


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
根-冠互作对棉花叶片衰老的影响
[张巧玉 , 王逸茹, 安静, 王保民, 田晓莉
, 王逸茹, 安静, 王保民, 田晓莉* ]
, 王逸茹, 安静, 王保民, 田晓莉]
|
|


第一作者联系方式: E-mail: qiaoyu306@126.com, Tel: 010-62734550
嫁接研究和实践表明, 植物不同生理过程普遍存在根-冠互作的多样性。作者曾以衰老较快的棉花品种中棉所41和衰老较慢的品种鲁棉研22为材料进行嫁接研究, 发现地上部对叶片衰老起主要作用。本文以中棉所41和另一个衰老较慢的品种中棉所49为材料进行普通嫁接(I型, 单接穗单砧木)和Y型嫁接(双接穗单砧木), 用低钾胁迫(0.03 mmol L-1)诱导叶片衰老, 结果表明I型嫁接中以中棉所41为砧木的处理, 其倒四叶SPAD值显著低于以中棉所49为砧木的处理, 提示根系对叶片衰老的作用较大。相应地, 以中41为砧木的处理, 其根系和叶片中的ZR+Z和iPA+iP浓度及砧木和接穗木质部汁液中的ZR+Z和iPA+iP流量在绝大多数情况下显著低于以中49为砧木的处理, ABA的结果则相反。Y型嫁接的结果与I型嫁接相似, 但根系对叶片衰老的作用未表现出绝对优势。从根-冠-根通讯的角度对叶片衰老的根-冠互作类型多样性进行了讨论。
Researches and practices associated with grafting indicate that the diversity of root-shoot interaction is universal in different physiological processes of plant. Our previous grafting study with CCRI41, an early senescence cotton cultivar, and SCRC22, a late senescence cultivar as materials found that shoot played a major role in mediating leaf senescence of cotton. In the present study, CCRI41 and another late senescence cotton cultivar CCRI49 were used to do standard grafting (I-grafting, one scion grafted onto one rootstock) and Y-grafting (two scions grafted onto one rootstock). Leaf senescence of grafts was induced by low potassium (K; 0.03 mmol L-1). Contrary to the grafting combination of CCRI41 and SCRC22, the combination of CCRI41 and CCRI49 showed that the role of root was more important than that of shoot for leaf senescence, as characterized by SPAD reading. Accordingly, the concentration of ZR+Z or iPA+iP in roots and the youngest fully expanded leaf and their delivery rate in xylem sap regarding I-grafting combinations with CCRI41 as rootstock were significantly lower than those with CCRI49 as rootstock in most of situations. However, the results of free ABA level were exactly opposite. With respect to the Y-grafting of CCRI41 and CCRI49, the results were similar to those of I-grafting, whereas the role of root was not absolutely predominant any more. The mechanism for different root-shoot interactions in cotton leaf senescence was discussed from the perspective of root-shoot-root distance communication.
