

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
黄淮海地区大豆主栽品种对8个大豆疫霉菌株的抗性评价
[李晓那1, 2, **  , 孙石
, 孙石2, ** , 钟超2 , 韩天富1, 2, * ]
, 孙石, 钟超]
|
|


第一作者联系方式: 李晓那, E-mail: 913714351@qq.com; 孙石, E-mail: sunshi@cass.cn
随着麦茬免耕栽培技术的推广应用, 黄淮海地区麦后夏播大豆生产中疫霉根腐病呈加重趋势。了解该地区大豆主栽品种对疫霉根腐病的抗性和筛选抗病亲本, 对培育新的高产广适抗病品种具有重要意义。本研究利用8个具有不同毒力的大豆疫霉菌株, 采用下胚轴创伤接种法, 对20世纪50年代以来黄淮海地区审定、推广的140个大豆主栽品种进行接种鉴定。表明除6个品种对8个菌株均无抗性外, 其余134个品种分别抗1~8个大豆疫霉菌株, 占鉴定品种总数的95.7%, 其中抗6~8个以上菌株的品种有83个, 占鉴定品种总数的59.3%。以14个鉴别寄主的抗病反应型为参照, 发现134个品种对8个大豆疫霉菌株共产生65种反应型, 其中19个品种产生的5种反应型与已知单基因或2个单基因组合反应型相同; 115个品种产生的60种反应型与含有已知单基因或2个单基因组合的反应型不同, 推测可能含有新的抗病基因或基因组合。根据研究结果合理选择亲本, 可培育出聚合多个抗性基因且综合性状优良的大豆新品种。
With the application of wheat stubble no-tillage cultivation technology, the occurrence rate of phytophthora root rot of soybean is getting higher in the Huang-Huai-Hai Rivers Valley. Therefore, it is crucial to investigate the resistance of the widely-grown soybean cultivars to phytophthora root rot and to screen disease-resistant resources for soybean improvement. In the current study, a total of 140 soybean cultivars, widely-grown in Huang-Huai-Hai Rivers Valley since the 1950s, were investigated for their response to eight strains of Phytophthora sojae through the hypocotyl inoculation method. Except that six cultivars were susceptible to all strains, 134 cultivars (95.7%) were resistant to 1-8 strains, including 83 cultivars (59.3%) resistant to 6-8 strains. A total of 65 reaction types in response to the eight strains were identified from the 134 resistant cultivars by means of 14 reference genotypes. Five reaction types produced by 19 cultivars fit the reaction-type profile of a known resistance gene or the combination of two genes. The remaining 60 reaction types by 115 cultivars were different from all reaction types of a single resistance gene or the combinations of two genes, indicating that theses cultivars might carry novel resistance gene(s)/gene combination(s) against P. sojae. These results provide a guide in parent selection for pyramiding resistance genes in soybean cultivars with good agronomic backgrounds.
