

不同小麦品种粒重和蛋白质含量的穗粒位效应分析
[李豪圣1  , 曹新有
, 曹新有1 , 宋健民1 , 刘鹏2 , 程敦公1 , 刘爱峰1 , 王灿国1 , 刘建军1, * , 孙正娟1 ]
, 曹新有, 孙正娟|
|


第一作者联系方式: E-mail: lihaosheng810@163.com
小麦籽粒的发育存在时空差异, 不同穗粒位的粒重和蛋白质产量也存在差异, 剖析粒重和籽粒蛋白质含量的穗粒位效应, 有助于深入了解小麦产量和品质的形成机制。于2009—2010和2010—2011小麦生长季进行大田试验, 选用3种类型4个品种, 分析了不同穗粒位的粒重、蛋白质积累和蛋白质含量的动态变化。结果表明, 粒重和蛋白质积累量的穗粒位间变异大于年份(环境)间变异和基因型间变异; 蛋白质含量的年份间变异大于基因型间变异和穗粒位间变异, 而成熟期穗粒位间变异最大。大粒品种易受环境影响, 小粒品种比较稳定。优质面包小麦品种开花后各时期的籽粒蛋白质含量普遍高于中筋小麦, 但不同时期、不同年份差异较大。开花后各时期, 强势粒的粒重、蛋白质积累量和蛋白质含量显著大于弱势粒, 中部籽粒显著大于上部和下部籽粒; 随着灌浆进程穗中部与下部籽粒的差异变小, 至开花后36 d时, 中部和下部籽粒的蛋白质含量无显著差异。随籽粒灌浆进程, 不同品种各穗粒位的粒重和蛋白质积累均呈“慢-快-慢”的“S”型曲线变化, 蛋白质含量均呈“高-低-高”的“V”型曲线变化, 灌浆后期, 中部和下部强势粒以及下部弱势粒的蛋白质含量增长速度明显快于其他穗粒位籽粒。粒重最大生长速率出现在开花后18~21 d, 快速增重时期为开花后12~26 d; 籽粒蛋白质最大积累速率出现在开花后21~24 d, 快速积累时期为开花后13~32 d。根据本研究结果, 我们认为高产优质小麦品种的特征是籽粒不宜过大, 小花位粒数不宜过多, 且中、下部籽粒较多, 开花后13~26 d灌浆速率快。
, CAO Xin-You, SUN Zheng-JuanWheat seeds are developed in a spatial-temporal order resulting in different grain weights and protein contents in different positions of spike. Dissecting the position effect of grain weight and protein content is helpful to go into the mechanism of yield and quality formation. This study was carried out in the 2009-2010 and 2010-2011 winter wheat seasons with four varieties in three quality types. The dynamic changes of grain weight, protein accumulation and protein content were measured according to spikelet-grain position. Variations of grain weight and protein accumulation affected by spikelet-grain position were greater than those by environment (year) or genotype. Variation of protein content affected by environment was larger than that by genotype or spikelet-grain position, however, the spikelet-grain position was the first affecting factor at maturity stage. Large-grain variety was more sensitive to environment, whereas, small-grain variety was relatively stable. During grain filling, the protein content of strong-gluten wheat was higher than that of medium-gluten wheat, and influenced greatly by filling stage and environment. The single grain weight, protein content, and protein accumulation of superior grains were significantly higher than those of inferior grains during the grain-filling period, and the mid-position grains were significantly superior to the upper- and lower-position grains. However, the difference between mid- and lower-position grains became smaller with the filling process, until no significant difference for protein content at maturity stage (36 days after flowering). For each genotype or spikelet-grain position, the dynamics of single grain weight and protein accumulation showed an “S” curve (slow-fast-slow) and the dynamics of protein content showed a “V” curve (high-low-high). During the late filling stage, the supervisor grains in mid- and lower-positions, as well as inferior grains in lower-position had faster increase of protein content than grains in other positions. For grain weight, the maximum growth rate appeared at 18-21 days after flowering, and the rapid growth period appeared at 12-26 days after flowering. For grain protein, the maximum accumulation rate appeared at 21-24 days after flowering, and the fast accumulation period was from 13 to 32 days after flowering. It is concluded that moderate grain size and floret number are important for high-yield and high-quality wheat breeding, and an appropriate increase of the mid- and lower-position grains may accelerate grain-filling speed at 13-26 days after flowering.
