
{kind=link}
{kind=link}
雄性不育小麦BS210育性转换特性
[孙辉**  , 张风廷
, 张风廷** , 王永波, 叶志杰, 秦志列, 白秀成, 杨吉芳, 高建刚, 赵昌平* ]
, 张风廷]
|
|


第一作者联系方式: E-mail: sunhui628@sina.com
二系法杂交小麦属我国原创性成果, 温光敏雄性不育小麦是二系法杂交小麦应用的核心。BS210是本课题组选育的小麦雄性不育材料, 2012—2014年通过3个生态区分期播种试验和人工气候箱温光调控试验, 研究了BS210的育性转换特性。在不育制种生态区, 适期播种条件下, BS210的不育度可达95%~100%, 其安全制种播种时期为安徽阜阳9月30日至10月10日、云南丽江9月30日; 在可育繁种生态区, 晚播条件下, BS210的自交结实率可达70%以上, 其安全繁种播种时期为北京海淀11月10日、北京顺义9月30日至11月10日。BS210以温度敏感为主, 光照时长对BS210育性影响较小, 温度转换敏感期为药隔至单核期。在光照10.0~12.5 h时, 其温度转换阈值为10~12°C; 在光照14 h时, 其温度转换阈值为8~10°C。在短日低温条件下, 花粉粒碘染以染败为主, 表现为不育; 在长日高温条件下, 花粉粒碘染为圆形、黑染, 表现为可育。通过明确BS210育性转换特性, 为指导BS210安全制繁种生态区的选择提供理论依据。
, ZHANG Feng-Ting
Two-line hybrid wheat is an innovation in China and its critical technique is the application of thermo-photo-sensitive male sterile line. BS210 is a male sterile wheat line developed by our group. We studied the fertility transition characteristics of BS210 with differentially sowing tests at three ecological locations and contorlled experiments in growth chambers from 2012 to 2014. In the hybrid seed-produing locations, BS210 showed sterility of 95%-100% under suitable sowing conditions with the safe sowing dates from September 30th to October 10th in Fuyang of Anhui and September 30th in Lijiang of Yunnan. In the self-reproducting locations, BS210 showed fertility of 70% under late sowing conditions with the safe sowing dates of November 10th in Haidian of Beijing and from September 30th to November 10th in Shunyi of Beijing. The fertility alteration of BS210 was induced mainly by temperature rather than photoperiod. The phase from pollen mother cell formation to mono-nucleate was sensitive to fertility alteration of BS210. The critical temperature of alteration was 10-12°C under the photoperiod of 10.0-12.5 h and 8-10°C under the photoperiod of 14 h. The pollen-staining observation indicated that the sterile pollen grains belonged to stained abortion type under short-day and low-temperature condition and the fertile pollen grains were round in shape and dark blue in color under long-day and high-temperature condition. Defining the fertility transition characteristics of BS210 will provide theoretical guidelines for selecting suitable ecological zone to ensure safe seed production.