近年来, 随着农村劳动力的转移和农村土地流转的加快, 黄淮海地区农业生产机械化水平逐步提高, 少耕免耕、秸秆还田等耕作栽培措施在生产上大面积推广应用[1], 这虽能起到保土、保水、增加土壤有机质的作用, 但也有利于病原菌的越冬、侵染和传播, 易造成作物根部病害流行和蔓延[2, 3]。据国家大豆产业技术体系调查, 近年来黄淮海地区大豆根腐病加重发生, 其中, 危害最为严重的是大豆疫霉根腐病(Phytophthora root rot, PRR)[4]。该病是一种土传性病害, 可以在大豆的任何生育期侵染植株并造成危害。培育和种植抗病品种是防治该病害最经济、有效和环境友好的方法[5]。大豆对疫霉根腐病的完全抗性由单显性基因控制[6]。迄今, 国内外已在大豆的9条染色体(第2、第3、第7、第10、第13、第16、第17、第18、第19染色体)上定位了30个抗疫霉根腐病基因, 其中11个是由我国研究人员鉴定的[5, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23]。大豆与大豆疫霉菌的互作为典型的基因对基因关系[24], 疫霉菌常因毒性基因的快速变异而导致高度的遗传分化, 特别是在抗病品种选择压力下, 新的毒力小种(毒力型)不断出现导致抗病品种抗性丧失[25, 26]。因此, 需要不断培育和利用新的抗病品种。大面积种植品种(以下称为“ 主栽品种” )由于产量高、综合性状好、适应性强, 常被育种家作为选育抗病品种的亲本, 因此, 解析主栽品种对疫霉根腐病的抗性对培育新的抗病品种具有重要意义。本研究对黄淮海地区20世纪50年代以来审定、推广的140个大豆主栽品种进行鉴定, 比较不同年代和不同省份主栽品种对大豆疫霉的抗性表现, 了解其抗性多样性, 初步推导它们所蕴含的抗病基因, 为黄淮海地区大豆抗病品种的选育和疫霉根腐病的防治提供依据。
140份材料来自黄淮海地区7省市(表1), 为20世纪50年代以来在该地区种植面积较大的大豆品种。此外, 选用14个含有已知抗大豆疫霉根腐病基因(Rps基因)的大豆品种(系)及感病对照品种Williams作为抗病基因的鉴别品种(表2)。
| 表1 不同年代黄淮海地区主栽大豆品种 Table 1 Main soybean cultivars of Huang-Huai-Hai Rivers Valley in different years |
| 表2 含已知抗病基因的大豆品种(系)对8个大豆疫霉菌株的抗性反应 Table 2 Reactions of soybean cultivars (lines) possessing Phytophthora resistance gene after inoculation with eight strains ofP. sojae |