禾谷类作物小穗发育存在时空顺序, 因而不同部位籽粒的结实特性和籽粒性状存在差异。在水稻上, Nagato[1]首次提出了不同部位籽粒灌浆存在差异, 认为早开的花为强势粒, 迟开的花为弱势粒。籽粒在穗部的位置决定了小麦穗部籽粒结实与物质积累的空间分布特征[2], 小麦穗部粒重的分布具有近中优势, 即中部小穗粒重具有明显的优势[3], 其次是下部小穗, 而上部小穗粒重较低[4, 5]。不同粒位的粒重分布比较复杂, 同一小穗上, 粒重分布重心随结实率的不同而不同[6, 7]。屈会娟等[8]研究发现, 无论小穗结实粒数如何变化, 各小穗第1、第2粒位的粒重均显著大于其他粒位的粒重; 黄开红等[9]也报道, 小麦不同粒位的粒重都有一个共同的变化趋势, 即以穗轴中部小穗、小穗基部粒位粒重最高, 以此为中心向穗轴两端与小穗上方优势逐渐减弱。左毅等[10]以不同筋力小麦品种为试验材料, 发现强筯和弱筋品种的籽粒蛋白质含量均表现为穗下部> 穗中部> 穗上部, 而中筋品种则表现为穗中部> 穗下部> 穗上部。刘晓冰和李文雄[11]认为, 不同穗位籽粒蛋白质积累相对含量的顺序为中部粒> 下部粒> 上部粒, 相同穗位不同粒位蛋白质积累相对含量的顺序则是第2小花粒≥ 第1小花粒> 第3小花粒。吕淑芳等[12]研究认为, 小麦强势籽粒的蛋白质含量和积累量均显著高于弱势籽粒。然而, 已有的研究主要集中在小麦穗粒位粒重和蛋白质含量的差异, 而关于小麦穗粒位粒重和蛋白质含量变化动态差异的报道较少。本研究从分析小麦穗粒位籽粒形成过程中单粒粒重和蛋白质含量变化动态及蛋白质积累规律差异着手, 深化对小麦穗部籽粒发育变化规律的认识, 为高产优质品种的培育提供理论依据。
2009— 2010和2010— 2011连续两年小麦季在山东省农业科学院作物研究所试验农场(济南, 36.66° N, 116.99° E)进行大田试验。试验田土质为中壤土, 2009— 2010年度小麦播种前耕层(0~20 cm)土壤pH值为7.96, 含有机质1.80%、全氮0.12%、碱解氮114.32 mg kg-1、速效磷(P2O5) 18.2 mg kg-1、速效钾(K2O) 143.0 mg kg-1; 2010— 2011年度耕层土壤pH值为7.92, 含有机质1.76%、全氮0.12%、碱解氮102.1 mg kg-1、速效磷(P2O5) 17.6 mg kg-1、速效钾(K2O) 135.0 mg kg-1。两年土壤基础肥力基本一致。