叶片衰老是其发育的最后一个阶段, 被认为是一种程序性细胞死亡(PCD), 对植株将成熟叶片中积累的物质分解并运送至其他生长旺盛的部位进行再利用非常关键[1, 2]。棉花熟相包括正常成熟、早衰和贪青晚熟3种[3], 主要根据叶片衰老进程和吐絮速率的关系划分, 对棉花生产具有重要的指导意义。
叶片衰老受植物激素的调控, 细胞分裂素(CK)一般可以延缓叶片衰老[4, 5], 而脱落酸(ABA)往往对叶片衰老起促进作用[6, 7, 8]。CK在植物根尖合成是一个普遍接受的事实[9, 10, 11, 12, 13, 14], 根系合成的CK可以通过木质部运输至叶片, 并调节叶片衰老[15, 16, 17]。根系也是ABA的重要合成部位[18], 之后ABA在蒸腾流的驱动下通过木质部导管向叶片中输送[19]。嫁接试验也表明根系是棉花[20]和番茄[21, 22, 23]叶片衰老的主要决定部位。然而, 也有试验得到不同的结果。豌豆[24]和拟南芥[25]的地上部具有调节根系输出CK的能力, Gan和Amasino[5]及Faiss等[26]发现, 过量合成CK的砧木未能延缓接穗叶片的衰老。Koeslin-Findeklee等[27]的嫁接试验表明, 冬油菜在氮饥饿条件下的持绿表型主要受叶片控制。衰老较快的棉花品种中棉所41和衰老较慢的品种鲁棉研22互相嫁接时发现, 地上部是决定棉花叶片衰老(缺钾诱导)的主要部位[28, 29]。引人注意的是, 中棉所41与另一个衰老较慢的棉花品种中棉所49互相嫁接时, 根系对叶片衰老的决定作用又占据了优势[30]。可见, 叶片衰老的根-冠互作类型存在多样性, 但其在根-冠-根之间的通讯机制尚不明确。
本文采用单砧木单接穗(I型)和单砧木双接穗(Y型)嫁接方式比较系统地研究了中棉所41与中棉所49互相嫁接的根系、木质部汁液和叶片中的活性CK及游离态ABA的水平, 并讨论了根系为主型和地上部为主型调控叶片衰老的根-冠-根通讯机制。研究结果可为揭示叶片衰老的生理机制提供部分依据, 并可为棉花生产中制定防早衰措施提供一定指导。
供试棉花品种中棉所41 (衰老较快, 以下简称中41或41)和中棉所49 (衰老较慢, 以下简称中49或49)由中国农业科学院棉花研究所提供。
试验于中国农业大学光照培养室中进行, 光照强度为400 μ mol cm-2 s-1, 光照/黑暗时间为14 h/10 h, 昼/夜温度为(28± 2)℃/(22± 2)℃, 相对湿度为70%~80%。种子经10%双氧水消毒15 min清洗数遍后, 置去离子水中浸24 h, 露白后播于沙床, 出苗2 d后转移至K+浓度为0.1 mmol L-1的1/2改良Hoagland’ s营养液中培养, 营养液配方(mmol L-1) 为2.5 Ca(NO3)2, 1 MgSO4, 0.5 (NH4)H2PO4, 2× 10-4CuSO4, 1× 10-3 ZnSO4, 0.1 Fe Na EDTA, 2× 10-2H3BO3, 5× 10-6(NH4)6Mo7O24和1× 10-3 MnSO4。营养液培养所用容器为16 cm× 13 cm× 16 cm的塑料盒, 幼苗基部用海绵包裹, 以预先打好孔的聚乙烯泡沫板固定于塑料盆上, 每盒4株。每塑料盒装入营养液2.2 L, 每4 d更换营养液一次, 用气泵24 h通气。
砧木移入营养液中培养时播种接穗, 当砧木第1片真叶展开、接穗子叶完全展开时嫁接。参照李博等[31]的方法对2个品种进行I型(单砧木单接穗, 表示为接穗/砧木)和Y型[单砧木双接穗, 表示为(接穗+接穗)/砧木]嫁接。I型嫁接共4个处理, 分别为41/41、49/49、41/49、49/41; Y型嫁接的4个处理为(41+41)/41、(49+49)/49、(41+49)/41、(41+49)/49。随机区组排列, 重复4次, 每重复3盒, 每盒4株。
