
{kind=link}
{kind=link}
{kind=link}
{kind=link}
甘蓝型油菜盐胁迫下幼苗鲜重和干重QTL定位及候选基因分析
[侯林涛**  , 王腾岳
, 王腾岳** , 荐红举, 王嘉, 李加纳, 刘列钊* ]
, 王腾岳]
|
|


第一作者联系方式: E-mail: 594632283@qq.com
盐胁迫是非生物胁迫中影响作物产量的一个主要因素, 利用分子标记方法选育油菜耐盐品种对提高油菜产量具有重要意义。选用来自GH06与P174杂交后通过单粒传法连续自交获得的高世代重组自交系群体, 以含16 g L-1 NaCl的Hoagland溶液培养幼苗进行盐胁迫处理25 d后, 分别测定叶和根的鲜重及干重, 根据已构建的高密度SNP遗传连锁图谱进行QTL定位, 在QTL物理区间筛选耐盐相关基因并以极端表型材料进行qRT-PCR分析。采用复合区间作图法(CIM), 在对照和盐胁迫处理中共检测到19个QTL, 其中与盐胁迫相关的有6个, 可解释的表型变异7.16%~16.15%, 分布在A02、A04和C03染色体上, 将QTL置信区间序列和拟南芥中与盐胁迫相关的基因比对分析, 共找到8个候选基因。对其中4个候选基因在极端表型材料中的表达分析表明, BnaA02g14680D与 BnaA02g14490D基因在盐胁迫处理后的48 h或72 h表达量均高于对照组, 即基因的表达由盐胁迫引起, 而 BnaC03g64030D在敏感型材料中的相对表达量高于在耐盐型材料中, BnaC03g62830D在敏感型材料中没有明显变化, 但在耐盐型材料中呈现先升高后降低的表达特征, 其表达可能会增强植株对盐胁迫的耐受力。本研究为油菜耐盐基因功能挖掘和油菜耐盐品种选育奠定基础。
Salt stress is one of the main abiotic stresses affecting crop yield and it would be very important by using the salt tolerance related markers in rapeseed breeding to improve the oilseed production. In this research, the Brassica napus L. high generation recombinant inbred lines (RIL) population derived from the cross of GH06 and P174 via single seed descent propagation was used for QTL mapping and candidate gene analysis. The fresh and dry weight of leaf and root were measured at 25 days after the seedlings were grown in Hoagland solution with 16 g L-1NaCl. Composite interval mapping (CIM) was used to identify the related QTLs according to the high density SNP genetic map, and the candidate gene expression in the extreme lines tested by qRT-PCR. A total of 19 QTLs were identified in the control and salt stress treatment, and six QTLs were mapped on chromosomes A02, A04, and C03 under salt stress, with contribution rate ranged from 7.16% to 16.15%. Eight genes were identified according to the BLAST of genes in the QTL confidence intervals and the salt stress related genes in Arabidopsis. The expression of four candidate genes in the extreme lines showed that BnaA02g14680D and BnaA02g14490D under salt stress treatment for 48 or 72 hours had higher expression than the control, which indicates that the expressions are induced by salt stress. The relative expressions of gene BnaC03g64030Din sensitive extreme lines were higher than those in tolerant extreme lines. There were no changed in expression for gene BnaC03g62830D in sensitive extreme lines but increased expression at 48 hours and reduced expression at 72 hours after salt treatment in tolerant extreme lines, showing the enhance of plant salt tolerance possibly. Our research laid a foundation for the function research of salt tolerant gene in rapeseed and the breeding of salt tolerant rapeseed.