采用单因素试验设计, 供试品种为济麦22 (常年粒重和蛋白质含量分别为41.1 mg 粒-1和13.0%, 属中粒、中筋小麦)、石4185 (常年粒重和蛋白质含量分别为39.7 mg 粒-1和13.1%, 属中粒、中筋小麦)、济麦20 (常年粒重、蛋白质含量和面包评分分别为37.6 mg 粒-1、14.3%和96.3, 属小粒、优质面包小麦)、济麦6487 (常年粒重、蛋白质含量和面包评分分别为44.2 mg 粒-1、15.1%和91.0, 属大粒、优质面包小麦)。分别于2009年10月9日和2010年10月2日采用奥地利小区播种机(WINTERSTEIGER)播种, 种植密度为225× 10-4 hm-2, 小区面积1.5 m × 4.0 m = 6.0 m2, 随机区组设计, 3次重复。播种前施复合肥(总养分≥ 25%, N、P2O5和K2O的比例为8:10:7) 750 kg hm-2, 拔节期追施尿素300 kg hm-2, 其他按大田常规管理。
抽穗开花期, 选择每个品种正常结实、小穗数一致、同天开花的穗挂牌标记120穗, 分别在花后6、12、18、24、30和36 d (成熟)取样, 每次取15~20穗(去除取样过程中落粒的)。取回的样穗, 于120℃下杀青30 min, 于65℃烘至恒重, 按不同穗粒位分别集中脱粒、称重, 换算单粒重, 用MRK-RETSCH旋风磨磨粉待测蛋白质含量。按照《NY/T2017-2011植物中氮、磷、钾的测定》方法测定全氮含量, 然后乘以系数5.7换算成粗蛋白质含量。
穗位的划分, 先确定中部穗, 即中间两侧各3个小穗, 中部穗以下小穗定义为下部穗(退化小穗数除外), 中部穗以上小穗定义为上部穗。粒位的划分, 每小穗第1、第2花位粒定义为强势粒, 剩余花位粒定义为弱势粒。
应用DPS7.05和SPSS17.0统计软件进行数据分析, 采用LSD法进行多重比较。参考戴国俊等[13]描述的方法计算Logistic生长方程中相关参数。
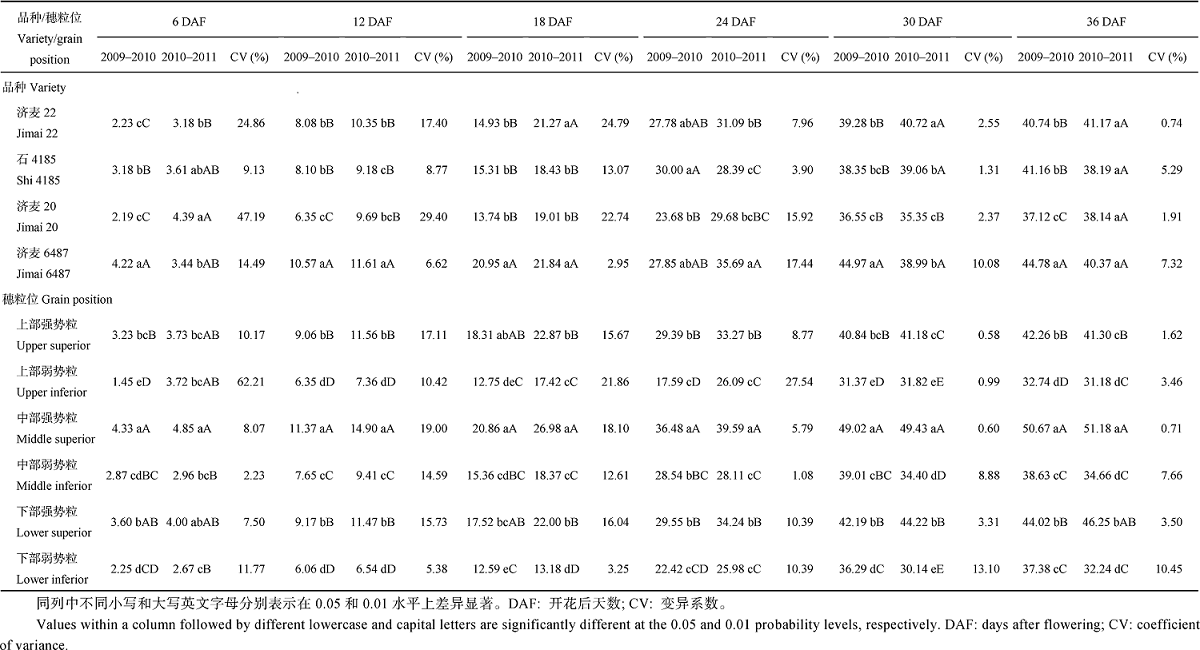
方差分析结果显示, 小麦开花后不同时期的粒重在年份间、品种间及穗粒位间存在差异, 除了成熟时(开花后36 d)的年份间差异达5%显著水平外, 其他均达到1%显著水平, 其中开花后6 d、18 d的年份间和开花后12、24、30和36 d的穗粒位间差异更为显著。可以看出, 粒重的穗粒位间变异大于年份间(环境)变异和基因型间变异, 而在成熟时, 基因型间变异大于年份间变异。年份× 品种互作除了对开花后36 d的影响达5%显著水平外, 其他均达到1%显著水平。年份× 穗粒位互作除了对开花后24 d的影响不显著外, 其他均达到5%或1%显著水平。品种× 穗粒位互作除了对开花后30 d的影响达1%显著水平外, 其他均未达到显著水平。年份× 品种× 穗粒位互作除了对开花后24 d的影响达5%显著水平外, 其他均未达到显著水平(表1)。
| 表1 小麦粒重、籽粒蛋白质积累量和含量变异的方差分析 Table 1 ANOVA of grain weight, protein accumulation, and protein content of wheat |
