{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
作物驯化和品种改良所选择的关键基因及其特点
[张学勇1, *  , 马琳
, 马琳1 , 郑军2 ]
, 马琳|
|


近15~20年作物基因组学迅速发展, 特别是第2代测序技术的普及, 显著降低了测序成本, 使单核苷酸多态性(SNP)分析和单元型区段(也称单倍型区段)分析渗透到生命科学的各个领域, 对系统生物学、遗传学、种质资源学和育种学影响最为深刻, 使其进入基因组学的全新时代。一批驯化选择基因的克隆, 特别是对一些控制复杂性状形成的遗传基础及其调控机制的解析, 更清晰地揭示了作物驯化和品种改良的历史, 提升了人们对育种的认知, 推动育种方法的改进。驯化和育种既有相似之处, 也存在明显的差异。驯化选择常常发生在少数关键基因或位点, 对基因的选择几乎是一步到位; 而现代作物育种虽然只有100年左右的历史, 但其对基因组影响更为强烈, 是一些重要代谢途径不断优化的过程。随着生态环境或栽培条件的变化, 育种选择目标基因(等位变异)会发生相应的变化或调整, 因此对基因(等位变异)的选择是逐步的。此外, 强烈的定向选择重塑了多倍体物种的基因组, 使其亚基因组与供体种基因组明显不同。在群体水平上, 系统分析驯化和育种在作物基因组和基因中留下的踪迹, 凝炼其中的规律, 将为品种改良和育种提供科学理论和指导, 本文也简要介绍了“十三五”国家重点研发计划专项“主要农作物优异种质资源形成与演化规律”的基本研究思路。
Crop genomics made great progress in last 15-20 years. Second generation sequencing technology has dramatically reduced the cost of genome sequencing, brought life science into the times of genomics, and strongly promoted development of system biology, genetics, breeding and genetic resources. Single nucleotide polymorphism (SNP) and haplotype block analysis are currently widely used for exploring animal and plant genetic resources and breeding. Successful isolation of many important genes helps us elucidate the history of crop domestication and breeding, and predict the future of breeding. It has been changing the breeding concepts and strategies. Most crop cultivars used today have experienced two major steps of harsh artificial selection, i.e., domestication and breeding. Despite so many similarities between domestication and breeding, they are different in some aspects. Domestication relatively affects small regions of plant genome. The dramatic reduction of diversity is usually caused by bottleneck effect. Although the breeding history is only about 100 years, it has brought tremendous alteration in most crop genomes. Breeding targets much more genes than domestication. It is very difficult for further selection of alleles at domestication targeted locus, usually only one allele is kept, which referred as fixed. However, it is not in the case of selecting alleles at breeding targeted locus. Few alleles can be present at the same time in cultivar populations and rotate at the same locus in varieties released in different periods or regions. Frequency of favored alleles in new cultivars has been increased dramatically because of positive selection. In addition, strong artificial selection