土壤盐渍化是危害农业生产的主要因素之一, 中国盐渍土面积大、分布广、类型多, 总面积达9913万公顷, 约占国土面积的1.03% [1]。土壤中过量的盐分会引起土壤物理和化学性质的改变, 导致农作物生长环境的恶化[2]。在我国, 油菜是继水稻、玉米、小麦、大豆之后的第五大作物, 也是第一大油料作物, 常年种植面积约670万公顷, 总产超过1100万吨, 年产菜籽油约450万吨, 占自产植物油总量的40%以上, 占草本植物油总量的57%以上[3]。甘蓝型油菜在三大栽培类型油菜中种植面积最大, 被认为是比较耐盐的作物之一[4]。因此我们期望通过甘蓝型油菜耐盐性定位研究为其耐盐性和品种选育提供研究基础, 进而实现盐碱地的农业高效利用, 这对我国耕地农业生产能力的提升, 耕地数量的增加, 国家粮食安全的保障具有重要意义[5]。
随着对油菜研究的不断深入及SNP芯片和重测序的普及, 利用SNP分子标记技术建立高密度的遗传图谱, 对油菜重要性状的QTL分析已经成为新常态, 并且已经定位了与油菜含油量、千粒重、株高、产量等相关的QTL [6, 7, 8, 9, 10], 对于油菜在盐胁迫条件下的研究大多集中在生理反应的测定[11, 12, 13], 而疏于对油菜耐盐性状基因的定位。Li等[14]利用甘蓝型油菜DH系群体在正常和干旱胁迫条件下的5个指标(株高、根长、叶干重、根干重和总重)数据的QTL分析, 分别检测到28个和31个QTL, 其中在正常环境下, 以油菜幼苗干重为指标检测到18个QTL, 单个位点可解释的表型变异为6.43%~18.97%; 旱胁迫环境下, 检测到与幼苗干重相关的20个QTL, 单个位点可解释的表型变异为5.87%~15.97%。荐红举等[15]利用本研究的RIL群体对盐胁迫下种子发芽率的QTL分析, 检测到11个相应的QTL, 位于A03和A09染色体上, 可解释的表型变异为4.9%~10.9%。Moursi[16]利用DH群体在正常和盐胁迫环境下的干重和鲜重数据进行QTL分析, 正常环境下找到4个QTL, 分别位于A06、A05、C02和C03染色体上, 贡献率为36%; 盐胁迫环境下找到3个QTL, 分别位于C03和C06染色体上, 共同可解释的遗传变异25%。
本试验利用已经构建的高密度SNP遗传连锁图谱对盐胁迫环境下RIL群体叶和根的鲜重和干重进行QTL定位, 进而筛选出与盐胁迫相关的候选基因并分析其在极端表型材料中的表达量, 为油菜耐盐基因的进一步研究提供理论基础。
以母本GH06与父本P174杂交, 采用单粒传法连续自交10代, 构建高世代重组自交系群体(recombinant inbred line, RIL), 之后随机选取其中的172个重组自交系进行SNP标记分析, 构建高密度SNP遗传连锁图谱[17]。
挑选健康饱满、大小均一的油菜种子, 置铺2层定性滤纸的9 cm培养皿内, 每皿20粒, 加入适量的1/4 Hoagland营养液培养7~8 d。待种子发芽长成幼苗时, 从每株系各取生长健康一致的1植株, 3个重复, 分别置装有质量浓度为0 g L-1和16 g L-1NaCl溶液[15](以Hoagland营养液为母液配制)的塑料盆中, 于温室培养25 d, 设定温度25℃, 光照16 h, 黑暗8 h。