将嫁接苗置刺有若干小孔的保鲜袋内(注意避免叶片与保鲜袋直接接触), 放在光强为80~100 μ mol cm-2 s-1、昼夜温度为29℃/20℃的条件下缓苗, 5 d后揭掉保鲜袋, 7 d后移至400 μ mol cm-2 s-1光强下开始低钾胁迫(0.03 mmol L-1)诱导衰老, 以充足供钾(2.5 mmol L-1)为对照。
嫁接植株培养至七至八叶期时, 用SPAD-502叶绿素仪(浙江托普仪器有限公司, 中国杭州)测定各处理所有植株功能叶(倒四叶)叶绿素的相对含量, 每个叶片测量20点, 取平均值。
测完叶绿素含量后, 取下倒四叶用去离子水冲洗干净, 吸水纸吸干水分, 称取0.2 g, 置-40℃冰箱中待测。
取完叶片后分别在嫁接位点上部(接穗下胚轴)和嫁接位点下部(砧木下胚轴)采用压力室法[32]收集木质部汁液, 所用压力室为SEC 3005 plant Water Status Console (Soilmoisture Equipment Corp., CA, USA)。各处理2个部位同时收集, 每个部位收集4株, 重复3次。收集前用去离子水冲洗切面, 之后将干净的气门芯套紧在切断的下胚轴上。设置压力为3 bar, 收集时间为0.5 h。记录每株收集到的木质部汁液体积, 然后将同一处理的各株收集液混合。将一定体积收集液转入离心管放置在-80℃低温冰箱中冷冻48 h以上, 用冻干机(SIM FD5-6, LA, USA)冻干后贮存在-40℃待测。
选择未进行木质部汁液收集的代表性植株, 剪取2~3 cm长的侧根, 以去离子水冲洗并用吸水纸吸干水分, 剪碎混匀后称取0.2 g, 置于-40℃冰箱中待测。
取倒四叶和根系鲜重0.2 g, 冰浴下加入80% (v/v)甲醇溶液[含1 mmol L-1丁基羟基苯甲醚和1% (w/v) PVP]研磨, 4℃下提取4 h以上, 然后在4℃下离心15 min (4000转min-1), 取上清液用氮气吹干, 保存残留物于-40℃冰箱待测。
用样品稀释液(含NaCl 137 mmol L-1, KCl 2.7 mmol L-1, Na2HPO4 10 mmol L-1, KH2PO4 2 mmol L-1)溶解上述冻干的木质部汁液和氮气吹干的叶片及根系提取物, 采用间接酶联免疫吸附法(ELISA)测定样品中的游离态ABA和ZR(玉米素核苷)及iPA(异戊烯基腺嘌呤核苷)等活性CK含量, 单克隆抗体由中国农业大学作物化学控制研究中心制备[33]。由于ZR和iPA单克隆抗体分别与Z (玉米素)和iP (异戊烯基腺嘌呤)存在交叉反应, 因此实际测定结果为ZR+Z和iPA+iP的含量。
I型和Y型嫁接试验均经过3次以上重复, 各次结果趋势一致, 本文选用其中具有代表性的一次。利用SAS Ver.8 (SAS Institute Inc., Cary, NC, USA)进行方差分析, 用Duncan’ s法进行多重比较。
砧木和接穗效应是指某一品种在互相嫁接处理中作为砧木或接穗时与另一个品种自身嫁接的对比。叶片SPAD值、植物激素水平及接穗木质部汁液中植物激素水平与叶片衰老关系密切, 其砧木和接穗效应计算公式如下, 其中粗体标出的接穗为参与计算的接穗。
I型嫁接:
49砧木效应= (41/49-41/41)/(41/41)
49接穗效应= (49/41-41/41)/(41/41)
41砧木效应=(49/41-49/49)/(49/49)
41接穗效应=(41/49-49/49)/(49/49)
Y型嫁接:
49砧木效应
=[(41+49)/49-(41+41)× ½ /41]/[(41+41)× ½ /41]
49接穗效应
=[(41+49)/41-(41+41)/41]/[(41+41)/41]
41砧木效应
=[(41+49)/41-(49+49)× ½ /49]/[(49+49)× ½ /49]
41接穗效应
=[(41+49)/49-(49+49)/49]/[(49+49)/49]
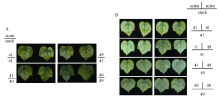
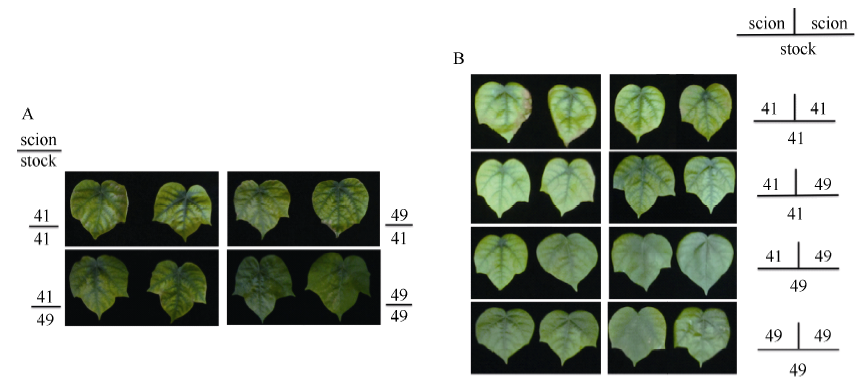
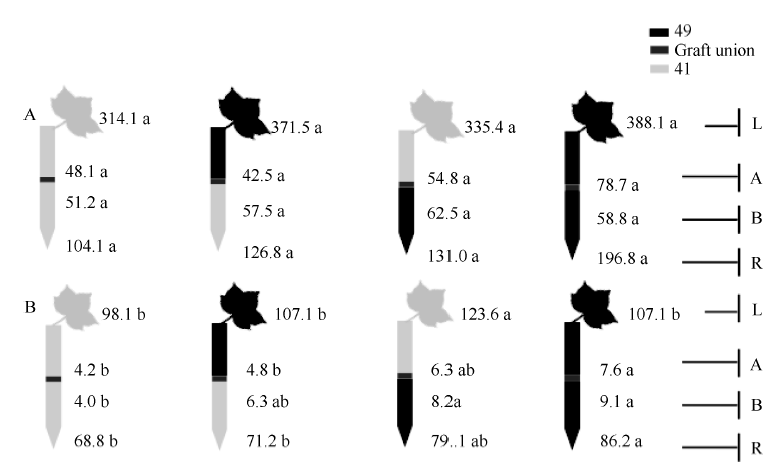
由图1和表1可知, 充足供钾条件下I型和Y型嫁接各处理接穗倒四叶的叶色和SPAD值无显著差异, 说明在正常条件下中41和中49的叶片衰老程度基本一致。低钾条件下, I型嫁接各处理倒4叶的SPAD平均值较充足供钾条件下降低了44%, Y型嫁接降低了40% (表1), 说明低钾胁迫下叶片出现衰老现象(图2)。此外, 低钾条件下以中41为砧木的嫁接处理倒四叶的SPAD值低于以中49为砧木的处理, 其中I型嫁接达到显著水平, Y型嫁接差异不显著, 各处理叶片衰老程度(图2)与SPAD值结果一致。