usually reshapes the sub-genomes in polyploid species, which made them quite distinct from donor’s genomes. Therefore, it would be a good strategy to highlight future breeding through elucidating the basic rule of crop genome and gene in reaction to artificial selection at the targeted regions. Here, we briefly review the current major strategies for dissection of genes, haplotype blocks as well as the major genes targeted in crop domestication and breeding selection. We also give a brief introduction on the mission and strategies for “formation and evolution mechanism of funder genotypes and famous cultivars in major crops”, a newly initiated national key research and development program of China.
今天人们赖以生存的作物如水稻、小麦、玉米、大豆、棉花、油菜、蔬菜、果树等, 基本都经历了两次强烈的人工选择, 即驯化和育种。作物驯化和育种及基因组学的相关基础研究, 不仅有助于揭示在人工选择作用下生物遗传变异的规律以及物种形成的机制, 还能够加深人们对特定生物性状的起源、变异和进化的认识, 为进一步开发新的种质资源、更有效地改良品种提供理论基础和指导[1, 2, 3]。
驯化前作物野生种主要依靠群体内的多样性及个体遗传适应机制在自然界生存和繁衍。驯化的过程实际是作物的某些性状随着人们的意愿而改变, 是两者相互依存度不断强化的过程。在人工干预下, 受驯化的植物逐渐失去其野生祖先的部分生理、形态和遗传特性, 而人们需要的性状不断得到积累和加强。作物驯化的基本特征性状是从落粒到不落粒、由匍匐转向直立、分枝减少、休眠减弱、开花、成熟趋于一致, 果实大而均匀, 苦、涩味降低, 绝大部分栽培种在自然状态下的自我繁殖与传播能力降低, 须依靠人类的播种、管理、收获、留种才能有效繁衍后代。大量研究表明这些驯化相关性状大多为单基因或寡基因控制的质量性状。
由野生种驯化到栽培种的过程中往往改变了多个性状, 不同的性状在驯化时间上有先后之分。例如水稻由匍匐转向直立、由落粒成为非落粒应该是最早驯化的性状, 而籽粒由红色变成白色及糯性等应是比较晚一些的驯化性状, 关键的性状(基因)驯化选择完成后, 可能传播到周边或更远的地方, 完成二级性状的驯化选择。因此, 仅仅依据个别二级驯化性状基因的分布, 判断一个国家或地区就是某种作物的起源地、特别是初生起源地并不客观[4]。
一些多倍体物种如小麦、油菜、棉花等, 在不同的倍性水平都有驯化选择发生, 即驯化与多倍体化相伴而行, 例如小麦在四倍体水平已基本完成了驯化, 形成了多个栽培种, 而六倍体化后的新物种又具有野生或半野生性状, 经再次驯化后成为今天的普通小麦。人工合成小麦基本不能直接应用于生产, 未完成二次驯化可能是重要原因之一。芸薹属甘蓝型油菜的形成过程与小麦类似。
驯化过程中作物形态、生理和生存环境发生了改变, 究其根本是物种遗传基础发生了改变。20世纪50年代以前, 科学家们对于进化的研究主要集中在古生物学、分类学、胚胎发生学、比较解剖学、生物地理学、生理学、群体遗传学等方面。此后, 随着分子生物学和分子遗传学的兴起, 以及核苷酸和氨基酸序列数据的累积, 进化学家逐渐开始从分子水平研究生物进化的原理和机制, 分子进化也逐渐成为研究进化的主要手段。大量研究表明, 由于人工选择的作用, 致使栽培种与野生种之间, 现代品种与古老的地方品种之间在群体遗传结构及性状上形成了很大的差异[8, 9]。遗传多样性显著降低是驯化的一个重要标志, 即所谓的驯化瓶颈[2, 8](domestication bottleneck)。由于驯化过程中, 野生群体中只有少部分个体被采集, 用于人工种植和驯化, 驯化群体的种群数量较野生群体急剧减少, 基因组水平的多样性大大降低, 而承受选择的基因在群体中的多样性降低则更为明显, 同时这些基因附近区域的遗传多样性也随之下降。遗传学中把这种对少数基因选择导致侧翼区域遗传多样性降低的现象称为选择牵连效应(hitchhiking effect), 亦称选择谷(selection sweep)[10]。此外, 自然选择、群体扩张或分化等都可能作用于驯化物种并在基因组中留下“ 印迹” 。了解这些因素对驯化物种的作用和影响程度, 对了解驯化物种的进化历史具有重要意义[11]。
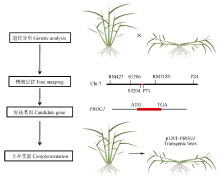
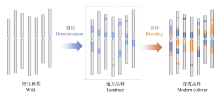
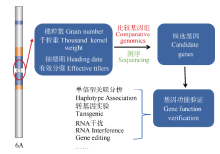