用滤纸吸干每株系3个植株水分后分别用电子天平测量幼苗的叶鲜重、根鲜重, 取平均值。将幼苗放入105℃烘箱杀青30 min, 70℃烘箱烘干48 h至恒重, 然后用电子天平称量幼苗的叶干重、根干重, 取平均值。
所用SNP分子标记构建的遗传连锁图谱[17], 包含2795个SNP多态性标记位点, 总长1832.9 cM, 相邻标记间平均距离为0.66 cM。采用QTL分析软件Windows QTL Cartographer 2.5[18]及复合区间作图(composite interval mapping, CIM)法对RIL群体幼苗的叶鲜重、根鲜重与叶干重、根干重进行QTL定位及效应检测[19]。CIM分析时, 选取1 cM的步长(walking speed), 按照假定检测10和Zmapqtl模型3, 选取参数1000次回归, 显著水平为0.01。LOD≥ 3时, 即认为该区间可能存在一个QTL。运行软件同时给出性状QTL的加性效应和解释的表型变异。按照McCouch等[20]的方法命名检测到的QTL, 以“ q” 加相对应的性状再加染色体编号表示, 字体为斜体。
将检测到的QTL的置信区间在甘蓝型油菜基因组[21]上查询到对应的序列, 然后与拟南芥基因组序列进行BlastN, E值设定为E-20, 最后筛选出每个QTL置信区间内匹配E值小于阈值的盐胁迫相关基因, 共筛选出8个与盐胁迫相关的基因。
根据RIL群体叶、根鲜重在盐胁迫环境与正常环境下的比值, 筛选出对盐敏感型材料(F215的比值分别为0.78和0.42, F328分别为0.76和0.37)和耐盐型(F285的比值为分别1.02和1.50, F293分别为1.00和1.38)的极端表型材料, 在盐胁迫处理24、48、72和96 h后取叶片, 将样品迅速于-80℃保存。按照EASYspin植物RNA快速提取试剂盒(RN09)说明提取样品的总RNA。采用Bio-Rad iScript cDNA Synthesis Kit试剂盒反转录, 反转录体系为5× iScript reaction mix 4 μ L, iScript reverse transcriptase 1 μ L, RNA模板(1 μ g) 1~3 μ L, ddH2O补充体积至20 μ L。反应条件为25℃ 5 min, 42℃ 30 min, 85℃ 5 min, 4℃保存。
在甘蓝型油菜基因组[21]中查找到候选基因的cDNA序列, 利用软件Primer Premier 5.0设计特异引物, 由上海生工生物工程股份有限公司合成(表1)。按照Bio-Rad公司的SsoAdvanced Universal SYBR Green Supermix 试剂盒使用说明在Bio-Rad CFX96上运行实时荧光定量PCR。反应体系为SsoAdvanced universal SYBR Green supermix (2× ) 10 μ L, 正向引物0.4 μ L, 反向引物0.4 μ L, cDNA 2 μ L, ddH2O补充体积至20 μ L。反应初始变性温度和时间为95℃/30 s, 然后95℃/10 s, 退火时间为30 s, 温度因引物而异, 40个循环。熔解曲线为65~95℃之间以0.5℃递增, 5 s。所有反应3个重复。用2-Δ Δ CT方法以内参(油菜Actin基因-AF111812)为标准计算每个基因的相对表达量。
| 表1 荧光定量PCR引物 Table 1 Primers used for RT-PCR |