杂交小麦被认为是今后小麦产量大幅度提高的重要途径。近50年来, 国际小麦杂种优势利用主要集中于“ 三系法” 和“ 化杀法” 两种途径, 但至今未能实现杂交小麦的大面积推广。进入21世纪, 我国创建的“ 二系杂交小麦应用技术体系” 逐步完善, 具有制种程序简便、成本低等优点, 开始进入快速示范应用阶段[1]。
小麦雄性不育研究始于1951年, 日本学者Kinara[2]将普通小麦细胞核导入尾形山羊草细胞质中产生A型雄性不育系。1962年, 美国的Wilson和Ross[3]利用提莫菲维小麦(Triticum timopheevi)与普通小麦杂交, 育成了世界上第一个具有实用价值的T型不育系, 并三系配套。1979年, Sasakuma和Ohtsuka[4]报道了具有D2型山羊草细胞质普通小麦“ 农林26” 的育性表现与光照时长变化有关; 后续研究表明, 具有D2型细胞质的农林26在≥ 15 h长日照条件下几乎雄性完全不育, 而在≤ 14.5 h短日照条件下则育性恢复[5, 6, 7]。近年来, 我国陆续发现和应用了一批不同类型温光敏雄性不育小麦材料, 如何觉民等[8]利用小麦远缘杂交, 发现了核型温光敏雄性不育材料ES-3、ES-4、ES-5等; 谭昌华等[9]通过杂交选育, 发现并培育出温光型核不育小麦C49S和C86S; 赵昌平等[10]育成了ZP35、组1、BS366等一批冬性小麦温光敏不育系; 荣德福等[11]培育了两极光敏型雄性不育小麦337S。
小麦温光敏雄性不育属于基因与环境互作雄性不育类型[12]。赵昌平等[10]提出了“ 小麦雄性育性的相对性原理” , 认为可以通过环境条件的改变实现小麦雄性器官的完全不育或完全可育, 达到生产杂交种的目的。不同类型的不育系表现出不同的育性光温转换特点。在温光敏不育小麦中, C49S温光敏感时期为孕穗期, 临界光照时长12 h, 临界温度为12~14℃[13]; ES-10温光敏感时期为雌雄蕊分化期至四分体形成期, 临界光照时长小于11.5 h, 临界温度为9~11℃[14, 15]。在温敏不育小麦中, A3314的敏感时期为孕穗期, 临界温度为18℃[16]; KTP3314A的敏感时期为四分体至单核期, 临界温度为18~20℃[17]; BNS的敏感时期为雌雄蕊分化期至四分体形成期, 临界温度为8~12℃[18, 19]。此外, BS20为低温短日不育, 敏感时期为药隔形成期至单核期, 临界温度为10~12℃[20]; 337S在穗分化阶段的短日低温和长日高温条件下均表现不育, 在适当的播期表现可育[21]。光照强度对温光敏雄性不育小麦的育性有重要影响, 但不同材料对光照强度的响应程度有差异[22]。
温光敏雄性不育小麦是二系法杂交小麦应用的核心, 其育性转换受光和温的调控。在不育条件下, 可以实现杂交种生产; 在可育条件下, 可以自我繁殖。明确不育系的育性转换特性, 可以很好地为不育系材料安全制种和繁种提供技术支撑。BS210是北京杂交小麦工程技术研究中心2000年育成的一个优良温光敏雄性不育系, 具有异交结实性好, 开颖角度大, 开颖历时长, 配合力高, 恢复源广等优点, 其配制的杂交小麦品种京麦6号, 已在生产上大面积使用。本研究通过多生态区分期播种鉴定与人工气候箱温光调控试验相结合, 进一步揭示BS210育性转换特性, 为其规模化高效安全的制种与繁殖种植区的建立提供理论依据。
小麦温光敏雄性不育系BS210源自普通小麦组合(北农2号/冀麦36)系选单株, 为红粒、冬性品种, 2011— 2012年度在3个不同生态点田间种植, 全生育期231~260 d, 株高61~78 cm, 株型半直立, 长芒, 高抗白粉病, 其主要农艺特性见表1。对照为常规小麦品种京麦9158。
| 表1 2011-2012年度BS210在3个生态点的田间表现 Table 1 Agronomic characteristics of BS210 in three test locations in the 2011-2012 winter wheat season |