驯化和育种中对基因的遗传学研究主要有两种方法: 一是将栽培种与野生种杂交、回交, 建立重组自交系或导入系, 针对驯化性状开展QTL定位及克隆分析, 研究驯化基因结构特点、等位变异及驯化的机制(图1)[12, 13]; 二是对栽培种(特别是古老的地方品种)与野生种、半野生种进行基因组重测序分析, 通过选择谷作图(selection sweep mapping), 确定受选择的基因组区段和基因(图2和图3)。例如, Zhou等[7]通过重测序, 系统分析水稻著名品种“ 黄华占” 的形成过程, 发现对其形成贡献最大的是另一个著名品种“ 特青” , 由此提出保守传递的基因组区段通常含有对产量、适应性有重要作用的基因; Xie等[14]对1496个品种(系)重测序后发现约7.8%的基因组区域在育种中受到比较强烈的选择, 品种(系)产量的高低与其拥有正向选择区段的数量正相关。
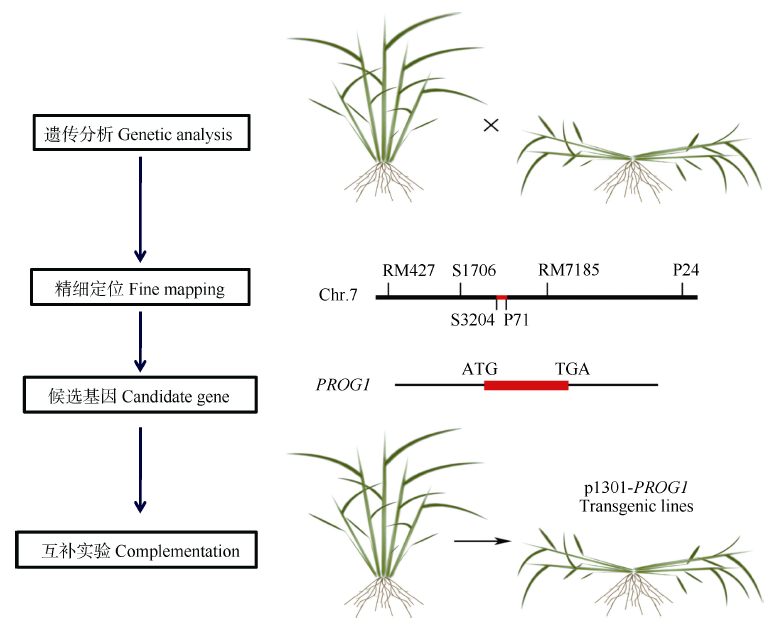 | 图1 基于基因图位克隆, 对驯化和育种中选择基因的系统遗传学分析[12, 13]Fig. 1 Map-based clone and function certification of genes targeted in domestication and breeding[12, 13] |
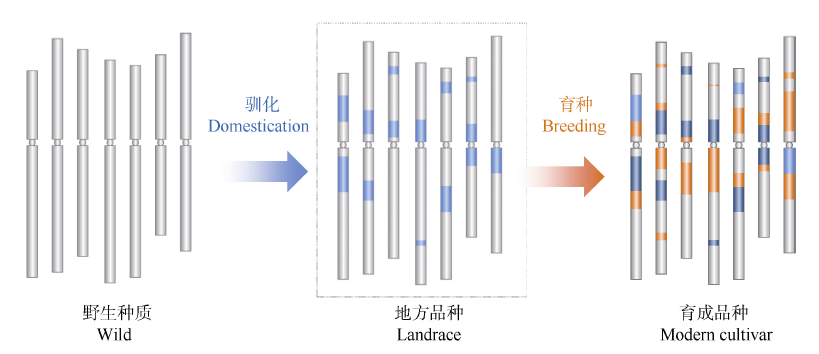 | 图2 基于选择牵连效应对驯化和育种选择基因组区段的分析(以小麦A基因组为例) 浅蓝色: 驯化选择区段; 深蓝色: 驯化和育种共同选择区段; 橘黄色: 育种选择区段。受选择的区段DNA序列的多样性显著降低。Fig. 2 Identification of haplotype blocks formed in domestication and breeding based on genome resequence and selection sweep analysis (using the A genome of wheat as a model) Bright blue: domestication haplotype blocks; Dark blue:haplotype blocks extensively selected from domestication to breeding; Orange: breeding haplotype blocks. Diversity was dramatically declined at the genomic regions targeted in both domestication and breeding. |
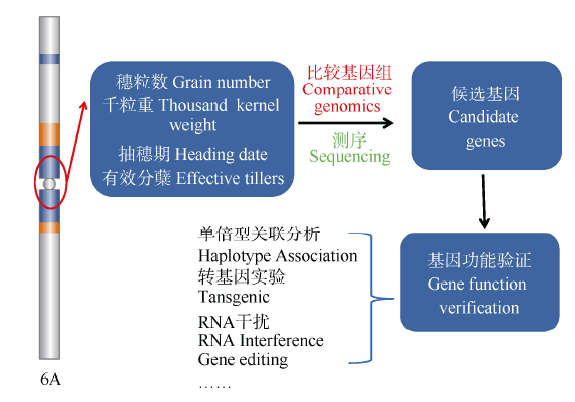 | 图3 以单元型区段的关联分析为基础, 通过测序和基因组比较分析, 进行关键基因的确认(以小麦6A染色体为例)Fig. 3 Genetic dissection and function verification of the causing genes at the crucial blocks (using wheat 6A as a model) |
驯化性状一般是从野生种到栽培种过渡过程中最早选择的性状, 如休眠的弱化、果实或种子的变大, 成熟断穗、落粒特性的丧失, 植株个体形态(株型)的改变等都属于典型的驯化性状。驯化基因一般具备3个特点, 即与驯化性状之间的相关性明确; 在驯化过程中受到很强的正向选择; 在由一个群体演化而成的不同种群中, 几乎完全丧失了多样性(固定)[9]。
大量驯化基因的研究结果表明, 野生性状的丢失和栽培性状的获得往往是野生个体的基因发生突变, 基因功能丧失、歧化或是新功能获得后产生的结果。这些性状之所以被选择, 是因为它们符合早期人类的需要, 在自然环境中驯化后的个体是很难生存的, 因此, 驯化选择的方向与自然选择方向往往是相矛盾的。从基因本身来看, 与驯化密切相关的基因大多为转录因子和激酶调控过程中的关键基因, 这类基因在生长、发育、结实和成熟过程中起着重要调控作用。一些重要代谢途径上关键节点的催化酶编码基因也常常是驯化的目标基因, 如谷类作物的淀粉合成途径、氮吸收利用途径、葫芦科的葫芦素合成途径等[8, 9, 15, 16]。
落粒是许多植物适应自然环境, 自我繁殖、保存种群的一个基础生物学特性, 却是作物栽培和生产中首先要克服的一个缺点。落粒性的丧失是谷类作物人工驯化过程中的一个关键事件, 是谷类作物驯化的首要性状, 也是驯化研究的典型性状。科学家在日本晴与Kasalch人工作图群体中精细定位控制水稻落粒性的主效QTL并克隆了目的基因qSH1, 其编码一个BEL1型同源异型蛋白, 5' 端调节区的一个单碱基突变(SNP)引起离层不能形成而丧失落粒性[17, 18]。余建民小组对落粒基因在谷类作物中的平行进化进行了系统分析, 发现水稻中的Sh4和Sh1在高粱中仍然控制着落粒性, 说明一些重要的与进化相关的基因在物种间功能趋于保守[19]。但相关机制在物种间也存在差异, 例如大麦和小麦在禾本科中遗传关系较近, 然而控制断穗的基因并不是同源基因。