 | 图1 高钾条件下(2.5 mmol L-1) I型(A)和Y型(B)嫁接植株的倒四叶Fig. 1 The youngest fully expanded leaf of I grafts (A) and Y grafts (B) under sufficient potassium (2.5 mmol L-1) |
 | 图2 低钾条件下(0.03 mmol L-1) I型(A)和Y型(B)嫁接植株的倒四叶Fig. 2 The youngest fully expanded leaf of I grafts (A) and Y grafts (B) under potassium deficiency (0.03 mmol L-1) |
| 表1 供钾水平对I型和Y型嫁接各处理倒四叶SPAD值的影响 Table 1 Effect of potassium (K) supply on SPAD value of the youngest fully expanded leaf of cotton standard grafts (scion/rootstock) and Y grafts (scion+scion/rootstock) |
低钾条件下, 中41和中49在I型嫁接中对叶片SPAD值的砧木效应和接穗效应均达到显著水平, 其中中41效应为负值, 中49效应为正值。2个品种的砧木效应均大于接穗效应(表2), 反映出砧木对叶片衰老的决定作用大于接穗。Y型嫁接中2个品种的砧木效应和接穗效应较I型嫁接中为低, 且均不显著, 但砧木效应仍大于接穗效应。
| 表2 低钾条件下(0.03 mmol L-1)不同嫁接方式倒四叶SPAD值的砧木和接穗调控效应 Table 2 Effects of rootstock and scion on SPAD value of the youngest fully expanded leaf under low K+ (0.03 mmol L-1) condition (%) |
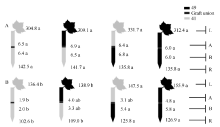
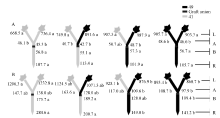
与充足供钾相比, 低钾胁迫下各嫁接处理倒4叶的平均ZR+Z和iPA+iP浓度分别降低67%和54%, 而ABA浓度增加171% (图3~图5), 这与叶片的衰老表型有关。
 | 图3 供钾水平对I型嫁接各处理根系(R)和倒四叶(L)的ZR+Z浓度(ng g-1 FW)及嫁接位点上部(A)和下部(B)木质部汁液ZR+Z流量(ng plant-1 h-1)的影响 A: 充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 3 Effect of K deficiency on ZR+Z level of cotton standard grafts (scion/rootstock) A: sufficient K (2.5 mmol L-1). B: low K (0.03 mmol L-1). The ZR+Z concentration (ng g-1FW) in roots (R) and the youngest fully expanded leaf (L), and ZR+Z delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4). |
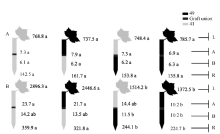
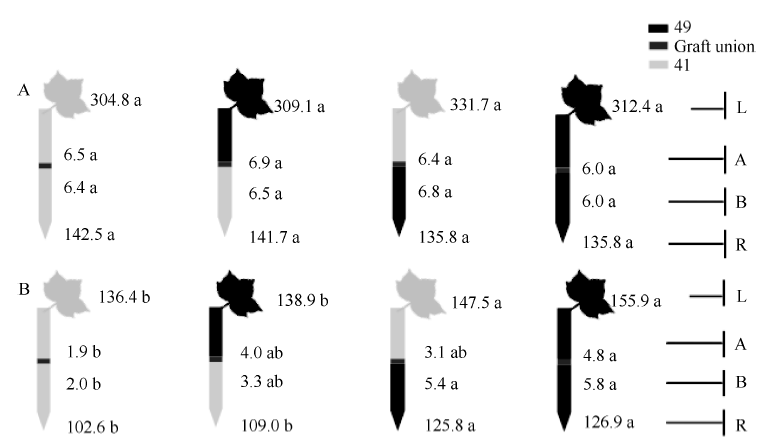 | 图4 供钾水平对I型嫁接各处理根系(R)和倒四叶(L)的iPA+iP浓度(ng g-1 FW)及嫁接位点上(A)和下部(B)木质部汁液iPA+iP流量(ng plant-1 h-1)的影响 A: 充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 4 Effect of K deficiency on iPA+iP level of cotton standard grafts (scion/rootstock) A: sufficient K (2.5 mmol L-1). B: low K (0.03 mmol L-1). The iPA+iP concentration (ng g-1FW) in roots (R) and the youngest fully expanded leaf (L), and iPA+iP delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4). |