利用SPSS 17.0软件完成相关性分析。利用MapChart 2.2作图软件分析染色体和QTL之间的遗传距离, 在Microsoft Excel 2003和WPS 2016中处理其他数据。
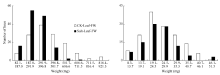
RIL群体的鲜重、干重, 各数值均呈连续分布和双向超亲分离(表2、图1和图2), 表明这些性状均为多基因控制的数量性状, 其偏斜度和峰值均小1, 适合于QTL分析。除了盐胁迫处理下根干重均值高于对照组外, 其余叶、根的鲜重、干重均值均低于对照组, 这表明盐胁迫环境对幼苗的生长和发育产生了影响, 而根干重高于对照组可能由于幼苗在盐胁迫环境下促进根系的生长。盐胁迫下的叶鲜重均值要远远低于对照组, 而叶干重却相差不大, 可能是由于盐胁迫条件下离子浓度过高而影响植株的水分吸收。
| 表2 油菜幼苗在盐胁迫处理和对照处理下干重、鲜重的分布特征 Table 2 Phenotype variation of dry weight and fresh weight under salt stress and control conditions |
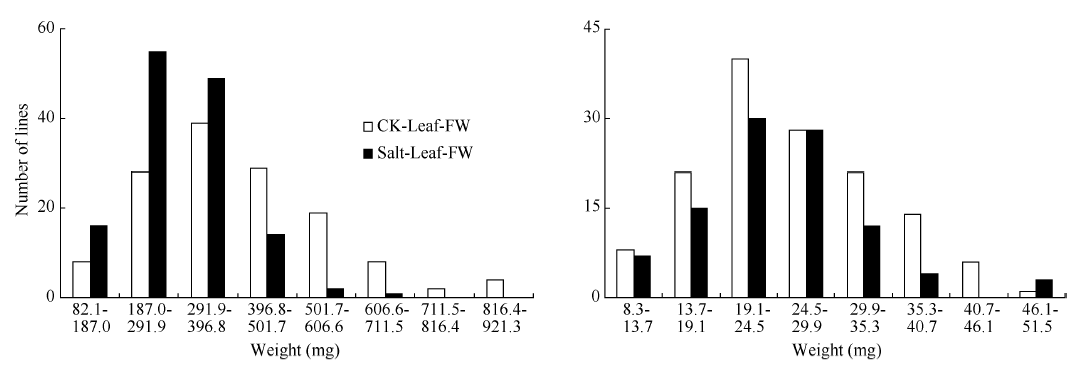 | 图1 油菜幼苗叶片和根鲜重在盐胁迫和对照下的频率分布图Fig. 1 Frequency distribution of fresh weight of B. napusunder control and salt stress conditions |
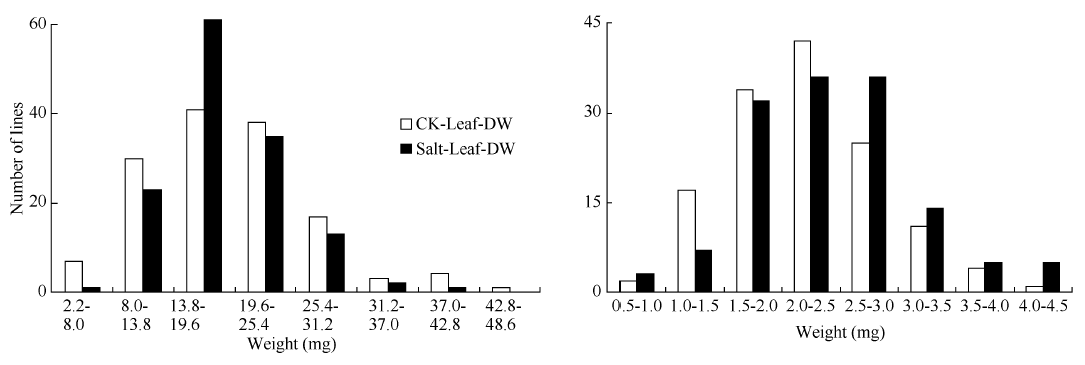 | 图2 油菜幼苗干重在盐胁迫处理和对照处理下的频率分布图Fig. 2 Frequency distribution of dry weight of B. napus under salt stress and control conditions |
从表3可以看出2种处理下的叶干重显著正相关, 相关系数为0.205。盐胁迫环境下叶干重和根干重极显著正相关, 相关系数为0.771, 盐胁迫环境下叶鲜重和根鲜重极显著相关, 相关系数为0.380。相关性分析表明, 相同的处理下, 相同部位干重和鲜重均极显著相关。在对照和处理间, 只有叶片的干重和鲜重表现显著和极显著相关, 相关系数分别为0.205和0.246, 而根部在对照和处理间相关性不显著。
| 表3 油菜幼苗盐胁迫处理和对照干重、鲜重的相关系数 Table 3 Correlation coefficients between indication of dry weight and fresh weight of B. napus in salt stress treatment and control |
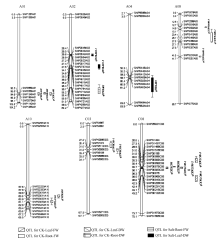
总共检测到13个QTL (表4和图3)。油菜叶和根干重相关的QTL有4个, 分布在染色体A01、A10和C08上, 单个QTL可解释的表型变异为8.43%~ 12.19%。其中C08染色体上的qCKDLC08贡献率为12.19%, LOD值为6.28。油菜幼苗叶和根鲜重的QTL有9个, 单个QTL可解释的表型变异为7.08%~ 15.08%, 分布在A01、A08、A10和C08染色体上。其中分布在C08染色体上的QTL有4个, 贡献率为7.56%~15.08%, 置信区间有部分重叠。检测到A08染色体上3个QTL, 贡献率为8.00%~8.84%, 置信区间相互靠近。在A01染色体上的QTL qCKWLA01和qCKDLA01置信区间完全重合, 推测在此区间内可能有控制这一性状的基因。