2011— 2012年度和2012— 2013年度小麦生长季, 分别在北京海淀(39º 58′ N, 116º 18′ E, 海拔50 m)、北京顺义(40º 08′ N, 116º 39′ E, 海拔35 m)、安徽阜阳(32º 35′ N, 117º 14′ E, 海拔17.5 m)和云南丽江(26º 86′ N, 100º 25′ E, 海拔2416 m)进行田间试验, 其中北京海淀和顺义为BS210繁种区, 安徽阜阳和云南丽江为杂交小麦制种区。在各试点均分5期播种, 播种时期依次是9月30日、10月10日、10月20日、10月30日和11月10日。小区行长1.5 m, 种4行, 行距25 cm, 株距3 cm。每试点均设2次重复。
试验在人工气候箱(KBW, 德国Binder)内进行。温度和光照敏感期试验于2012年10月至2013年6月进行, 光温互作试验于2013年10月至2014年6月进行。试验苗于10月1日播种于花盆, 置于田间, 花盆26 cm × 23 cm, 每盆留苗8株, 底肥施磷酸二胺0.88 g、尿素0.63 g, 于四叶一心期再追施尿素0.63 g。春化完成后, 将花盆移入温室, 直到苗生长至所需生育期, 然后移入人工气候箱进行不同温度和光照处理, 非处理时期生长条件控制为光照时长12 h d-1, 抽穗前温度12℃, 抽穗后温度16℃。每处理4盆, 每盆8株, 试验重复2次。
温度敏感期试验设4个处理时期, 分别为小花分化至雌雄蕊分化期、雌雄蕊分化至药隔期、药隔至四分体期、四分体至单核期。低温(10℃)处理, 光照时长12 h d-1, 处理时间8 d。
光照敏感期试验设3个处理时期, 分别是二棱至小花分化期、小花分化至药隔期、药隔至单核期; 设3个光照时长处理, 分别为10、12和14 h d-1, 温度12℃, 处理时间16 d。
光温互作为药隔至单核期的双因子交互试验, 处理平均温度为8℃、10℃、12℃、14℃、16℃ (图1), 处理光照时长为10、12和14 h d-1。小麦抽穗期挂牌定株, 抽穗后3~4 d, 取穗中部第1位或第2位小花的花药, 用碘-碘化钾染色, 观察花粉粒育性。
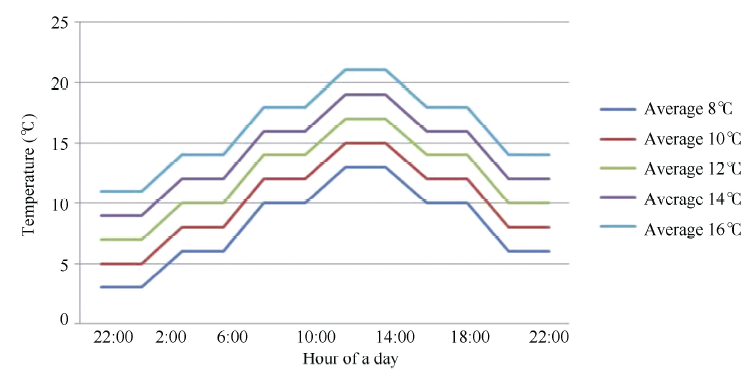 | 图1 人工气候箱5个温度处理的日温设置Fig. 1 Daily temperature control of the growth chamber for the five treatments |
于开花前对每株主茎穗和1~2个分蘖穗套袋, 成熟后调查小穗数和穗粒数。结实率(%) = 穗粒数 / (小穗数 × 2) × 100。采用SPSS20.0软件统计分析, 用Duncan’ s法进行多重比较。
在不同生态点的分期播种试验中, BS210在繁种区的结实率明显高于制种区。在北京海淀, BS210晚播结实率高于早播, 11月10日播种的结实率最高, 2011— 2012与2012— 2013年度分别为82.82%和73.09%, 显著高于10月30日前的播种处理, 表明11月10日播种可安全繁种。在北京顺义, 随着播期推迟结实率有所增加, 但不同播期之间结实率差异较小, 也是以11月10日播种的结实率最高, 两年度结实率分别为76.42%和78.60%, 表明9月30日至11月10日播种均可安全繁种(表2)。
| 表2 BS210和京麦9158在3个生态点的结实率 Table 2 Seed-setting rate of BS210 in three test locations (%) |
在安徽阜阳和云南丽江, 不同播种时期之间晚播结实率都显著高于早播。在阜阳, 两年度试验中10月30日、11月10日播种与9月30日、10月10日播种相比结实率差异显著, 9月30日和10月10日播种的结实率低于2%, 达到高度不育, 为安全制种播种时期。在丽江, 9月30日播种两年度的结实率分别为2.05%和1.59%, 为安全制种播种时期; 10月20日、10月30日和11月10日播种与前两个播种时期相比结实率显著提高(表2)。
对照品种京麦9158虽然不同地区分期播种晚播结实率高于早播结实率, 但差异不显著, 不同年份、不同地域、不同播种时期的结实率均达到80%以上(表2)。
以上分析表明, 不同生态区、播种时期对BS210和京麦9158结实率影响是不同的。不同地域、播种时期对BS210的育性影响非常明显, 结实率差异显著; 对京麦9158的育性影响较小, 育性表现正常。