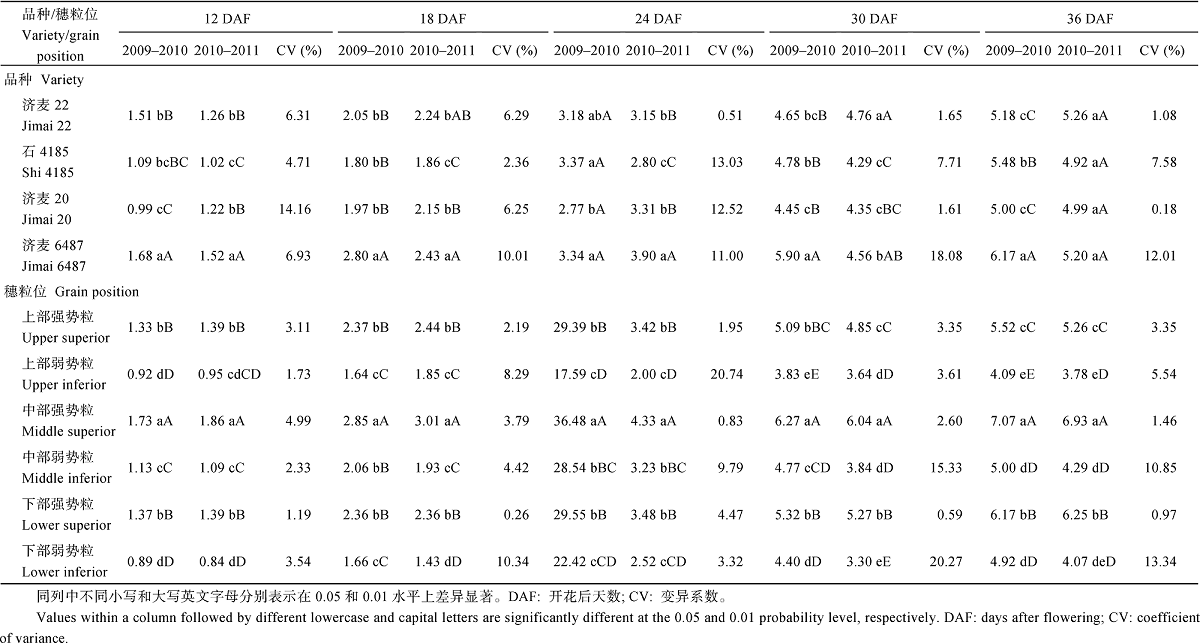
从蛋白质积累量来看, 除了开花后12、18和24 d的年份间外, 其他差异均达到1%显著水平, 其中穗粒位间差异更为显著。年份× 品种互作的影响均达到1%显著水平。年份× 穗粒位互作对开花后30 d蛋白质积累量的影响达到1%显著水平, 对开花后24 d和36 d的影响达到5%显著水平, 其他未达到显著水平。品种× 穗粒位互作对蛋白质积累量的影响, 与年份× 穗粒位互作一致。年份× 品种× 穗粒位互作除了对开花后30 d的影响达5%显著水平外, 其他均未达到显著水平。
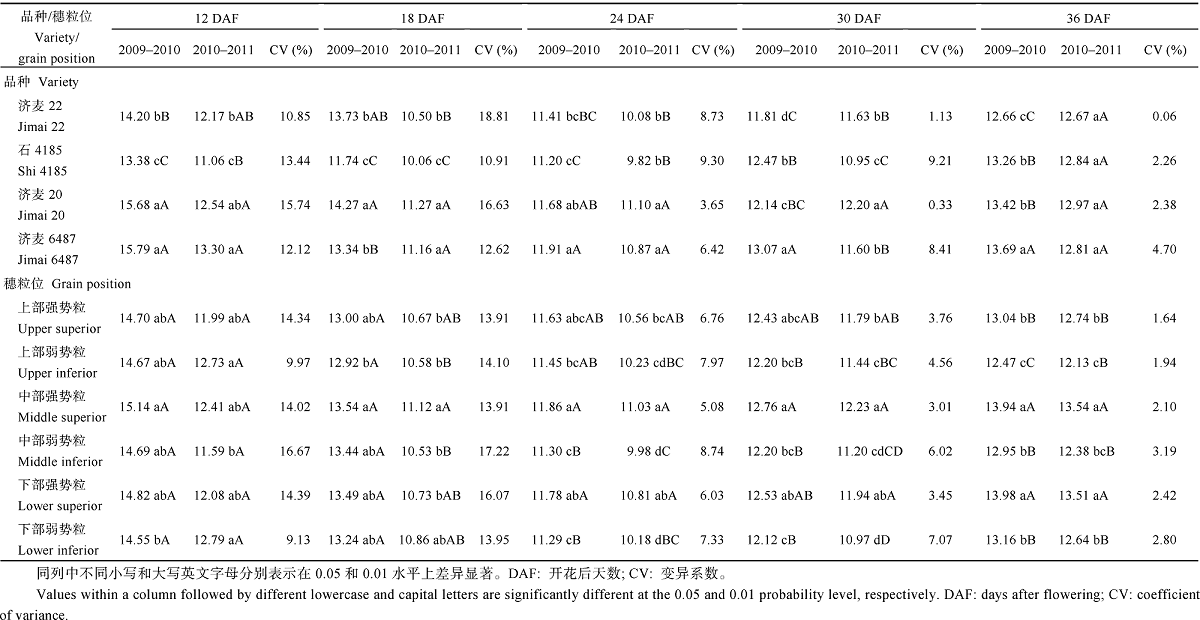
从籽粒蛋白质含量的差异表现来看, 除了开花后12 d的穗粒位间差异外, 年份间、品种间及穗粒位间的差异均达到5%或1%显著水平。可以看出, 除开花后36 d外, 籽粒蛋白质含量的年份间变异大于基因型间变异和穗粒位间变异, 而成熟期(开花后36 d)穗粒位间变异最大。年份× 品种互作对蛋白质含量的影响, 除了开花后12 d外, 其他均达到1%显著水平。年份× 穗粒位互作除了对开花后30 d的影响达1%显著水平外, 其他均无显著影响。品种× 穗粒位互作除了对开花后30 d的影响达到5%显著水平外, 其他均未达到显著水平。年份× 品种× 穗粒位互作对蛋白质含量的影响均未达到显著水平。
2.2.1 粒重差异 由表2可见, 大粒品种济麦6487, 在2009— 2010年度开花后各时期的粒重明显高于其他品种, 除开花后24 d外, 差异均达到了显著水平; 而2010— 2011年度优势不明显, 成熟期甚至小于中粒品种济麦22, 说明大粒品种粒重稳定性差。变异系数也证明了这一点, 在灌浆中后期(开花24 d后)济麦6487粒重变异系数最大。小粒品种济麦20, 在2009— 2010年度开花后各时期的粒重明显小于其他品种; 而2010— 2011年度, 各时期与中粒品种差异较小, 成熟期差异不显著。济麦20成熟期变异系数小, 说明小粒品种粒重稳定性较好。2个中粒品种, 在2009— 2010年度济麦22粒重除开花后30 d外, 均小于石4185, 但差异不显著(开花后6 d除外); 而2010— 2011年度, 除开花后6 d外, 济麦22粒重均大于石4185, 但成熟期差异不显著。说明粒重相近品种对环境的反应存在基因型差异。从成熟期变异系数可以看出, 济麦22粒重稳定性更好。
| 表2 小麦不同灌浆阶段的粒重变化 Table 2 Variation of grain weight at different filling stages(mg grain-1) |