小麦方穗基因(Q/q)作为小麦驯化的关键基因(位于小麦5A染色体上, 5B和5D上各有一个功能较弱的同源基因), 是一个AP2转录因子, 是影响穗型(长方穗/尖穗)的关键基因, 也是影响裸粒性的重要基因, 并存在明显的剂量效应, 其中一个氨基酸的改变使小麦由尖穗(拟斯卑尔脱型)转变为长方穗, 非裸转变为裸粒, 给收获和打碾带来极大的方便, 推动了小麦在全球的迅速扩张, 促进了农耕文明的发展[20]。在相当长的时期内, 大多数科学家认为就某个单一性状而言, 驯化往往只发生在一个地区, 这就为作物的驯化起源地的争论埋下了隐患。大麦成熟时断穗与否由位于3H染色体上紧密连锁的2个基因Btr1和Btr2同时控制, 其中任何一个基因发生隐性缺失突变, 都可使大麦由断穗(野生)成为非断穗(栽培), 基因单元型分析显示btr1突变发生在中东地区南部, 而btr2发生在北部[21]。可见一种作物伴随着已有栽培种的扩散、与野生祖先种之间的基因渗透, 以及二次驯化等原因, 可能形成多个起源或驯化的地区, 也可能形成数个次生多样性中心, 例如小麦起源于中东, 但我国是很重要的次生多样性中心, 多个重要基因是在我国发现的, 如“ 大拇指矮” (Rht3), 太谷核不育基因(Ms2), 苏麦3号(FHB1)及糯性(Wax)基因的部分重要等位变异等。
Doebley等[8]、Studer等[22]和Zhou等[23]将玉米与大刍草杂交、回交, 发现从大刍草向玉米过渡的关键基因tb1 (分枝/单秆), 玉米在该基因的上游64 kb附近一个转座子(hopscotch)的插入, 提高了基因的表达量, 使个体的顶端优势更加明显, 从而抑制了分枝的产生, 这是转座子调控基因表达影响物种进化的直接证据。
Tan等[13]在普通野生稻(沅江野生稻)导入系后代群体中精细定位并克隆了控制匍匐/直立的关键基因(OsPROG1, 图1), 该基因编码锌指蛋白, 被定位于细胞核中, 是典型的转录因子, 栽培稻中该基因突变后丧失功能, 使植株从匍匐转向直立, 提高了单位面积穗容量, 同时也提高了穗粒数, 是水稻驯化的关键基因。Jin等[12]也对该基因进行了系统的功能研究, 发现该基因主要在叶腋中表达, 促进大量分蘖的形成, 在栽培稻中发生氨基酸替换, 基因的功能减弱, 分蘖减少, 植株直立。
从蔓生无限结荚转变为直立有限(或亚有限)结荚是大豆驯化的最重要性状之一。Tian等[24]和Ping等[25]发现有限和无限结荚习性受2个互作的基因Dt1和Dt2同时控制, Dt1 (GmTfl1)编码一种信号蛋白, 主要在顶端分生组织中表达, 其内部4种非同义突变是导致大豆结荚习性发生改变的主要因素, 在不同生态区, 遗传类型明显不同, 有限习性主要分布在南方, 而无限习性主要分布在北方, 2种单元型近似均等的分布在黄淮流域, Dt1表达越强, 无限特性越突出, Dt2为MADS类转录因子, 对Dt1的表达有一定的抑制作用, 促进大豆由营养生长向生殖生长的转变。因大豆的结荚习性由Dt1和Dt2位点的等位变异共同决定, 所以形成比较丰富的中间过渡类型, 满足不同生态环境和栽培模式需求。
果实大小、数量及其色泽也是重要的驯化性状, 因其直接关系到产量及商品性。水稻单株穗粒数、粒重的研究引领着谷类作物产量性状的遗传解析。GS3、GS5、GW5、GW8、GIF1在水稻驯化过程中受到比较明显的选择[26]。GW2、qGL3.1和TGW6虽然遗传效应不小, 但在水稻驯化和育种中并未受到很强的选择, 可能是普通野生稻群体中优异等位变异频率太低造成。在小麦籽粒性状研究过程中TaGS5-3A受到比较强烈的选择瓶颈效应[27]。我们在对小麦淀粉合成途径关键基因的遗传效应分析中, 发现蔗糖合酶基因(TaSUS1-7A)在小麦多倍化及驯化过程中受到强烈选择, 其酶活性与千粒重密切相关[15]。
Lin等[6]和Cong等[28]通过基因组重测序结合SNP分析, 发现番茄从野生种到现代加工型品种, 浆果平均重量大幅度提高是经过2个大的历史阶段完成的, 第一阶段(驯化)在番茄基因组中186个区段出现选择谷, 选择的主要基因有fw1.1、fw5.2、fw7.2、fw12.1、icn12.1; 第二阶段(现代育种)在133个区段形成选择谷, 选择的主要目标基因有fw2.1、fw2.2、fw2.3、fw11.1、fw11.2、fw11.3、inc2.2等13个, 第一阶段所形成的186个选择区段中有21%在第二阶段继续受到选择。
谷类作物籽粒中的淀粉、蛋白和脂肪是人类的主要能源和营养源。不同国家或地区的居民因生活习惯及饮食偏好, 在作物的驯化过程中留下了印迹。最典型的例子就是对食物蒸煮后的黏性(糯性)的驯化。我国南方及周边的泰国、越南、柬埔寨等国家, 居民喜欢吃糯米食物, 如糍粑、年糕等, 蒸煮后相对较为黏稠的品种成为农民稻田中的所爱, 早期的选择留下许多糯性的地方品种, 这些材料共同的遗传特点是籽粒淀粉合成途径上直链淀粉合成基因(Wx)发生突变, 其功能弱化或受阻, 使籽粒中支链淀粉的含量提高, 而直链淀粉含量显著下降。小麦、玉米、高粱、糜子、谷子、马铃薯、甘薯中该基因发生了许多自然突变, 并产生了糯性品种, 是早期农民选择的重要性状。此外, 淀粉的合成与谷类作物、块根、块茎类作物的收获器官大小、淀粉含量及直链/支链比例相关, 在早期的驯化过程中淀粉合成途径上的一些关键酶编码基因普遍经历了比较强烈的选择, 是重要的驯化间接选择基因[15, 29]。“ 糯性淀粉” 是谷类、块根、块茎类作物中共同选择、受同一基因控制的性状, 是平行进化的范例。
苦涩味的降低或消除既是重要的驯化性状, 也是重要的育种性状。一个很有启发性的研究是黄三文课题组对黄瓜苦味形成基因网络的研究。野生黄瓜叶片、卷须及果实中都含有苦味物质葫芦素, 可能与抵御昆虫的侵袭密切相关, 而叶片、卷须和果实中苦味物质由同一代谢途径合成, 其受组织特异的转录因子调控(Bl、Bt分别调控叶及果实中苦味物质合成基因的表达), 这些基因在第1、第3和第6染色体上呈现相邻分布的特点, 并受Bl或Bt的共调控。早期驯化选择主要发生在Bt转录因子上, 其中个别碱基的改变降低了其对果实中苦味物质合成的正向调控作用, 果实苦味基本消除, 但叶片和卷须仍然是苦的。此类化合物合成途径上关键基因Bi表达活性的降低, 使叶片、瓜蒌、果实均失去苦味, 该基因在现代育种中也受到强烈正向选择[16]。