8个具有不同毒力的大豆疫霉菌株分别为PsUSAR2 (毒力型: 1b、2、3c、5)、Ps41-1 (毒力型: 1a、1b、1d、2、3a、3b、3c、4、5、6、7、8)、PsMC1 (毒力型: 1b、1c、1d、1k、2、3a、3b、3c、4、5、7)、PsRace1 (毒力型: 1a)、PsRace3 (毒力型: 1a、1d)、PsRace4 (毒力型: 1a、1d、3b)、PsRace5 (毒力型: 1a、7)和PsJS2 (毒力型: 1a、1b、1c、1d、1k、2、3a、3b、3c、4、5、6、7、8)。疫霉菌株由单个孢子纯化后保存在V8汁琼脂培养基上, 接种鉴定前将菌株转入含1.5%琼脂的胡萝卜(CA)培养基培养皿中, 在25℃黑暗条件下培养7~10 d, 接种到Williams幼苗以恢复毒力, 植株发病后再进行病原菌分离和纯化, 接种一套寄主矫正毒力, 保证接种的准确性。
将140个供试大豆品种、14个鉴别寄主和感病品种Williams分别取15粒播种于以蛭石为基质、直径为10 cm的塑料花盆中, 出苗期温度控制在(25~29)° C, 出苗后保留10株长势一致的健康植株, 待第一对真叶(单叶)完全展开时(出土后7~10 d), 采用下胚轴创伤接种法接种[27]。用10 mL注射器上的针尖在大豆幼苗子叶下约0.5 cm处向下划开1 cm长的伤口, 深度不能超过茎粗的1/3, 然后将混匀的菌丝体注于伤口处。接种后在(18~25)℃条件下保湿48 h, 然后转入相同温度的温室继续培养。接种5~6 d后参照朱振东等[28]的抗性评价标准进行病情调查。植株死亡≥ 70%视为感病(susceptible, S), < 30%则为抗病(resistant, R), 在30%~70%之间为中间类型(intermediate, I)。试验过程中进行3次重复鉴定。
将中间反应类型归为抗病反应[29], 比较待鉴定品种与14个鉴别寄主对8个大豆疫霉菌株的反应型。若待鉴定品种的反应型与某个鉴别寄主的反应型或某2个鉴别寄主组合的反应型一致, 则说明产生这种反应型的大豆品种很有可能含有和该鉴别寄主相同的抗病基因或2个鉴别寄主携带的抗病基因组合; 若待鉴定品种的反应型完全不同于14个鉴别寄主所含抗病基因或基因组合的反应型, 则这些品种可能携带未知的新基因或基因组合。
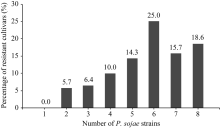
在黄淮海地区的140个大豆主栽品种中, 共有134个对1~8个菌株表现出抗性, 占鉴定总数的95.7%; 6个对8个菌株均无抗性。对不同菌株表现抗性的品种比例见图1, 其中, 对PsRace5菌株表现抗性的比例最高, 为82.1%; 抗PsRace1菌株的品种最少, 为54.3%。对不同数目疫霉菌株表现抗性的抗病品种所占比例见图2, 其中26个品种抗8个菌株, 占鉴定总数的18.6%; 22个品种抗7个菌株, 占15.7%; 35个品种抗6个菌株, 占25.0%。