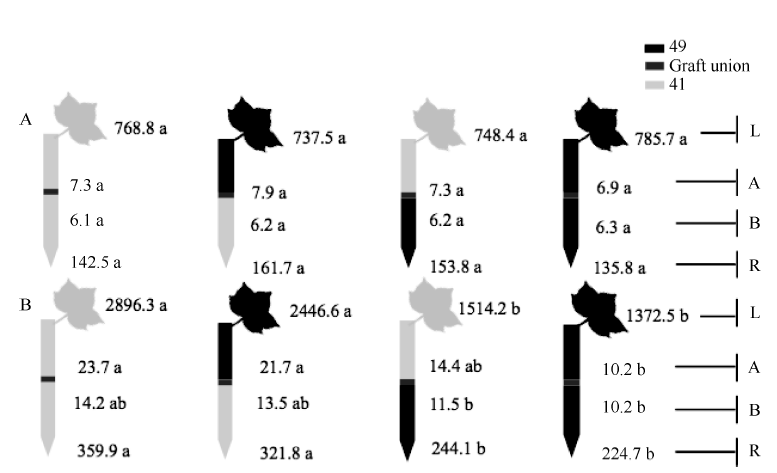 | 图5 供钾水平对I型嫁接各处理根系(R)和倒四叶(L)的ABA浓度(ng g-1 FW)及嫁接位点上部(A)和下部(B)木质部汁液ABA流量(ng plant-1 h-1)的影响 A: 充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 5 Effect of K deficiency on ABA level of cotton standard grafts (scion/rootstock) A: sufficient K (2.5 mmol L-1). B: low K (0.03 mmol L-1). The ABA concentration (ng g-1 FW) in roots (R) and the youngest fully expanded leaf (L), and ABA delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4). |
从图3-A、图4-A和图5-A可知, 充足供钾条件下各处理根系和叶片中的ZR+Z、iPA+iP和ABA浓度及砧木和接穗木质部汁液中几种植物激素的流量均无显著差异。但在低钾胁迫下, 以中41为砧木的处理, 其根系和叶片中的ZR+Z和iPA+iP浓度及砧木和接穗木质部汁液中的ZR+Z和iPA+iP流量在绝大多数情况下显著低于以中49为砧木的处理(图3-B、图4-B), ABA的结果则相反(图5-B)。
从表3可看出, 中41砧木和接穗对ZR+Z和iPA+iP的效应为负值、对ABA的效应为正值, 而中49砧木和接穗对ZR+Z和iPA+iP的效应为正值、对ABA的效应为负值。砧木效应均大于接穗效应, 且砧木对叶片激素浓度的效应均达到显著水平, 而接穗效应均不显著(图3-B、图4-B、图5-B)。
| 表3 低钾条件下(0.03 mmol L-1) I型嫁接倒四叶和接穗木质部汁液中激素水平的砧木和接穗效应 Table 3 Effect of rootstock and scion on phytohormone levels in the youngest fully expanded leaf and the xylem sap above graftunion for standard graftings (one scion/one rootstock) under low K+ (0.03 mmol L-1) condition (%) |
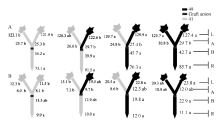
与充足供钾相比, 低钾胁迫下各嫁接处理倒四叶的平均ZR+Z和iPA+iP浓度分别降低86%和51%, 而ABA浓度增加21% (图6~图8)。
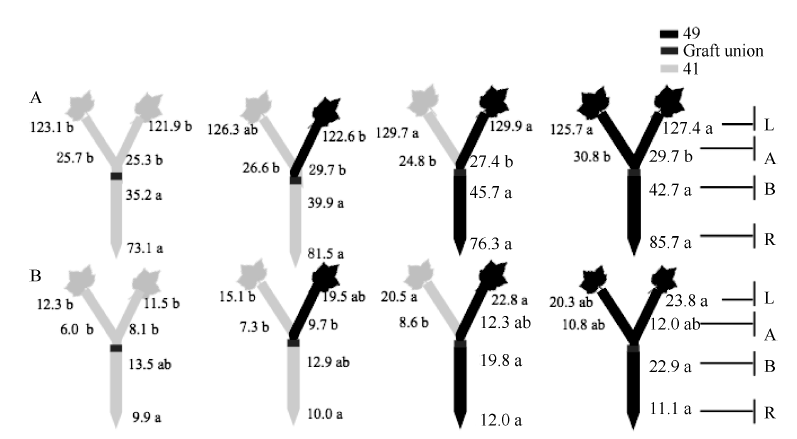 | 图6 供钾水平对Y型嫁接处理根系(R)和倒四叶(L)的ZR+Z浓度(ng g-1 FW)及嫁接位点上部(A)和下部(B)木质部汁液ZR+Z流量 (ng plant-1 h-1)的影响 A:充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 6 Effect of K deficiency on ZR+Z level of cotton Y grafts (two scions/rootstock) A: sufficient K (2.5 mmol L-1); B: low K (0.03 mmol L-1). The ZR+Z concentration (ng g-1 FW) in roots (R) and the youngest fully expanded leaf (L), and ZR+Z delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4). |