开花后各时期强势粒粒重显著大于弱势粒, 中部籽粒显著大于上部和下部籽粒、灌浆后期与下部籽粒差异变小, 两年趋势基本一致。可以看出, 在整个籽粒灌浆过程中近中优势明显, 而灌浆后期下部优势又大于上部。不同穗粒位粒重的变异系数, 灌浆前中期总体大于后期, 弱势粒大于强势粒。说明小麦籽粒随着灌浆的进程不断自我调节, 而相对于弱势粒, 强势粒具有更大的优势。
2.2.2 蛋白质积累量差异 从表3可以看出, 不同基因型与穗粒位的籽粒蛋白质积累量差异与粒重相似。大粒品种济麦6487 (优质面包), 在2009— 2010年度开花后各时期的蛋白质积累量具有明显优势, 而在2010— 2011年度开花后30 d前的蛋白质积累量显著大于其他品种, 而在成熟期与其他品种差异未达到显著水平。小粒品种济麦20 (优质面包)在开花后各时期的蛋白质积累量, 与中筋品种相比没有显示优势, 成熟期低于济麦22。两个中粒、中筋品种, 济麦22开花后各时期的平均蛋白质积累量, 在大粒和小粒优质面包小麦品种之间, 2010— 2011年度灌浆后期(开花30 d后)略高于优质面包大粒品种; 而石4185在2009— 2010年度大于优质面包小粒品种, 2010— 2011年度小于优质面包小粒品种。说明济麦22蛋白质积累量较大, 且稳定性好于石4185。从变异系数来看, 济麦6487大、稳定性差, 济麦20虽然前期较大, 但后期(开花30 d后)稳定性最好。
| 表3 小麦不同灌浆阶段的籽粒蛋白质积累量变化 Table 3 Variation of protein accmulation in grains at different filling stages(mg grain-1) |
对于小麦不同穗粒位籽粒蛋白质积累量, 强势粒在1%水平下显著大于弱势粒, 穗中部和下部大于上部, 且这种优势随着灌浆的进程更加突出。两年趋势基本一致。不同穗粒位蛋白质积累量的变异系数, 弱势粒普遍大于强势粒, 尤其灌浆后期(开花30 d后), 说明相对于弱势粒, 灌浆后期强势粒积累蛋白质更加稳定。
2.2.3 蛋白质含量差异 多重比较分析表明(表4), 优质面包小麦品种济麦6487和济麦20, 在开花后各时期的蛋白质含量平均高于中筋小麦品种济麦22和石4185, 除2010— 2011年度成熟期外, 差异均达到显著水平。说明小麦蛋白质含量是面包烘烤品质的重要指标, 但也可以看出, 在不同时期、不同年份差异较大。从变异系数来看, 开花后24 d和30 d济麦20最小, 而成熟期济麦22最小、济麦6487最大, 说明济麦20和济麦22的品质比较稳定, 济麦6487品质稳定性差。
| 表4 小麦不同灌浆阶段的籽粒蛋白质含量变化 Table 4 Variation of protein content in grains at different filling stages(%) |