| 表4 利用复合区间作图法检测油菜幼苗干重、鲜重在正常环境下的QTL Table 4 Putative QTL detected by composite interval mapping for dry weight and fresh weight of B. napus in normal environment |
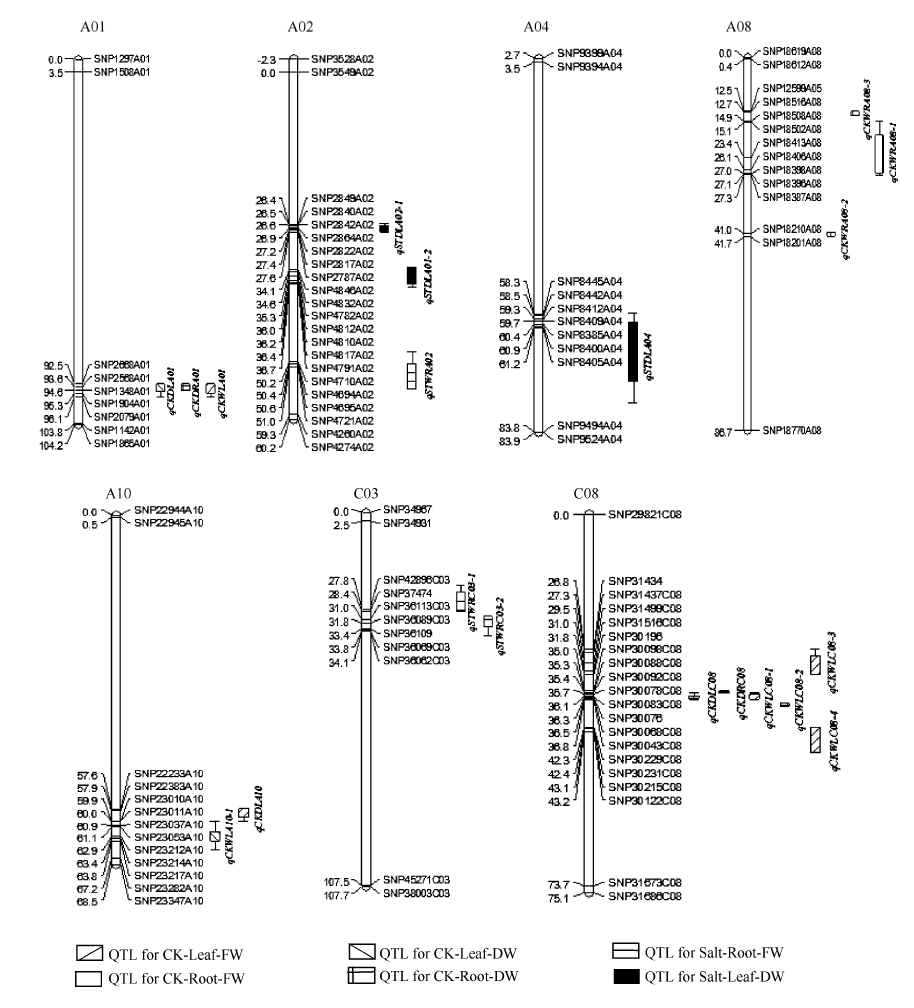 | 图3 甘蓝型油菜幼苗盐胁迫处理下干重、鲜重QTL在连锁群上的分布情况 QTL的染色体上仅展示了QTL区段的标记及染色体两端各2个标记。Fig. 3 Putative QTL locations of dry weight and fresh weight in salt stress treatment on the SNP genetic map Showing the markers in the QTL confidence intervals, along with the terminal two markers at each end of the QTL containing chromosomes. |
检测到与盐胁迫相关的QTL 6个(表5和图4), 阈值为3.21~4.59, 单个QTL可解释的表型变异为7.16%~16.15%, 分布在染色体A02、A04和C03上。分布在A02染色体上的QTL有3个, 单个QTL可解释的表型变异为7.16%~11.75%。A04染色体上1个QTL, LOD值为4.66, 贡献率为11.60%, 置信区间为58.00~77.40。C03染色体上2个QTL, 为qSTWRC03和qSTWRC03, LOD值分别为4.47和4.59, 共同可解释的表型变异是31.75%, 置信区间有部分重叠。
| 表5 利用复合区间作图法检测到油菜幼苗干重、鲜重在盐胁迫环境下的QTL Table 5 Putative QTL detected by composite interval mapping for dry weight and fresh weight of B. napus in salt stress environment |
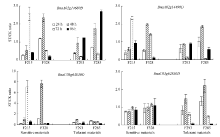
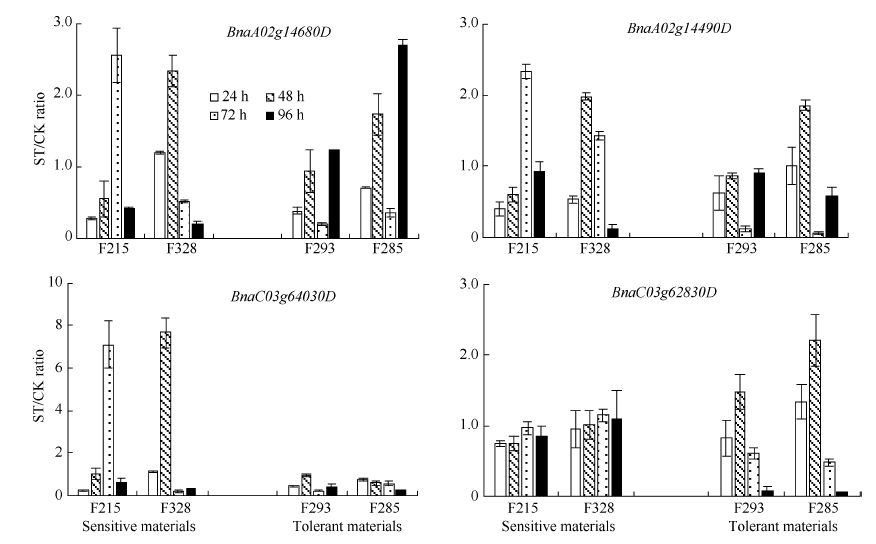 | 图4 甘蓝型油菜4个基因在盐胁迫下表达量的比值Fig. 4 Expression ratio of four genes under salt stress (ST) and normal environment (CK) in B. napus |
在盐胁迫环境下共检测到6个QTL, 但由于qSTWRA02和qSTWRC03-2在甘蓝型油菜基因组中没有找到相应的序列, 故只能将4个QTL置信区间序列与拟南芥中盐胁迫相关基因比对, 共检测到8个候选基因(表6和表7)。其中在qSTDLA02-1区间内1个, 匹配E值为E-137。qSTDLA02-2区间内3个, 匹配E值介于0~2E-52之间。qSTDLA04区间内只1个, 匹配E值为5E-25。qSTWRC03-1区间内3个, 匹配E值为4E-35~E-167。