2.2.1 BS210育性转换的温度敏感期 在不同的穗分化阶段进行低温(10℃)处理, BS210的结实率差异很大, 以药隔至四分体期的结实率最低, 其次是四分体至单核期, 且显著低于其他生育期(表3), 说明该穗分化阶段育性对低温最敏感, 低温与该生育阶段相遇则制种安全性较高。对照京麦9158各处理的结实率均达70%左右, 育性表现稳定和正常, 对低温不敏感。
| 表3 BS210在不同穗分化阶段低温(10 ° C)处理的结实率 Table 3 Seed-setting rate of BS210 treated with low temperature (10 ° C) at different phases of spike initiation (%) |
2.2.2 BS210育性转换的光照敏感期 3个生育阶段的3个光照长度处理表明, BS210在相同处理期不同光照时长下的结实率差异不显著, 说明BS210对光照时长反应不敏感。药隔至单核期各光照时长处理, BS210的结实率介于10%~20%之间, 而另两个时期处理的结实率都在40%以上(表4), 原因是药隔至单核期是BS210的温度敏感时期, 本试验的背景温度是12℃, 这一温度导致BS210在药隔至单核期结实率较低, 而非光照时长直接引起的结实率低。对照京麦9158在所有处理中的结实率均在70%以上, 表现正常, 对不同光照时长和处理期反应均不敏感(表4)。
| 表4 BS210在3个穗分化阶段不同光照长度下的结实率 Table 4 Seed-setting rate of BS210 under different photoperiods treated at three phases of spike initiation (%) |
2.2.3 光温互作条件下BS210的育性转换 在明确了BS210温度、光照敏感期的基础上, 为了精确分析BS210温光互作关系, 利用人工气候箱进行5× 3温光互作试验。结果表明, 温度对BS210结实率的作用极显著, 光照时长作用不显著, 温光互作效应不显著(表5)。因此认为, BS210育性转换过程中, 温度对育性起主要作用。
| 表5 温光互作试验的方差分析 Table 5 Analysis of variance on the effects of temperature-photoperiod interaction |
在各个光照时长处理下, BS210的结实率都是随着处理温度的提高而增加, 均温8℃处理结实率最低, 均温12℃以上结实率显著提高, 均温16℃处理结实率最高。在光照10 h d-1和12 h d-1条件下, BS210在均温8℃和10℃时结实率都低于5%, 且两处理差异不显著, 而均温12℃以上处理结实率超过15%, 显著高于均温8℃和10℃处理; 在光照14 h d-1条件下, BS210在均温8℃时结实率低于5%, 而均温10℃处理的结实率显著升高至8.61% (表6)。因此, BS210育性转换, 光照10 h d-1和12 h d-1时温度阈值为10~12℃, 光照14 h d-1时温度阈值为8~10℃。
| 表6 温光互作处理下BS210的结实率 Table 6 Seed-setting rate of BS210 under temperature-photoperiod interaction (%) |
阜阳点的田间试验与人工气候箱试验结果一致。在阜阳, 随着播期推迟, BS210的结实率逐渐升高, 所对应的温度敏感时期平均气温也不断提高(表7)。9月30日和10月10日播期处理, 两年的结实率都低于5%, 敏感时期平均温度为10.3~11.6℃; 播期推迟至10月20日或更晚时, BS210的结实率超过10%, 并随着播期推迟而逐步增加, 与此同时, 敏感时期的平均温度也逐步提高。阜阳地区在敏感期内每天平均光照时长约为12.5 h, 田间观测结果进一步证实, 在此光照条件下BS210的温度转换阈值为10~12℃。
| 表7 BS210在安徽阜阳温度敏感时期平均温度和自交结实率 Table 7 Seed-setting rate and sensitive period to temperature of BS210 grown in Fuyang, Anhui province |
BS210在短日低温(10 h d-1, 10℃)条件下, 花药不开裂, 碘-碘化钾染色表明, 花粉粒形状多数正常, 但不能被染色或染色浅, 花粉粒败育; 在长日高温(14 h d-1、16℃)条件下, 花药开裂, 花粉粒为圆形、被染成黑色, 花粉粒可育(图2)。