矿质营养的吸收和利用是作物起源和进化研究中常被忽视的性状, 比如对普通野生稻氨离子(NH4+)转运蛋白编码基因(OsAMT1; 1)分析后发现24种单元型, 而在栽培稻中仅保留了4种单元型, 其中高效转运的Hap-1占栽培品种的96.3%, 对该基因的选择导致其上、下游150 kb内21个基因也丧失了多样性, 毫无疑问, OsAMT1; 1是一个重要的驯化基因, 并在育种中继续受到选择[30]。因此, 一些重要营养元素的吸收和代谢可能是重要的驯化目标性状, 相关基因在今后的研究中应引起重视。
对低温的耐性是水稻北扩的重要限制因子, 也是粳稻和籼稻地理分布差异的遗传基础。我国北方的水稻全是粳稻, 而南方以籼稻为主, 长江流域则是籼、粳共存。针对水稻感知温度的分子机制, Ma等[31]研究发现, COLD1基因合成一种蛋白, 通过与细胞膜上G-蛋白互作, 参与细胞内钙离子流的调控, 过量表达COLD1可以显著提高水稻的耐冷性, 而抑制其表达, 则显著降低耐冷性。该基因第4外显子一个碱基的改变, 导致第187位氨基酸由Met/Thr (甲硫氨酸/苏氨酸)变成Lys (赖氨酸), 使得该蛋白对钙离子流的激发能力发生变化, 引起抗冷性的差异。所有抗冷的粳稻品种均保留了这一新的等位变异, 同时在我国的普通野生稻中也发现了相同的变异类型, 因此认为COLD1也是粳稻驯化的重要基因。
相对而言, 驯化对基因组的影响是局部的, 基因组多样性的显著降低多与瓶颈效应密不可分, 而育种对基因组的影响范围更宽、更广, 对基因组整体多样性的冲击更大。育种目标主要包括高产、优质、抗倒伏、适应性、抗病以及抗逆等性状, 每一类性状选择基因是不同的, 但其又是互相依存的[32], 为便于行文, 我们仍然以性状为基础, 概述相关基因研究进展。
水稻基因组测序的完成极大地推动了水稻乃至小麦和玉米产量性状的基因解析, 科研人员对控制水稻分蘖(MOC1等)、穗粒数(Gln1)、粒重(GW, GS)等进行了深入的研究。到目前为止, 从水稻中克隆的多个与产量密切相关的基因, 基本可以归纳为与植物激素代谢相关的基因, 与蛋白体降解相关基因(泛素代谢途径), 细胞分裂增殖调控基因, G-蛋白信号途径, 淀粉、蛋白、油脂合成途径等[26, 33]。这些发现为建立水稻复杂农艺性状的分子调控网络以及新一代的超级水稻分子设计育种提供了重要理论依据。
4.1.1 控制分蘖的基因 分蘖是构成产量的基本要素之一, 主要受植物体内激素分布和环境因素的控制。生长素、细胞分裂素、独脚金内酯和油菜素内酯均是影响分蘖的主要激素。水稻中已克隆出参与生长素合成和运输的YUC和OsPIN1等基因[34], OsIPT是调控细胞分裂素合成的关键基因[35]。参与独脚金内酯合成与信号转导的D3、D7、D10、D14、D27等基因也陆续报道与分蘖能力密切相关[36, 37]。MOC1是调控分蘖芽生长发育的主效因子, 对其系统研究揭示了其参与细胞周期调控、影响分蘖以及株型建成的机制[38]。此外, Wang等[39]报道了与水稻顶端优势相关的DWT1基因, 通过影响一个未知的信号分子在穗部的合成或运输, 调节细胞分裂素的稳态以及茎、节对赤霉素的响应能力, 从而控制主茎和分蘖茎节的伸长。水稻中分蘖基因及其主要代谢途径的发现, 必将推动对麦类以及其他作物分蘖性状的解析和认识。
4.1.2 穗粒数控制基因 穗粒数的遗传力较分蘖高, 增加每穗粒数是提高产量的重要途径。控制穗粒数的基因大致可分为两类, 一类与抽穗期相关, 并影响每穗颖花数, 一般具有多效性, 同时控制抽穗、株型和每穗颖花数等性状, 如Ghd7和Ghd8控制穗粒数、抽穗期和株高3个性状[40]。另一类则主要参与穗分化过程, 通过调控分化速率和分化方向影响穗粒数。
水稻细胞分裂素氧化酶编码基因(OsCKX2)是作物中最早发现影响穗粒数的关键基因, 在水稻中通过控制一、二次枝梗的数量对结实数起到决定作用[41]; Li等[42]发现锌指蛋白转录因子DST (drought and salt tolerance)直接调控生殖器官生长锥中细胞分裂素氧化酶基因的表达, 从而调控细胞分裂素在分生组织中的分布, 对颖花数和籽粒大小起调控作用。Liu等[43]将关联分析与图位克隆相结合, 克隆了主效QTLKRN4, 其位于SBP-box基因Unbranched3 (UB3)下游60 kb处, 是一个1.2 kb的转座子片段的插入/缺失, 该位点作为UB3的顺式调控因子, 调控UB3基因的表达量, 控制玉米穗行数的变异, UB3启动子区个别核苷酸与该重复序列对穗数(穗粒数)的效应存在明显的加性效应, KRN4在玉米驯化和育种中均受到强烈的选择。
Zheng等[44]发现mRNA转录延伸因子(TaTEF- 7A)表达量对小麦穗粒数和穗下节间长度有很强的影响, 所有变异都发生在启动子区。Liu等[45]发现生长素响应因子(ARF18)基因内一个165 bp的缺失与油菜果荚长度和重量的增加显著相关, 该缺失造成第6内含子非正确剪切, 导致ARF18不能形成蛋白同源二聚体, 丧失与DNA的结合能力, 解除了ARF18对生长素基因转录的抑制作用。可见, 影响穗粒数的关键基因多与细胞分裂素的合成、运输及响应密切相关。