| 表6 甘蓝型油菜基因组中QTL置信区间候选基因与拟南芥盐胁迫相关基因的比对 Table 6 Alignment of candidate genes in QTL confidence interval in B. napus with the salt related genes in A. thaliana |
| 表7 筛选的在拟南芥中盐胁迫相关候选基因功能 Table 7 Functions of the screened candidate genes related to salt stress in A. thaliana |
由图4可知, 在敏感型材料中, BnaA02g14680D基因盐处理后的48 h或72 h表达量较高, 之后96 h表达量降低; 而在耐盐型材料中, BnaA02g14680D基因在盐处理后的表达量先升高后降低之后在96 h又升高。BnaA02g14490D基因与BnaA02g14680D基因的表达水平特征相似。而BnaC03g64030D基因在敏感型材料中盐处理后的表达水平都有急剧升高的时期, 之后下降; 而其在耐盐型材料中的表达量则没有明显变化。BnaC03g62830D基因则在敏感型材料中没有明显变化, 在耐盐型材料中呈现先升高后降低的表达特征。
作物生长会受到多种因素的影响, 包括生物胁迫和非生物胁迫, 非生物胁迫中盐胁迫成为限制作物产量的一个重要因素[30]。油菜的营养价值和经济价值极高, 已经成为植物油的主要来源, 然而其生长、产量和出油量会因为土壤盐渍化而有所降低, 特别是油菜种子发芽和幼苗早期的生长发育更容易受到盐害的影响[31]。从油菜幼苗在正常环境下和盐胁迫环境下鲜重和干重的分布特征可以看出, 盐胁迫下叶鲜重、干重的分布范围较窄, 均值也低于对照组, 尤其是叶鲜重, 盐胁迫环境下的叶鲜重均值几乎为对照组的一半, 根干重和鲜重则变化不大。推测可能是由于外界环境盐浓度高而影响了幼苗对水分的吸收, 从而导致植株叶鲜重有所下降, 这一结果与Tunç tü rk等[32]的研究结果一致。
植物对盐胁迫的响应是一个复杂的生理过程[33, 34], 它会引起很多相关性状的改变, 如种子发芽率、植株鲜重干重等。我们采用SNP高密度遗传连锁图谱对正常环境和盐胁迫环境下的幼苗重量QTL分析, 发现两种环境下检测到的QTL有很大差异, 且分布的染色体也不同, 可能是由于油菜幼苗在不同处理下差异基因适应环境而产生不同的响应。盐胁迫环境下共检测到6个QTL, 其中分布在染色体A02上的有3个, 置信区间十分靠近且有部分重叠, A04染色体上有1个, 加性效应均来自于父本P174; C03染色体上有2个, qSTWRC03-1与qSTWRC03-2共同可解释的表型变异为31.75%, 是盐胁迫环境下找到的贡献率最高的两个QTL, 而在正常环境下并没有检测到, 因此推测它们可能是与盐胁迫环境下幼苗根鲜重相关的主效QTL, 加性效应来自于母本GH06。Moursi[16]利用DH群体对正常和盐胁迫环境下干重和鲜重得到的数据进行QTL分析, 盐胁迫环境下找到3个QTL, 共同可解释的遗传变异为25%, 分别位于C03和C06染色体上, 但与qSTWRC03-1和qSTWRC03-2位置相距较远。荐红举等[15]利用本研究的RIL群体对盐胁迫下种子发芽率进行QTL分析, 检测到11个相应的QTL, 可解释的表型变异为4.9%~10.9%, 位于A03和A09染色体上, 与我们检测到的QTL所在染色体不一致, 推测可能是由于油菜在盐胁迫环境下种子发芽和植株生长响应机制不同。