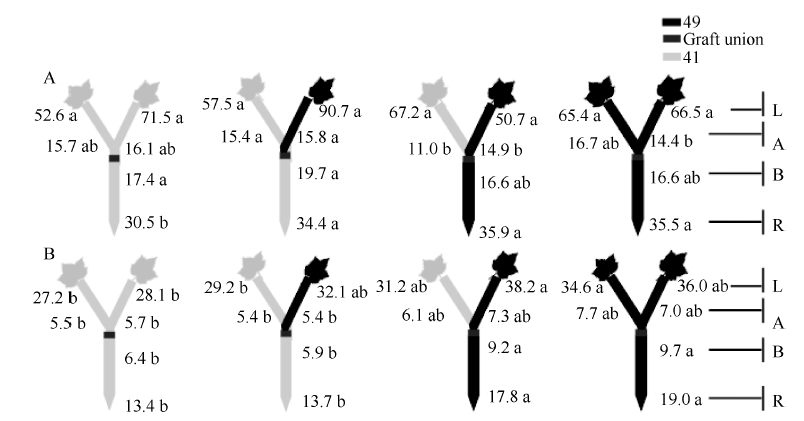 | 图7 供钾水平对Y型嫁接各处理根系(R)和倒四叶(L)的iPA+iP浓度(ng g-1 FW)及嫁接位点上部(A)和下部(B)木质部汁液iPA+iP流量(ng plant-1 h-1)的影响 A:充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 7 Effect of K deficiency on iPA+iP level of cotton Y grafts (two scions/rootstock) A: sufficient K (2.5 mmol L-1). B: low K (0.03 mmol L-1). The iPA+iP concentration (ng g-1 FW) in roots (R) and the youngest fully expanded leaf (L), and iPA+iP delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4). |
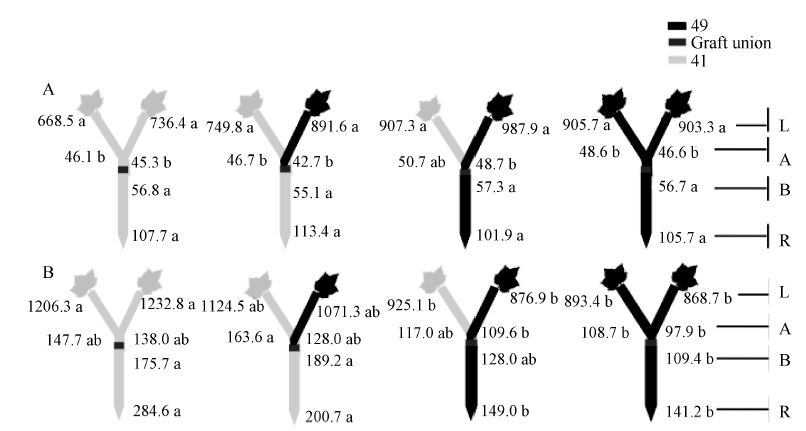 | 图8 供钾水平对Y型嫁接各处理根系(R)和倒四叶(L)的ABA浓度(ng g-1 FW)及嫁接位点上部(A)和下部(B)木质部汁液ABA流量(ng plant-1 h-1)的影响 A: 充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 8 Effect of K deficiency on ABA level of cotton Y grafts (two scions/rootstock) A: sufficient K (2.5 mmol L-1); B: low K (0.03 mmol L-1). The ABA concentration (ng g-1 FW) in roots (R) and the youngest fully expanded leaf (L), and ABA delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4). |
与I型嫁接相同, 充足供钾条件下Y型嫁接各处理根系和叶片中的ZR+Z、iPA+iP和ABA浓度及砧木和接穗木质部汁液中几种激素的流量均无显著差异(图6-A、图7-A、图8-A; 接穗木质部汁液中的iPA+iP流量除外)。在低钾胁迫下, 以中41为砧木的处理, 其根系和叶片中的ZR+Z和iPA+iP浓度及砧木和接穗木质部汁液中的ZR+Z和iPA+iP流量低于以中49为砧木的处理(图6-B、图7-B), ABA的结果则相反(图8-B), 但相当多情况下差异未达到显著水平。
当比较低钾条件下的砧木和接穗效应时(表4), 发现Y型嫁接的砧木和接穗效应大多数情况下较I型嫁接为低, 且中49砧木对中41接穗木质部汁液中iPA+iP流量的调控效应不再为正值、对ABA流量的调控效应不再为负值。此外, 仅中49砧木对中41接穗叶片ZR+Z和ABA浓度的调控效应达到显著水平, 其他砧木和接穗效应均不显著(图6-B、图7-B、图8-B)。
| 表4 低钾条件下(0.03 mmol L-1) Y型嫁接倒四叶和接穗木质部汁液中激素水平的砧木和接穗效应 Table 4 Effect of rootstock and scion on phytohormone levels in the youngest fully expanded leaf and the xylem sap above graf tunion for Y graftings (two scion/one rootstock) under low K+ (0.03 mmol L-1) condition (%) |
Li等[28]和Wang等[29]将衰老较快棉花品种中41与衰老较慢品种鲁棉研22 (以下简称鲁22或22)进行I型、Y型和A型(单接穗双砧木)嫁接, 发现地上部对叶片衰老的决定作用大于根系。而本文结果显示, 将中41与另一个衰老较慢的棉花品种中49进行I型嫁接, 根系对叶片衰老(以SPAD值表征)的决定作用大于地上部; Y型嫁接的结果虽然不如I型嫁接明显, 但表现出相同的趋势。
