麦受到假禾谷镰孢侵染后主要表现为下调表达, 推测其抑制了脱落酸信号传导途径; 也可能因为具有能量调控和对逆境应答反应等多种重要调控功能的SnRK2基因本身非常的敏感, 这种表达形式可能是对重复实验中病原菌侵染程度和寄主小麦发病程度差异的一种体现。进一步的解释还需要对发病小麦内源脱落酸进行定量测定。
在病原与寄主互作过程中的植物激素调节是个非常复杂的过程, 本文仅就假禾谷镰孢侵染过程中3种植物激素的表达进行了初步探讨, 以后的研究中还应考虑对不同激素的产生进行定量测定, 外源激素的恢复实验, 以及明确是否假禾谷镰孢也像其他真菌一样主动分泌激素来调节寄主植物根系生长发育, 以适应其侵染定殖。
假禾谷镰孢侵染小麦引起茎基腐病, 对小麦生长发育有显著影响。病菌胁迫下, 寄主小麦基因转录水平显著变化, qRT-PCR选择性分析传统植物激素相关通路基因的表达也基本能够和转录组测序结果相拟合。假禾谷镰孢在侵染致病过程中, 能够诱导小麦生长素、细胞分裂素和脱落酸信号传导途径的相关基因差异表达, 推测影响生长素的极性运输和增强刺激细胞分裂素途径信号传导, 两者协同作用影响小麦根部的细胞伸长, 造成小麦的长势减弱。在假禾谷镰孢侵染胁迫下, 脱落酸信号传导下调可能是周麦24对假禾谷镰孢侵染表现中抗的生理机制。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|