 | 图2 不同温光条件下BS210花药及花粉粒碘染情况 A: 10 h d-1和10℃条件下, BS210花药不开裂; B: 14 h d-1和16℃条件下, BS210花药开裂; C: 10 h d-1和10℃条件下, 花粉粒不能被碘-碘化钾染色或染色浅; D: 14 h d-1和16℃条件下, 碘-碘化钾染色后花粉粒呈黑色。Fig. 2 Observation of anthers and pollen grains of BS210 under different temperature-photoperiod conditions A: no anther dehiscence of BS210 under 10 h d-1 and 10 ° C condition; B: anther dehiscence of BS210 under 14 h d-1 and 16 ° C condition; C: anthers were not stained or lightly stained by I2-KI under 10 h d-1 and 10° C condition; D: anthers were stained black with I2-KI under 14 h d-1 and 16° C condition. |
利用小麦雄性不育系生产杂交种, 是目前小麦杂种优势利用的最有效途径。随着中国二系杂交小麦技术体系的构建完成, 选育各种类型优良温光敏雄性不育系已成为二系杂交小麦的重点内容。大量测试结果表明, 以温度敏感主导的雄性不育是小麦温光敏雄性不育资源群体中的主体, 优良温敏型不育系应用的关键是安全制种生态区的选择, 跨区域的异地制种有利于提高二系杂交小麦制种的纯度和安全性。本研究表明, BS210以温度敏感为主, 在北纬33° 以南的安徽阜阳、云南丽江, 适期播种时可高度不育, 适合杂交制种。本课题组2002— 2015年在这两地进行大面积制种, 制种纯度为96%~99%; 而在北纬40° 以北的北京地区晚播种植, BS210育性可恢复到70%以上。2013— 2015年, 以BS210为母本配置的杂交小麦组合中, 筛选出比临近对照品种中麦175增产10%以上的组合4个, 其中一个组合增产达20.1% (表8)。表明不育系BS210不仅育性稳定, 已初步建立了繁种和制种体系, 而且由BS210配制的杂交组合具有明显的增产效果, 具有良好的应用前景。本研究通过人工气候箱试验以及多年多点的田间观测, 充分验证了BS210育性转换的温度和光照条件, 明确了该不育系在繁种和制种时敏感期和温度转换阈值, 并提出了繁种地和制种地的适宜播种时期, 为生产上进一步安全、有效地利用BS210提供了理论支持。
| 表8 2013-2015年BS210配制的杂交小麦组合的产量表现 Table 8 Yield potential of hybrid wheat derived from BS210 in 2013-2015 |
不同类型的温光敏不育系具有不同的温光反应特点, 将多生态区多年分期播种试验与精密人工气候箱温光调控试验相结合, 是揭示不育系不育特性的有效方法, 为二系杂交小麦科学安全利用该不育系提供有价值的理论指导。ES-8、ES-10、BNS的敏感时期在雌雄蕊至四分体时期[14, 15, 18, 19, 23]; C49S、A3314的敏感时期在孕穗期[13, 16]。本研究表明, BS210育性转换的温度敏感时期为药隔至单核期, 不同于上述不育系, 其温度敏感时期处于上述两类不育系之间。BS210表现温度敏感为主, 不论是人工气候箱温光调控试验还是田间试验, 温度转换阈值范围是一致的, 在光照10.0~12.5 h d-1时, 其温度转换阈值为10~12℃; 在光照14 h d-1时, 温度转换阈值为8~10℃, 这与C49S的临界温度12~14℃、A3314的临界温度18℃存在差异[13, 16], 但与BNS (8~12℃)、ES-10 (9~11℃)和BS20的临界温度(10~12℃)较为接近[14, 18, 19, 20]。另外, 碘-碘化钾染色结果表明, BS210花粉粒以染败为主, 不同于ES-14、K78S、BNS以典败为主的花粉粒败育类型, 而与ES-10、A3017、BS20的败育类型相同[15, 20, 24, 25, 26]。
BS210以温度敏感为主, 败育类型以染败为主, 温度转换敏感期为药隔至单核期, 温度转换阈值受光周期影响, 在光照10.0~12.5 h d-1时, 温度转换阈值为10~12℃; 在光照14 h d-1时, 温度转换阈值为8~12℃。明确了BS210在安徽阜阳、云南丽江制种和北京繁种的安全播种时期, 为该不育系的大面积应用提供了重要的理论指导。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|