小麦不同穗粒位籽粒蛋白质含量, 强势粒大于弱势粒, 穗中部和下部大于上部, 且这种优势随着灌浆的进程更加明显, 在成熟期表现为中部强势粒≈ 下部强势粒> 上部强势粒> 下部弱势粒> 中部弱势粒> 上部弱势粒。两年趋势基本一致。不同穗粒位蛋白质含量的变异系数, 灌浆前中期总体大于后期, 灌浆后期(开花24 d后)弱势粒大于强势粒。
不同基因型与穗粒位的蛋白质含量均呈“ V” 型曲线变化, 开花后24 d达到最低点。在开花后30~36 d, 中部强势粒、下部强势粒和下部弱势粒回升速度明显快于其他穗粒位籽粒。这说明上述3个穗粒位在提高蛋白质含量上具有较大的潜质。
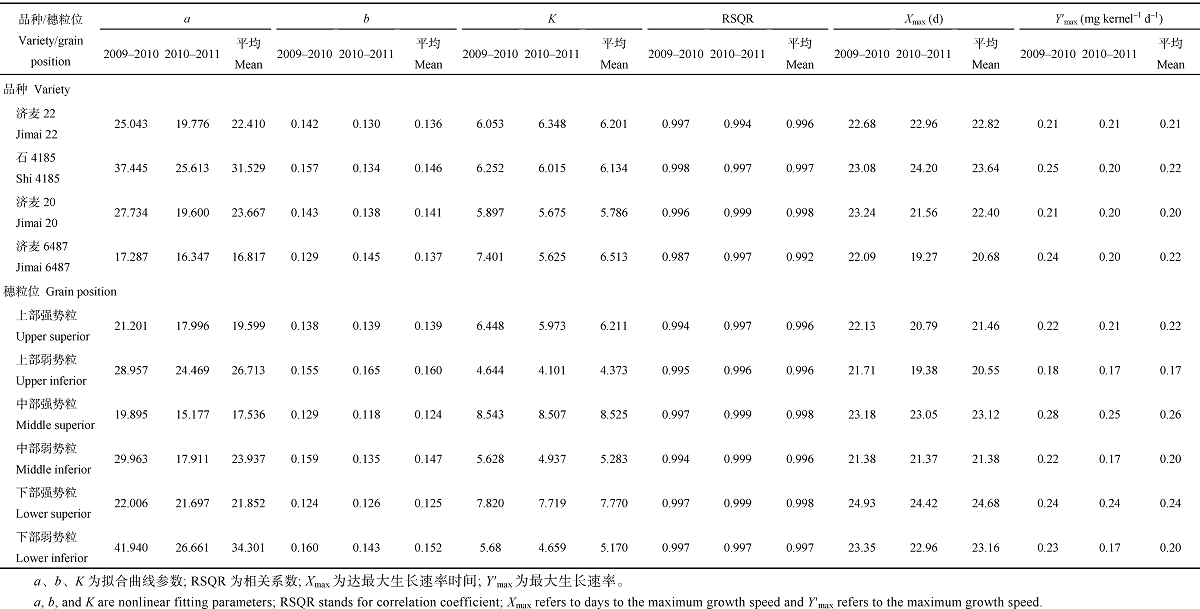
2.3.1 粒重动态 在籽粒灌浆过程中, 粒重呈 “ S” 型曲线变化, 可用Logistic生长方程Y= K/ (1+ae-bX)较好拟合, 相关系数均在0.99以上(表5)。根据方程的导数可求籽粒增长过程中“ 慢-快-慢” 转折点, 即曲线的拐点。从两年结果来看, 粒重最大生长速率一般出现在开花后18~21 d, 生长速率一般为1.9~2.2 mg 粒-1 g-1。4个不同基因型小麦品种中, 大粒品种济麦6487的粒重最大生长速率出现时间早、生长速率大, 小粒品种济麦20生长速率最小, 而中粒品种济麦22和石4185介于大粒和小粒两个品种之间。说明粒重越大, 最大生长速率越大。不同穗粒位粒重最大生长速率出现时间, 穗上部早于下部, 穗上部和下部强势粒早于弱势粒, 而中部强势粒略晚于弱势粒; 生长速率强势粒明显大于弱势粒。
| 表5 小麦粒重变化曲线的Logistic方程参数 Table 5 Parameters of Logistic equation for simulating grain weigth of wheat |
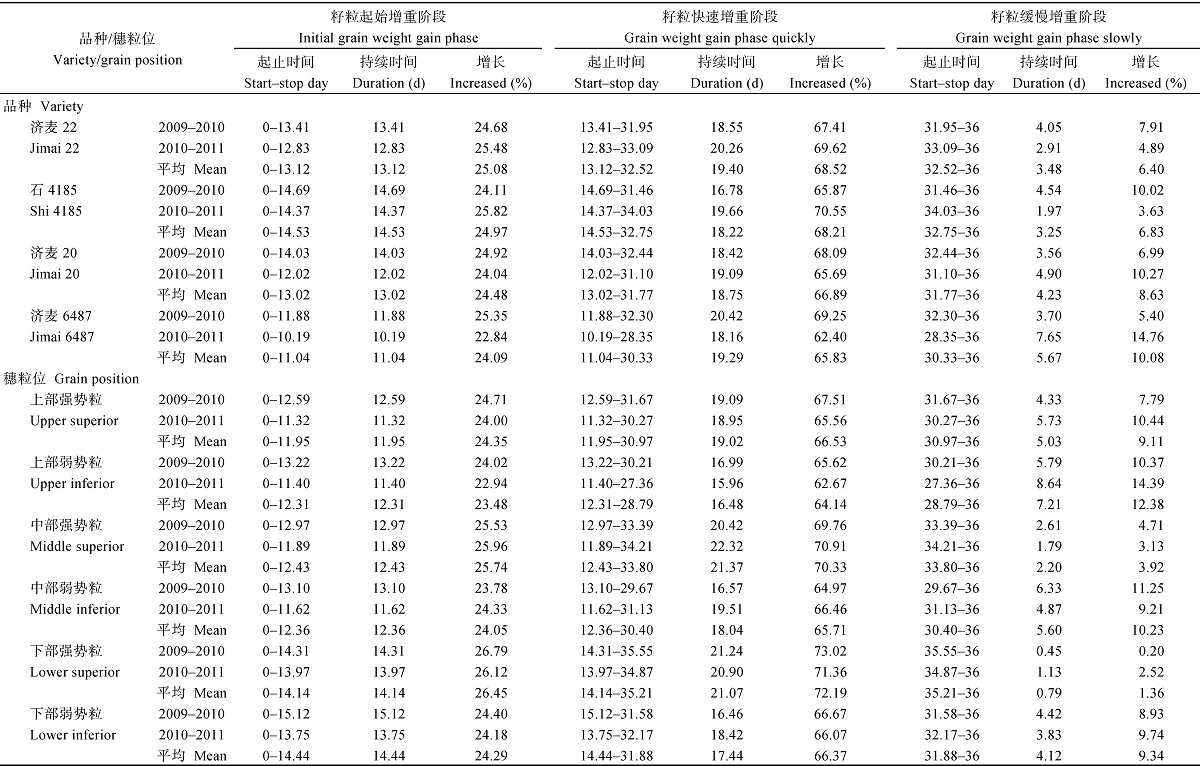
从表6可以看出, 籽粒起始增重阶段, 即建粒阶段, 粒重增加较慢, 起止时间一般在开花后0~12 d, 干物质积累22%~23%; 籽粒快速增重阶段一般在开花后12~26 d, 持续时间14 d左右, 干物质积累61%~62%; 籽粒缓慢增重阶段一般在开花26 d以后, 干物质积累15%~16%。大粒品种济麦6487, 在起始增重阶段和快速增重阶段的持续时间和干物质积累比例明显小于其他品种, 而进入缓慢增重阶段的持续时间和干物质积累比例明显大于其他品种。强势粒在起始增重阶段和缓慢增重阶段的持续时间和干物质积累比例基本小于弱势粒, 而在快速增重阶段大于弱势粒。
| 表6 小麦籽粒不同发音阶段粒重的变化动态 Table 6 Dynamics of grain weight ai different grain-developmental phases |