4.1.3 粒重控制基因 籽粒的形成基本是由2个阶段组成, 即胚乳细胞分裂和胚乳细胞中淀粉、蛋白、油脂大量积累、脱水, 引起细胞程序化死亡, 形成胚乳。因此, 影响胚乳细胞分裂、储藏物质合成和积累的基因, 必然影响籽粒的大小和容重。左建儒和李家洋对影响谷类籽粒大小的基因及生化基础分析后认为, 植物激素(油菜素内酯、细胞分裂素、生长素等)代谢、蛋白降解(proteasomal degradation)、G-蛋白信号途径及一些影响细胞分裂的基因(GS5、GW8、IPA等)对籽粒大小起着决定作用[26, 27]。在三大谷类作物中, 籽粒淀粉合成途径的畅通与否与产量密切相关, 该途径上的主要基因既是驯化选择的关键基因, 也是育种选择的重要目标基因[15, 29, 33]。我们对近十个影响小麦千粒重的基因在自然群体中变异及其遗传效应分析, 发现绝大多数变异发生在启动子和内含子区, 编码区变异极少, 可见产量基因的变异大多为基因表达量“ 高与低” 的差异, 而非功能“ 有和无” 的差异。
优质是育种的另一个主要目标。水稻品质主要与粒型、长宽比及淀粉特性有关, 小麦品质主要与蛋白含量、组成(HMW-GS中半胱氨酸的数量)及直链淀粉/支链淀粉的比例有关, 油脂类作物则主要与其籽粒不饱和脂肪酸、芥酸、硫苷化合物含量有关。最近对甘蓝型油菜及祖先种的基因组测序给我们很大的启发。甘蓝型油菜作为一个新物种, 新品种与老品种及其祖先二倍体种相比, 由于对油分含量的不断选择, 其合成相关的基因拷贝数明显增多, 而硫代葡萄糖苷(芥子油苷)合成途径的基因有删除和功能歧化的现象, 在人工新合成的四倍体油菜品种中A、C基因组之间基因、甚至染色体片段的置换是很普遍的[46]。
植株矮化是灌溉面积不断扩大、化肥广泛应用、机械化程度不断提高的必然要求, 是20世纪60年代后作物品种改良中变化最大的表型性状, 在小麦和水稻中矮秆基因的广泛应用, 引发了第一次绿色革命。对矮秆基因研究发现其多与植物激素赤霉酸的合成、转运、氧化和降解相关。Peng等[47]首次克隆了小麦的半矮秆基因Rht-B1/Rht-D1及玉米的dwarf-8 (d8), 通称为GAI基因(GA-Insensitive), 发现N端(DELLA区)个别氨基酸的改变降低了与赤霉酸受体蛋白GID的结合能力, 从而减弱了细胞核对赤霉酸的敏感度, 降低了植株的高度, 系列等位基因致矮的效应与其DELLA区和GID1蛋白(GA-Insensitive Dwarf1)的结合力高度负相关, 结合能力越差, 致矮能力越强。同一位点复等位基因致矮能力的差异正好满足了农业生产不同时期(或阶段)对作物株高和抗倒伏要求的差异[48, 49]。近50年的小麦育种对Rht-B1/Rht-D1进行了较强的选择, 我国主产区品种24.3%携带Rht-B1b, 46.9%携带Rht- D1b[50]。水稻中来源于我国台湾地方品种“ 低脚乌尖” 和“ 矮脚南特” 的半矮秆基因sd1是GA20 氧化编码基因(GA20ox-2)突变, 导致功能丧失, 赤霉酸合成受阻所造成, sd1可能有多个不同的等位变异, 其致矮能力也存在比较大的差异[51]。在水稻和小麦中, 虽然发现和定位了多个致矮基因, 但真正用于育种和生产的则屈指可数, 其原因是矮秆基因的负作用或连锁累赘, 或许将来的品种间基因组比较分析能给出一些比较准确的解释。
良好的适应性是物种在自然界生存的前提。许多作物如水稻、小麦、玉米从原始的驯化起源地向新的区域不断传播和扩张, 一些新育成的品种跨纬度推广种植, 依赖于品种对光(光周期反应)、温(春化)的准确反应, 准时启动生殖发育相关基因的表达, 使营养生长转入生殖生长, 完成一次生命周期的循环。
在作物中CCT类转录因子是决定开花时间的关键基因, 其特点是通常基因的C-端具有CCT功能域, 而N-端具有锌指结构域, 其表达均具有昼夜节律, 受其他基因的调控, 是品种适应性改良的关键基因。Hd1 (heading date1)是水稻中第一个克隆出的CCT基因; Ghd7除影响水稻株高和穗长外, 在ELF (early flowering)家族基因的作用下参与水稻花期调控, 强表达的单元型主要分布在热带(海南)和亚热带地区, 而功能最弱的则分布在我国东北稻区, 而华中、华南早稻区则多为基因功能缺失类型[40]。OsCO3编码产物与Hd1相同, 通过剂量效应影响Hd1; OsCOL4可上调Hd1的表达水平, 使水稻在长日照和短日照条件下抽穗期延迟。近来2个驯化相关的CCT家族基因成员DTH2和Ehd4也被克隆, 通过与MADS-BOX转录因子作用在不同光照条件下调控抽穗期, 在水稻从南向北的扩张及品种改良中发挥了很重要的作用[52]。Yang等[53]发现在玉米驯化完成后, 一个CACTA转座子片段插入到ZmCCT的启动子区, 部分抑制了该基因的表达, 从而减弱了玉米对日照长度的敏感性, 为玉米向温带地区的扩张提供了基础, 而该转座子片段几乎在所有温带品种中得到保留。