本研究将盐胁迫环境下检测到的4个QTL置信区间与拟南芥盐胁迫相关基因比对, 在每个区间内都找到了相应的候选基因, 共8个, 并对其中4个进行实时荧光定量PCR分析。研究结果表明, BnaA02g14680D基因在拟南芥中对应基因ATRPK1/RPK1的表达是由脱落酸、脱水、高盐和低温引起的, 本实验中该基因在盐胁迫环境下的表达量明显高于对照组, 与Hong等[23]的研究结果一致。Osakabe等[35]在实验中也发现在拟南芥中过量表达RPK1基因会增强对盐胁迫的耐受力。但Shi等[36]认为ATRPK1/RPK1基因的过量表达会使拟南芥对盐胁迫的耐受力降低, 当ATRPK1基因的表达量被限制时, 拟南芥对盐胁迫的耐受力明显升高。BnaA02g14490D基因在拟南芥中对应基因ATHVA22C的表达也是由脱落酸和盐胁迫引起的, 在敏感型和耐盐型材料中的表达特征与BnaA02g14680D基因相似, 在耐盐型材料中两个基因在盐处理后96 h表达量明显升高(图4), 推测可能是由于基因在耐盐型材料中的表达量较高而使其对盐胁迫环境的耐受力增强。BnaC03g64030D基因在拟南芥中对应ATCPL1/SHI4基因功能为编码的蛋白质在体外培养具有先天性磷酸酶活性, 突变体对脱落酸, 低温和氯化钠十分敏感[26], 其在敏感型材料中的表达量在48 h或72 h高于耐盐型材料(图4), 可能是由于其对盐胁迫环境更为敏感而高量表达以响应盐胁迫。BnaC03g62830D基因在拟南芥中对应ATCESA8/LEW2基因功能为编码一个纤维素合酶家族的成员, 其参与次生细胞壁的生物合成[28], 与对照组相比, 该基因在耐盐型材料中的盐处理后的表达量在48 h明显升高(图4), 之后降低, 推测是由于受到盐胁迫环境影响而高量表达以增强对盐胁迫环境的耐受力, 而敏感性材料则没有, 所以对逆境的耐受力较差。Long等[37]通过RNA测序和PCR荧光定量方法在油菜中找到31个与盐胁迫相关的基因, 但与我们发现的基因位点不一致, 推测可能是由于取材时间和部位不同, 由基因表达的时间和组织特异性引起的。
盐胁迫环境下油菜幼苗的鲜重和干重均低于对照组, 表明盐胁迫对油菜生长有一定的影响。在染色体A02、A04和C03定位了6个盐胁迫环境下油菜幼苗干重鲜重的QTL, 贡献率超过10%的QTL有4个, 表明油菜耐盐性是由多个微效基因调控的数量性状。共找到8个候选基因, 对其中4个基因在极端表型材料中的qRT-PCR分析表明, BnaA02g14680D和BnaA02g14490D基因的表达由盐胁迫引起, BnaC03g62830D基因相对高量表达可能会增强植株对盐胁迫环境的耐受力。今后的研究应对筛选出的候选基因进行功能验证等深入分析, 为耐盐基因定位和盐胁迫相关机制的研究提供理论依据。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|