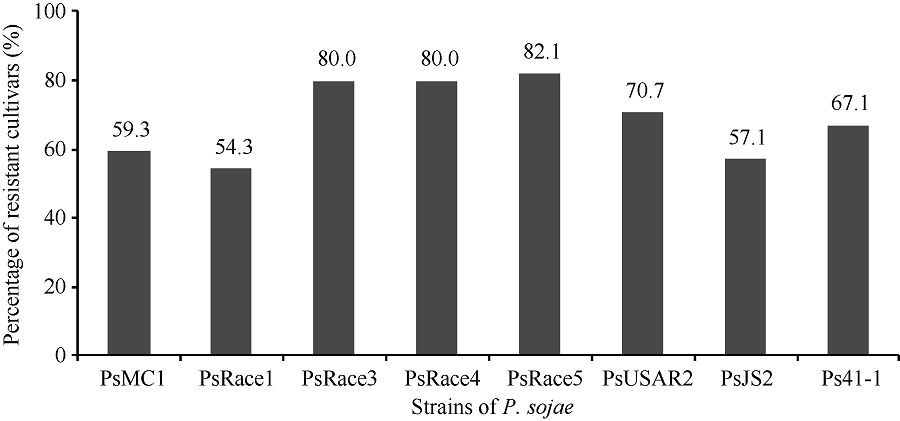 | 图1 抗不同大豆疫霉菌株的品种数量分布比例Fig. 1 Distribution of the number of cultivars resistant to different strains of P. sojae |
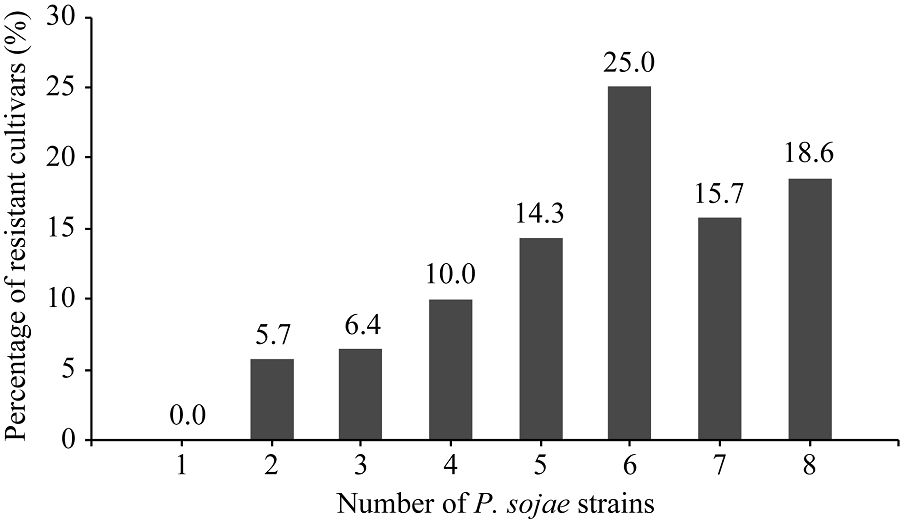 | 图2 供试大豆品种抗大豆疫霉菌株数目的分布Fig. 2 Distribution of cultivars resistant to different numbers of P. sojae strains |
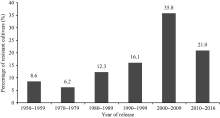
黄淮海地区不同年代大豆主栽品种对疫霉菌不同菌株抗性的比例有一定变化。图3显示, 20世纪80年代以后审定的品种对PsMC1菌株的抗性比例相对较高; 20世纪80年代审定的主栽品种对PsRace1全部表现为抗病; 1980— 2009年审定的主栽品种对PsJS2菌株抗病比例最高; 1970— 1999年审定的主栽品种对PsRace3、PsRace4和PsRace5的抗性比例较高, 均达到80.0%以上。从主栽品种对疫霉菌抗性的变化趋势可以看出, 尽管不同年代大豆主栽品种抗性比例有差别, 但每个年代均存在抗大豆疫霉根腐病的品种。抗6个以上菌株的品种共83个, 占鉴定品种总数的59.3%, 其在不同年代抗病品种中的分布比例如图4。可以看出, 20世纪70年代多抗品种比例相对较低, 随后不断增多, 在2000— 2009年间多抗品种比例高, 2010年后则有所下降。
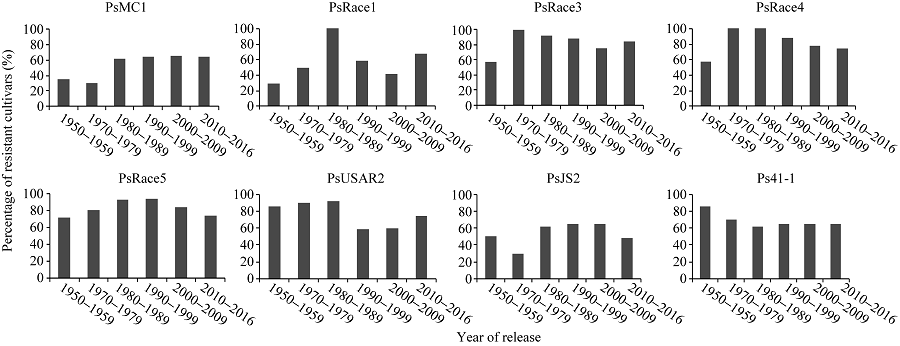 | 图3 不同年代大豆主栽品种对8个疫霉菌株的抗性比例Fig. 3 Percentage of soybean cultivars resistant to eight P. sojae strains in different years |
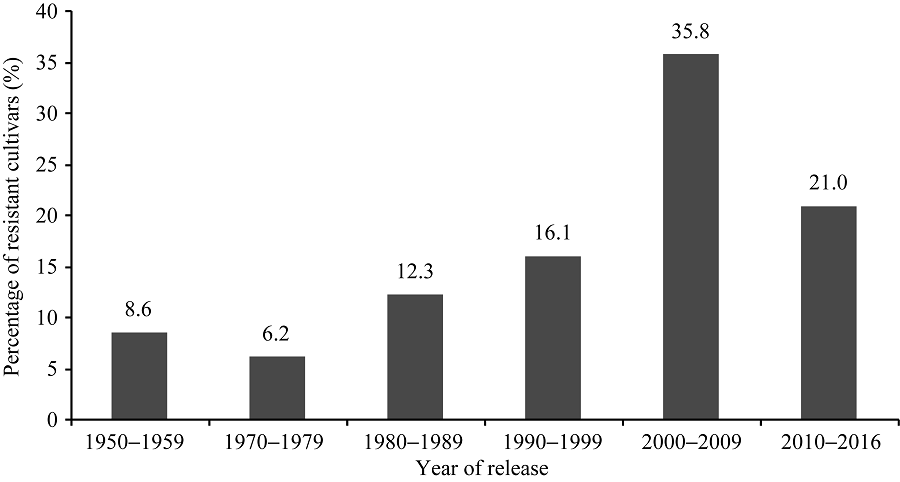 | 图4 抗6个以上菌株的品种在不同年代的分布比例Fig. 4 Distribution of soybean cultivars resistant to more than six P. sojae strains and released in different years |
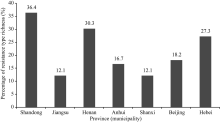