与水稻和玉米相比, 麦类作物(小麦、大麦、黑麦)生命周期更长, 一生所经历的环境更为复杂和严酷, 一般需经过“ 秋-冬-春-夏” 四季, 方能完成一个生命周期。小麦有冬麦、春麦之分, 因此其对温度、光照的反应和调控机制, 可能是最为复杂和精致的。大部分小麦品种需经过春化(感受低温), 在长日照下抽穗开花。小麦对低温的反应由位于5A、5B和5D染色体上的Vrn基因共同作用所决定, 其中作用最强的2个基因分别命名为Vrn1和Vrn2。Vrn1编码AP1 (APETALA1)类转录因子, 对开花起促进作用, 而Vrn2编码ZCCT蛋白(CONSTANS、CONSTANS-like和TOC), 是另一类转录因子, 抑制Vrn1的转录(负调控), 因此一个品种的冬春性由Vrn1和Vrn22个基因的等位变异共同决定[54, 55]。低温抑制Vrn2的转录, 促进营养生长向生殖生长的转变。几乎所有的小麦、大麦、黑麦原始种都是冬性的, 但一些材料在Vrn1启动子区发生缺失后丧失与ZCCT蛋白的结合能力, 解除了ZCCT蛋白对Vrn1表达的抑制, 转变为春性, 相对野生型其为显性, 从而使得麦类作物有了明显的冬、春之分(表1)。当Vrn2发生突变(vrn2)时, 不能正常转录或翻译, 丧失了对vrn1的抑制, 呈现无需春化就可开花的特点(表现为春性)。因此, Vrn1和Vrn2任一位点的突变, 都可导致品种冬、春性的改变, 增加了变异的类型, 如果考虑5B、5D染色体上作用相对较弱的Vrn1和Vrn2基因的表达及互作, 则变异类型更为丰富, 这也在一定程度上说明了小麦的冬性、半冬性、弱冬性和春性的遗传基础。而大麦则是通过Vrn2位点ZCCT基因的一次重复和变异, 满足不同地区对冬春性要求的差异[56]。
| 表1 小麦5A染色体上Vrn1和Vrn2基因的互作与品种冬、春性之关系 Table 1 Gene interaction and genetic decision of winter or spring types of wheat cultivars by two major genes Vrn1and Vrn2 on 5A |
调节麦类作物开花的另一套关键基因是光周期基因(Ppd1), 也属于CCT基因家族。小麦中该基因的表达上调, 明显促进抽穗、开花, 在我国及全球一年两熟地区主要以对光照长度不敏感的类型为主, 春麦区主要是光敏感类型[57, 58]。
抗病、抗虫是保障作物稳产的基础, 是育种选择的重要目标性状。作物的抗病性可基本分为垂直抗性和水平抗性两大类。从最早克隆的番茄PTO基因、水稻Xa21等基因的氨基酸序列分析发现, 其共同的特点是富含亮氨酸(LRR)和核酸结合域(NBS)的激酶[59], 后续研究发现大部分抗病基因都存在多个类似拷贝的重复, 且常位于基因组的重组热区, 主要的变异常发生在编码区和启动子区[60], 这种变异方式可能与作物适应病原菌小种的快速进化有关, 与抗性的增强也有关联, 例如Liu等[61]发现水稻对褐飞虱的抗性强弱与几个类似抗病基因的重复密不可分; 大豆对包囊线虫的抗性基因虽不属于NBS-LRR类基因, 但其抗性与基因的重复有密切关系, 一个拷贝表现高度敏感, 而抗病品种普遍携带10个串联重复[62]。
2009年报道了小麦持久抗病基因Lr34, 不仅对叶锈多个小种有很好的抗性, 同时对条锈病(Yr18)、白粉病(Pm38)也有很好的抗性, 三种抗性实际是同一个基因控制, 其编码ATP-结合型转运复合体蛋白。Lr34在生产中已被应用近50年, 但仍保持着良好抗性, 一个明显的负作用是引起叶片、特别是旗叶叶尖坏死[63]。最近克隆的另一个小麦多抗基因Lr67 (对三锈、白粉均有不同程度的抗性)则是一个己糖跨膜转运蛋白[64]。因此, NBS-LRR类蛋白代表了一类重要的抗病因子, 但并非全部。
作物抗病育种中一个有趣的现象, 即来自野生种或近缘物种的抗病基因常表现出比较广谱、持久的抗性特点, 如从簇毛麦和十倍体长穗偃麦草转移到小麦的Pm21和Lr19以及从野生稻转移到水稻的Xa21/23等, 这些抗病基因的应用研究虽已开展, 但其持久、广谱的机制目前还不是十分清楚。
干旱和盐害是作物生产中面临的2个最主要的非生物限制因子。就耐旱而言, 是作物在干旱条件下表现出的综合性状, 虽然进行了很长时间的研究, 但在遗传学上仍未取得突破性的进展, 目前比较肯定的是植物的DREB转录因子的表达与耐旱关系密切, 特别是提高其表达量可以明显地促进小麦等作物根系的发育[65]。相比而言, 对作物耐盐机制及关键基因的研究最近几年则取得了比较大的进展。
HKT1 (K+离子高亲和性转蛋白)、SOS1 (盐敏感)及NHX (Na+/H+交换泵)共同决定细胞内Na+的平衡, HKT1和SOS1蛋白控制其跨质膜转运, 而NHX则控制Na+从细胞质向液胞的转移。在栽培一粒小麦中HKT1有2个拷贝, 即位于2A染色体上的HKT1; 4和5A染色体上的HKT1; 5, 但在四倍体的圆锥及硬粒小麦中, 这2个基因均已丢失, 造成四倍体种对盐极度敏感, 而D基因组的加入, 六倍体中2个基因的功能得到恢复(2D和5D染色体上的同源基因发挥作用), 与其四倍体祖先种相比, 普通小麦的抗盐能力得到显著提升[66]。Munns等[67]证明, 从二倍体祖先种向普通小麦中重新引入HKT1后, 六倍体小麦在盐碱地的产量显著提高。此外, D基因显著提升了小麦对光、温的适应范围, 为其在全球范围的广泛种植、成为第一大粮食作物奠定了基础[68, 69, 70]。
品种的改良不仅是对当时、当地(如抗病、矮秆、优质)一些所急需性状(基因)的强烈选择, 而是许多重要代谢途径基因、调控元件不断优化的过程[5, 6, 15]。作者课题组对小麦微核心种质群体分析后发现在影响小麦穗粒数或千粒重的位点上, 与地方品种相比, 现代品种拥有更多的优异等位变异, 穗粒数的多少或千粒重的高低与品种拥有优异等位变异的总数呈正相关[71, 72]。
产量基因多与植物激素代谢、细胞分裂、淀粉和油脂的合成相关, 而矮秆基因多与赤霉素代谢有关, 适应性则与光、温感受基因CCT等相关, 抗病虫能力NBS-LRR基因与其重复密不可分, 而耐逆性(盐、旱)多与膜(细胞膜、液泡膜)上的转运蛋白有关。从遗传学来看, 作物对生态环境的适应性, 特别是光温反应、抗病、抗虫所牵涉的系统较其他性状可能更为复杂、多样, 基因的等位变异更为丰富, 位点之间组合可能更多, 从而为适应不同的环境奠定了基础。