2.3.2 蛋白质积累动态 籽粒蛋白质积累量呈“ S” 型曲线变化, 也可用Logistic生长方程较好拟合, 相关系数除2010年的济麦6487为0.987外, 均在0.99以上(表7)。可以看出, 蛋白质最大积累速率一般出现在开花后21~24 d, 较粒重晚3 d左右, 积累量一般为0.20~0.23 mg 粒-1 g-1。4个不同基因型品种中, 大粒品种济麦6487 (优质面包)蛋白质最大积累速率出现时间明显早于其他品种, 但蛋白质最大积累速率年度间具有较大变异; 而小粒品种济麦20(优质面包)蛋白质最大积累速率最小, 但年度间变异较小。从不同穗粒位籽粒蛋白质最大积累速率来看, 随着穗位的降低出现时间推迟, 强势粒又晚于弱势粒, 蛋白质最大积累速率表现为中部强势粒> 下部强势粒> 上部强势粒> 下部弱势粒> 中部弱势粒> 上部弱势粒。
| 表7 小麦籽粒蛋白质积累速率的Logistic方程参数 Table 7 Parameters of Logistic equation for fitting grain protein accumulation in wheat |
从表8可以看出, 籽粒蛋白起始积累阶段一般在开花后0~13 d, 蛋白质积累24%~25%; 籽粒蛋白质快速积累阶段一般在开花后13~32 d, 持续时间19 d左右, 蛋白质积累67%~68%; 籽粒蛋白质缓慢积累阶段一般在开花32 d以后, 蛋白质积累7%~ 8%。与中筋品种相比, 优质面包小麦品种在起始积累阶段和快速积累阶段蛋白质积累比例小, 而在缓慢积累阶段持续时间长、积累比例大, 但差异不大。从不同穗粒位籽粒蛋白质积累阶段差异来看, 起始积累阶段强势粒的持续时间小于弱势粒、蛋白质积累比例大于弱势粒, 但差异较小, 进入快速积累阶段强势粒的持续时间和蛋白质积累比例均明显大于弱势粒, 其中穗中、下部的粒位差更为明显, 而在缓慢积累阶段弱势粒又明显大于强势粒。说明强势粒积累蛋白质比较集中, 主要在快速积累阶段, 而弱势粒后期积累蛋白质相对较多, 可能更易受环境因素的影响。
| 表8 小麦籽粒不同发音阶段蛋白质积累量的变化动态 Table 8 Dynamics of grain pertein accumulation at different grain-developemntal phases |
强势粒在灌浆前期, 即开花后12、18和24 d表现负相关, 在灌浆后期, 即开花后30 d和36 d表现正相关(除下部强势粒在开花后36 d为负相关外), 但均未达到显著水平。除上部弱势粒的开花后24 d和下部弱势粒的开花后18 d为负相关外, 其他均为正相关, 其中中部弱势粒的开花后24 d和36 d达到显著水平。说明小麦籽粒C、N竞争主要在灌浆前期, 而灌浆后期粒重和蛋白质含量可以同步提高。