在所鉴定的大豆品种中, 来源于山东的品种所含抗病反应类型最丰富, 其次为来自河南、河北、北京、安徽、江苏和山西的品种。供试33个山东大豆品种表现出24种不同的反应型, 占总反应型(66种)的36.4%; 30个河南大豆品种表现出20种不同的反应型, 占30.3%; 25个河北大豆品种有18种不同的反应型, 占27.3%; 14个北京大豆品种有12种不同的反应型, 占18.2%; 16个安徽大豆品种有11种不同的反应型, 占16.7%; 11个江苏大豆品种有8种不同的反应型, 占12.1%; 11个山西大豆品种有8种不同的反应型, 占12.1% (图5)。对不同抗病反应型在各省(市)的分布情况比较后发现, 反应型为RRRRRRRR的大豆品种在山东、江苏、河南、安徽、山西和河北均有存在, 但部分反应型仅出现在某一个省份, 如RSRSSSSS反应型仅存在于山东大豆品种中; RRRRRSSR反应型仅存在于北京大豆品种中; SSRRSRSS反应型仅存在于河南大豆品种中; SSRRSRRR反应型仅存在于河北大豆品种中; RSRSRRRR反应型仅存在于山西大豆品种中; SSRRRRRR反应型仅存在于安徽大豆品种中; RRRRRSRR反应型仅存在于江苏的大豆品种中。由此可见, 黄淮海地区不同省(市)的大豆品种对疫霉菌的抗性水平存在明显差异, 在大豆抗病育种中, 应重视选用不同来源的亲本。由于本研究中所利用的品种数量有限, 对各省(市)品种抗病反应型的分析仍需要进一步研究。
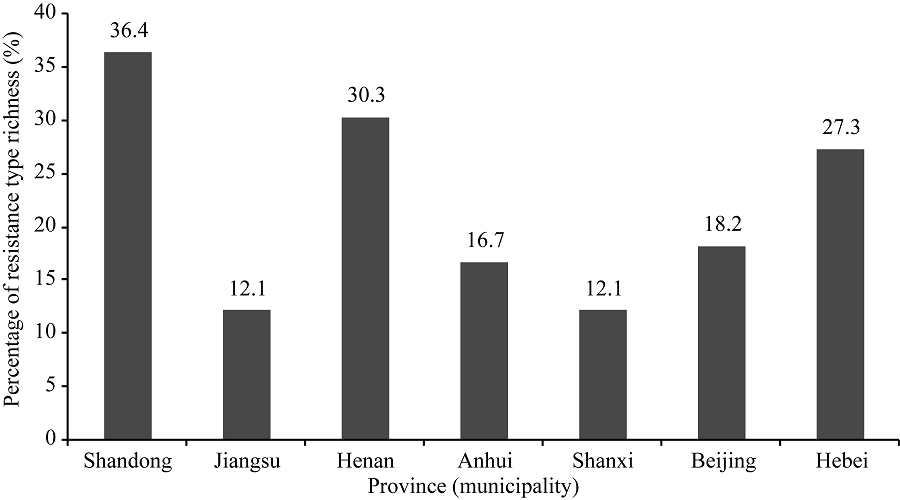 | 图5 7省(市)抗大豆疫霉根腐病的大豆品种分布情况Fig. 5 Distribution of resistance types of soybean cultivars to P. sojae in seven provinces (municipality) |
14个分别抗不同大豆疫霉菌株的大豆品种及对照感病品种Williams对8个菌株的反应型见表2。在140个主栽品种中, 郑120、邳县软条枝、徐豆12、中黄70、中黄79和邯豆5号对8个疫霉菌株的反应型表现为SSSSSSSS, 与感病对照品种Williams (rps)的反应型一致, 说明这些品种不含已知抗疫霉根腐病基因。有134个品种分别抗1~8个菌株, 共产生65种不同的反应型, 其中有19个品种产生的5种反应型符合已知单基因、2个单基因组合的反应型, 115个品种产生的60种反应型与含有已知单基因或2个单基因组合的反应型不同, 说明这些品种可能含有新的抗病基因或基因组合, 其中大豆疫霉菌株PsJS2能够击败所有已知抗病基因, 抗PsJS2的品种应该含有新抗病基因。