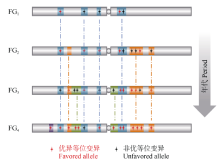
在一批受育种选择的微卫星(SSR)位点上, 无论多么优良的地方品种, 其等位变异整体组成都不能与育成品种相比[71, 72], 骨干亲本比其姊妹系品种拥有更多的优良等位变异, 新一代骨干亲本都是在上一代骨干亲本基础上进一步优化而成, 新品种比老品种拥有更多的优良基因[5, 7, 15]。因此, 早期的品种或骨干亲本无论当时多么优秀, 其田间表现和遗传背景与今天的优良品种或骨干亲本相比都有很大的差距, 这也是我们研究重要种质资源形成和演变规律的基本指导思想和出发点(图4)。
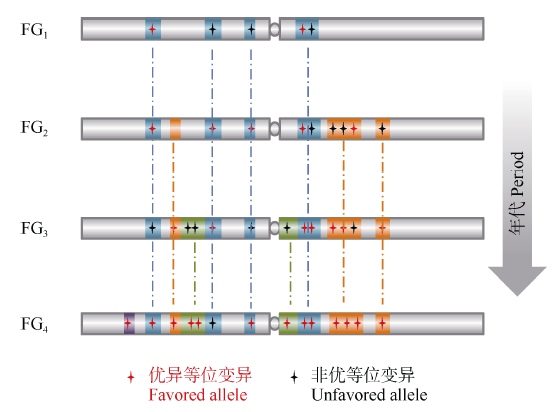 | 图4 不同时期育种骨干亲本的形成及演替规律解析, 育种的过程就是在驯化基础上对重要基因组区段、基因完善和优化的过程Fig. 4 Strategy for dissection of funder genotypes used at variant periods. Breeding gradually optimizes haplotype blocks and the crucial genes within the blocks |
远缘杂交是育种的重要组成部分, 但与常规杂交相比转移基因难度更大, 所需时间更长。一个物种越古老, 进化的时间越长, 其基因组越趋于保守, 而新物种的基因组可能更开放, 重组的机会可能更多。例如大豆、水稻与野生种基因重组已比较困难, 甚至籼、粳杂交育种也出现雌、雄半不育等困难; 而人工合成小麦与普通小麦的杂交则不存在大的障碍, 只是需要通过回交就可以转移有用的基因或片段。小麦族物种如黑麦、偃麦草、簇毛麦等物种的基因也可通过有性杂交、回交转移到小麦现代品种中。此外, 在甘蓝型油菜商业品种中A/C组之间基因的互换也是很普遍的现象[46], 白菜型油菜(Brassica rapa, AA)与甘蓝(B. oleracea, CC)人工杂交后合成的四倍体甘蓝型油菜, 经过持续多代双低(低芥酸、低硫苷)选择后形成的品种, 部分与芥酸、硫苷合成相关的基因丢失有关, 而油分合成相关基因的拷贝数则明显增加, 甚至发生染色体大片段的置换。多倍体物种越年轻, 基因组缓冲能力越好, 这一观点已得到很多科学家的认可, 但相关机制目前还不清楚, 依然存在争论。多数新合成的多倍体细胞分裂过程中染色体配对比较混乱, 须经多代自交才能趋于稳定, 本质是基因组间调整、适应、优化的过程。例如“ 剥离出的四倍体” 小麦(Extracted tetraploid wheat)生活力极弱, A、B基因组间甲基化、乙酰化及表达的基因数量和丰度存在显著差异, 与天然的四倍体小麦相比, 几乎无任何生存竞争力[73]。此外, 普通小麦基因组体积小于3个二倍体祖先种基因组之和(http://www.wheatgenome.org/News/Press-releases/Wheat-Sequencing-Consortium-Releases-Key-Resource-to- the-Scientific-Community), 因此, 多倍体物种中冗余基因和DNA片段的删除(deletion)可能比DNA片段的扩增(amplication & duplication)更普遍。所以克隆一些近缘种属的基因, 通过转基因或基因编辑技术实现有用基因的转移可能会取得更快、更好的效果。
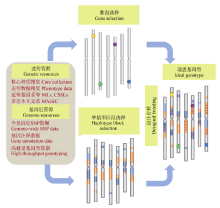
不可否认, 近100年的育种过程使现代品种集中了较多的优良等位变异、拥有了更优良的遗传背景(Haplotype blocks), 但仍有一些优良等位基因在品种改良过程中没有得到很好的利用。在提高抗病、抗逆基因的变异, 应对气候变化所带来的病、虫害的改变、病源小种(生物型)的变异以及生产环境的非稳定性(旱、涝等)的育种要求下, 如何开发和利用这批潜在的、有较大育种价值的基因或区段, 提高现代品种重要基因的多样性水平, 是当前和未来作物遗传改良必须面对的一个挑战[67, 74]。将现代品种与老品种、地方品种甚至野生种杂交, 从F2开始选择育成品种的概率很小, 但通过对杂交后代的连续选择性回交, 就可以消除连锁累赘, 培育只携带目标基因(或区段)的导入系, 在此基础上通过系间互交, 实现重要基因和优良背景的高效组装, 既提高育种群体遗传多样性, 又使相关性状定向改良, 从而培育出具有实用价值(和突破性)的品种(图5), 是现代育种的一大趋势, 应给予足够的重视[1, 7, 14, 74, 75, 76]。
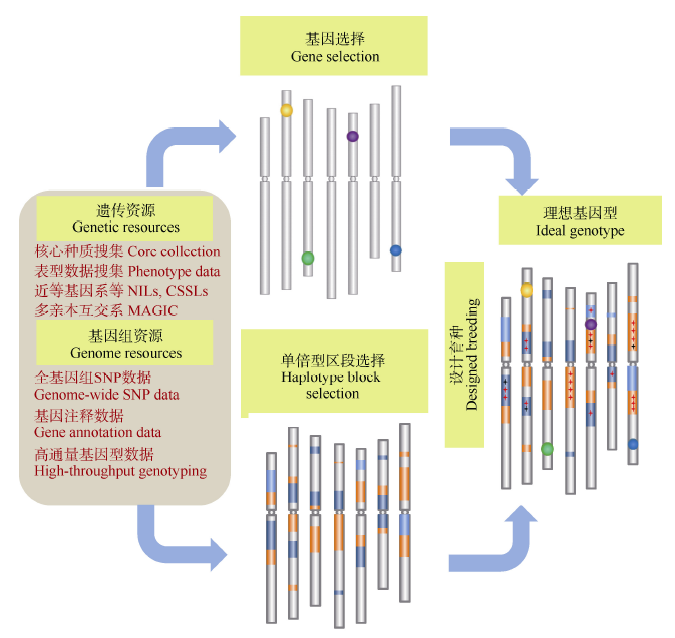 | 图5 以巢式关联群体(NAM群体)或多亲本符合杂交群体(MAGIC群体)为基础, 通过优良基因与背景的组装, 培育优良品种的基本思路Fig. 5 Strategy for creating ideatype cultivars through integrating important genes and favored haplotype blocks based on the NAM and MAGIC populations |
致谢: 感谢万建民院士和徐建龙研究员提出的宝贵意见和建议。因篇幅所限, 只引用了部分同行专家的相关文章, 敬请谅解。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|