采用嫁接方式提高果树和蔬菜对生物逆境和非生物逆境的抗性是一种常用措施, 其根-冠互作也表现出不同的类型, 如有的抗性砧木对某些基因型接穗具有改善和提高作用, 但对其他基因型接穗效果不佳[34, 35, 36, 37, 38, 39, 40]。可见, 根-冠互作类型的多样性在植物的不同生理过程中普遍存在。
叶片中的CK和ABA分别对衰老起正调控和负调控作用[4, 5, 6, 7, 8]。本文所测定的ZR+Z和iPA+iP为活性形式的CK[41], 所测定的ABA为游离态ABA。从理论上讲, 叶片中的活性CK和游离态ABA水平主要由三方面因素决定: 第一方面是根系的合成能力和输出能力; 第二方面是茎组织中的合成、活化、降解、钝化反应及与木质部之间的横向转移; 第三方面是叶片内的合成、活化、降解和钝化反应。
根系合成的CK[9, 10, 11, 12, 13, 14]和ABA[18]在蒸腾拉力的作用下向地上部移动[11, 19, 42, 43], 但这并不是一个完全被动的过程。以CK为例, 由根系向地上部输送的CK速率受到根系细胞保留CK能力的调节[44]。根系中的CK在装载入木质部导管之前, 必须进行跨膜运输。研究表明嘌呤透性酶(PUP)[45]、均衡核苷转运蛋白(ENT)[46]、AtABCG14 (ATP-binding cassette G14)[14]可介导这一过程, 这些载体的数量和活性也直接调控着根系输出CK的能力。此外, 位于根系木质部装载位点附件的CK氧化酶/脱氢酶(CKX)也会通过降解代谢[47]调节根系输出的活性CK数量。
一些直接和间接证据显示, CK和ABA在木质部运输过程中发生着活跃的变化。作者未发表数据表明, 棉花下胚轴韧皮部中的CK合成基因GhIPT (与AtIPT3同源性为55%)和运输基因GhPUP (与AtPUP2同源性为55%)表达量较高, 木质部中的ABA合成基因GhNCED (与AtNCED3同源性为66%)、活化基因GhBG (与AtBG1同源性为52%)和运输基因GhABCG (与AtABCG25同源性为64%)表达量较高[48]。而CK[49]和ABA[50]均可在茎组织中横向转移, 即从茎薄壁细胞转移到木质部, 或从木质部转移到薄壁细胞。Zhang等[51]在羽扇豆上的同位素示踪试验结果表明, [3H]ZR和[3H]DHZR通过蒸腾流被吸收后, 甚至可迅速(以核苷和/或相应的核苷酸形式)从木质部横向移动到茎表皮组织。木质部薄壁细胞中的ABA可能因木质部汁液中较高的pH向其中富集[52]。切花玫瑰叶柄木质部汁液中的ABA浓度与ABA传递速率之间缺乏联系[53], 也表明木质部汁液中的ABA浓度在运输途径中受到调节。
大量研究结果已经表明, 叶片具有合成、降解、活化和钝化CK的能力。Miyawaki等[54]和Takei等[55]为叶片具有合成CK的能力提供了证据, Nová ková 等[56]发现叶片中的CK水平受到CKX及β -葡糖苷酶(参与CK O-葡糖苷的水解)的精细调节。Koeslin- Findeklee等[27]的试验表明, 氮饥饿下早衰的冬油菜品种其叶片中以活性CK的钝化反应为主, 而持绿性品种的叶片中以CK的合成、活化和对活性CK的感知及响应为主。叶片也具有合成ABA[57]和钝化ABA (在ABA葡糖基转移酶的作用下形成ABA葡萄糖酯[58])的能力。McAdam和Brodribb[59]最近的研究表明, 在VPD (蒸气压差)变化时, 4种被子植物中的3种其叶片ABA含量显著升高, 而这种升高与叶片的合成能力有关。此外有研究表明, 切花玫瑰叶片气孔敏感性(对高的相对湿度)的基因型差异也主要由叶片内的ABA平衡/稳态决定[53]。
CK和ABA虽然可作为信息物质调控植物诸多发育过程, 但其在根系、茎组织和叶片中的合成、代谢和运输可能受到其他根-冠信号和/或冠-根信号的调控。近些年关于植物的根-冠和冠-根长距离通讯及根-冠-根通讯回路研究取得了一些重要进展。Notaguchi和Okamoto[60]在最近的综述中提出, 当土壤环境变化时, 信号分子首先在根系的某处或整个根系中产生, 然后通过木质部向地上部每一个成熟叶片中(很可能是小叶脉中)传播; 再次, 信号分子从木质部运输到韧皮部, 被位于韧皮部细胞中的受体感知后发出二级信号; 最后, 二级信号分子在韧皮部汁液中输送到库器官, 包括自上而下输送到根系。豆科植物根瘤形成[61, 62, 63]和植物对低氮(N)响应[64]的根-冠-根通讯回路目前研究的比较清楚。
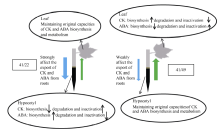