从不同年代大豆主栽品种抗病基因型的推导结果可知, 20世纪50年代审定的14个品种共产生12种反应型, 可能含有4个抗病基因Rps1c、Rps1k、Rps6或Rps8; 1970— 1979年审定的10个品种共产生10种反应型, 可能含有4个抗病基因Rps1c、Rps1k、Rps3a或Rps4; 20世纪80年代审定的13个大豆品种共产生9种反应型, 可能含有6个抗病基因Rps1c、Rps1k、Rps3a、Rps4、Rps6或Rps8; 20世纪90年代审定的17个品种共产生15种反应, 可能含有8个抗病基因Rps1b、Rps1c、Rps1k、Rps2、Rps3c、Rps5、Rps6或Rps8; 2000— 2010年审定的55个品种共产生36种反应型, 推断可能含有6个抗病基因Rps1c、Rps1k、Rps3a、Rps4、Rps6或Rps8; 2010— 2016年审定的31个品种共产生17种反应型, 推断可能含有6个抗病基因Rps1c、Rps1k、Rps3a、Rps4、Rps6或Rps8, 由此可知, 每个年代均有含抗病基因Rps1c或Rps1k的材料。从黄淮海地区7省(市)大豆品种所含抗病基因类型(表3)可知, 山东和河南大豆主栽品种含有已知抗病基因类型最多, 其次为江苏、河北、山西和北京的品种。
| 表3 7省(市)大豆主栽品种所含已知抗性基因的数量和类型 Table 3 Postulated number and types of known resistant genes of soybean cultivars from seven provinces (municipality) |
我国是大豆起源地, 拥有丰富的大豆种质资源。国内外研究学者已对我国大豆种质进行了疫霉根腐病抗性鉴定。孙石等[30]利用7个具有不同毒力公式的大豆疫霉菌株, 对黄淮海大豆主产区的96个大豆品种(系)进行接种鉴定, 发现黄淮海地区大豆品种对大豆疫霉根腐病具有丰富的抗性多样性; Lohnes等[31]用4个不同毒力型的生理小种接种726份大豆资源, 发现安徽、江苏、山东等地大豆品种抗疫霉根腐病的比例较高; 朱振东等[32]利用10个具有不同毒力型的大豆疫霉菌株对120个栽培大豆品种(系)进行抗性鉴定, 发现河南、安徽和山西等地大豆品种(系)抗性资源丰富。任海龙等[33]利用大豆疫霉菌株Pm14接种鉴定江西、湖南和广西273份野生大豆资源的抗病性, 发现54份抗病或中间类型的资源, 占供试材料的19.8%; 程艳波等[34]利用7个具有不同毒力型的疫霉菌株接种鉴定华南地区67个大豆品种(系)及其亲本的抗病性, 发现抗疫霉菌株PGD1的比例最高, 占鉴定品种总数的9.0%; 申宏波等[35]利用大豆疫霉菌1号生理小种对黑龙江省346份大豆种质资源进行抗性鉴定显示, 表现为抗病和中间类型的品种有27个, 仅占供试材料总数的7.8%。本研究利用8个不同毒力型的疫霉菌株接种黄淮海地区自20世纪50年代以来审定的140个大豆主栽品种, 发现其中95.7%品种表现抗性, 结果再次证明黄淮海地区大豆品种对抗疫霉根腐病存在丰富的抗性多样性。