就叶片衰老而言, 41/22和41/49分别代表地上部主导型和根系主导型根-冠互作类型。如图9所示, 鲁22和中49发出的根-冠信号可能不同, 因此中41叶片内的受体感知到二者的信号后产生不同的响应, 如感知到鲁22的信号后可能倾向于维持原有的CK和ABA合成、代谢能力; 但感知到中49的信号后可能CK合成、活化能力提高和/或降解、钝化能力降低, ABA的合成、活化能力则降低和/或降解、钝化能力提高, 从而使ZR+Z、iPA+iP和ABA浓度向中49的水平靠近。此外, 中41叶片感知到鲁22和中49砧木的根-冠信号后可能形成不同种类或不同强度的冠-根信号, 从而引发不同的反应。如在41/22嫁接体中, 冠-根信号对鲁22砧木的CK和ABA输出能力和/或茎组织中的CK和ABA合成、代谢及横向运输具有较强的调节作用, 使之接近中41的水平; 而在41/49嫁接体中, 冠-根信号向下传递到中49砧木后倾向于维持砧木原有的CK和ABA输出能力和/或木质部运输过程。
 | 图9 棉花叶片衰老中不同根-冠互作类型的根-冠-根通讯机制 41: 中棉所41; 49: 中棉所49; 22: 鲁棉研22; 41/22: 地上部为主型; 41/49: 根系为主型; 绿色上行箭头: 根-冠信号; 蓝色下行箭头: 冠-根信号; 箭头粗细代表信号强弱。Fig. 9 The diagram of root-shoot-root communication for different interactions between root and shoot in cotton leaf senescence induced by low potassium (0.03 mmol L-1) stress 41: CCRI41; 49: CCRI49; 22: SCRC22; 41/22: the role of shoot is more important; 41/49: the role of root is more important. Green up arrow: the signal of root-shoot. Blue down arrow: the signal of shoot-root. The thickness of arrow indicates signal strength. |
中41与鲁22无论采用I型、Y型还是A型嫁接方式, 均表现为地上部对叶片衰老起主要作用[29]。中41与中49的I型嫁接表现出根系起主要作用的方式, 但Y型嫁接中根系的作用不再具有明显优势, 仅略大于地上部的作用。推测原因如下, 中41与中49的Y型互相嫁接体中[(41+49)/41、(41+49)/49], 在另一个与砧木品种相同的接穗存在的情况下, 与砧木品种不同的接穗接收到的根-冠信号可能不同于二者之间I型互相嫁接的根-冠信号, 致使该接穗叶片中发生不同于I型互相嫁接的变化, 即叶片的CK和ABA合成、代谢反应不再发生类似于砧木品种的变化, 或变化强度降低。此外, 该接穗中的受体感知到砧木的根-冠信号后向下传递的冠-根信号可能也不同于I型嫁接, 因而对其下胚轴中的CK和ABA合成、代谢及横向运输表现出一定程度的调节作用, 类似于中41和鲁22组合[29]。
尽管我们初步描绘了棉花叶片衰老过程中根-冠-根的通讯回路, 但对其信号物质并不清楚。已知植物韧皮部汁液成分比较复杂, 含有糖类、脂类、氨基酸、多肽、蛋白质、编码和非编码RNAs、矿质养分和植物激素等一系列物质[65, 66, 67], 木质部汁液成分比较简单, 主要含有矿质养分、多肽、蛋白质和植物激素[67, 68, 69]。未来需要在众多的木质部汁液和韧皮部汁液成分中鉴定与棉花叶片衰老有关的根-冠信号和冠-根信号, 这将有助于人们采取针对性措施对棉花衰老予以调节。
以中41为砧木的处理, 其叶片中ZR+Z、iPA+iP的含量低于以中49为砧木的处理, ABA含量则相反, 表明根系对叶片衰老的作用较大。据分析, 中41和中49互相嫁接体(41/49和49/41)接穗叶片中的受体感知到来自砧木的缺钾信号后, 发出的冠-根信号对砧木输出CK和ABA的能力和/或木质部中CK和ABA的运输过程影响较小; 接穗叶片中CK和ABA的合成、代谢则可能发生类似于砧木品种的变化。
致谢: 感谢课题组王晔博士在试验过程中提供的帮助。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|