基因推导是分析寄主抗病基因的一种简单、快速的方法。本研究通过与14个鉴别寄主品种对8个鉴别菌株的反应型相比较, 发现供试品种在对疫霉根腐病的抗性方面表现出66种不同的反应型, 其中有5种反应型符合已知单基因、2个单基因组合的反应型, 相关主栽品种可能含有8个抗疫霉根腐病基因Rps1b、Rps1c、Rps1k、Rps2、Rps3c、Rps5、Rps6和Rps8。在我国已定位和鉴定的11个抗病基因中就有5个是从黄淮海大豆主栽品种中发现的, 其中RpsYu25、Rps9、Rps10、RpsZS18和RpsYB30分别来源于豫豆25、鲁豆4号、皖豆15、早熟18和诱变30[10, 11, 12, 13, 14, 15, 17]。本研究对豫豆25、鲁豆4号和诱变30的抗病基因推导结果表明, 这3个品种可能含有新的抗病基因。基因推导方法虽然具有一定的局限性[36], 但是可利用此方法推导出品种可能含有的抗病基因, 然后进一步应用常规遗传学的方法和分子生物学技术进行基因鉴定和定位。事实也证明此种策略有效, 我国已定位和鉴定的11个大豆疫霉抗病基因都是采取这种策略获得的。所以基因推导法在大豆抗病材料筛选和和抗病育种中仍有重要价值。
根据不同地区疫霉菌株基因型有针对性地选育和利用抗病品种是防控疫霉病害的有效措施。在本研究所用的8个疫霉菌株中, PsMC1和PsJS2是从黄淮海地区分离到的, 主栽品种对这2个菌株的抗性频率分别为59.3%和57.1%, 而14个鉴别品种对PsJS2均感病。PsRace3是分离自东北地区的大豆疫霉菌株, 河南省地区大豆品种抗PsRace3的比例高达90.0%。唐庆华等[37]对来自全国8省(市)的115个大豆疫霉分离物进行毒力测定发现, 含有抗病基因Rps1a、Rps1c和Rps1k的鉴别寄主可以抵抗90.0%以上大豆疫霉分离物的侵染, 因此, 提出在中国种植含有Rps1a、Rps1c和Rps1k的大豆品种将会有效地降低疫霉根腐病的发生。在本研究中, 推导出8个品种可能含有Rps1c或Rps1k, 其中兖黄1号、郑州135、文丰7号、豫豆8号、郑9007五个品种同时抗6个大豆疫霉菌株, 而海阳爬蔓青、徐豆9号和邯豆8号3个品种同时抗7个疫霉菌株, 可以利用这些品种和产量高、综合性状好的主栽品种配制杂交组合, 也可以利用这些品种与含有其他抗性基因的品种杂交, 进一步聚合多个抗性基因, 培育高产、多抗的大豆品种。
在黄淮海地区不同年代审定的140个大豆主栽品种中, 6个品种对8个大豆疫霉菌株均无抗性, 134个品种分别抗1~8个大豆疫霉菌株。19个品种产生5种反应型, 可能含有单基因或2个单基因组合, 其中8个品种可能含有抗病基因Rps1c或Rps1k, 这些品种在大豆抗疫霉根腐病育种中具有重大价值, 是培育多抗品种的基因资源。
致谢: 中国农业科学院作物科学研究所朱振东研究员提供大豆疫霉菌菌株, 并在试验和论文撰写中提供指导, 谨致谢意。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|