在小麦产量构成三因素中, 粒重的遗传力最大[14], 因此许多育种者将提高粒重作为实现高产优质的主攻方向。远彤等[15]研究发现, 大粒型品种输导组织(尤其是大维管束)以及同化组织发达, 使其籽粒形成期具备了良好的物质转运与积累系统, 但由于籽粒库容大、灌浆期长, 易受极端气候(温度等)的影响, 造成产量和品质不稳定。本研究结果表明, 大粒品种济麦6487, 在生长有利年份, 开花后各时期的粒重和蛋白质积累量显著大于其他品种, 而在不利年份, 低于中粒品种。相对于大粒品种, 小粒品种籽粒库容小, 且光合调控能力强[16], 容易得到充实。小粒品种济麦20, 在开花后各时期的粒重和蛋白质积累量小于其他品种, 但成熟期变异系数小, 产量和品质比较稳定。两个参试中粒品种中, 济麦22粒重、蛋白质积累量和蛋白质含量均稳定; 而石4185蛋白质含量比较稳定, 粒重和蛋白质积累量变异大。通常高温胁迫容易造成小麦的产量及品质下降[17]。分析气象资料发现, 在2011年小麦灌浆后期最高气温达到34.6℃(数据来源于山东省气候中心), 从而反映出济麦22具有一定的耐热特性, 这与之前我们的耐热研究结果一致[18]。小麦穗部各小穗位和粒位的籽粒发育受营养物质供应水平、遗传因素以及外界环境因素影响, 籽粒发育存在不均衡性, 导致不同穗粒位粒重和蛋白质含量存在很大差异[19, 20, 21, 22, 23, 24, 25]。本试验表明, 粒重、蛋白质积累量和蛋白质含量的穗粒位间变异大于年份间变异和基因型间变异, 且花后各时期强势粒显著大于弱势粒, 中部籽粒显著大于上部和下部籽粒, 而随着灌浆进程中部与下部籽粒的差异变小, 成熟期中部和下部籽粒的蛋白质含量无显著差异。这是因为籽粒形成和灌浆过程的初期, 转运到中部小穗的养料最多, 其次是上部小穗, 下部小穗较少; 在灌浆过程的中、后期, 除供应中部小穗较多的养料外, 供应下部小穗较上部小穗为多[3]。综上所述, 小麦品种籽粒不宜过大, 应注重品种的耐热性, 减少弱势粒, 增加中、下部籽粒。
不同品种和穗粒位的蛋白质含量均呈“ V” 型曲线变化, 在开花后24 d达到最低点。这与前人研究结果基本一致[26, 27, 28, 29]。在灌浆后期, 中部强势粒、下部强势粒和下部弱势粒蛋白质含量回升速度明显快于其他穗粒位籽粒, 可见这3个穗粒位在提高籽粒蛋白质含量上具有很大的潜质。不同类型品种粒重和蛋白质积累动态基本一致, 灌浆前中期积累速率快、量多, 积累绝大部分, 奠定基础; 灌浆后期主要合成储藏蛋白[26], 对加工品质非常关键, 但更易受环境因素、气象灾害如干热风等的影响, 面包小麦后期蛋白质积累量相对较高。同样, 强势粒前中期积累速率快、量多, 弱势粒后期积累相对较多, 可能更易受环境影响, 对最后粒重和蛋白质含量/品质稳定性影响较大。杜金哲等[28]研究认为, 籽粒蛋白质绝对累积量与粒重的消长一致。但籽粒灌浆不同阶段对粒重和蛋白质积累的影响不同, 其中最大灌浆速率与粒重正相关[30], 快速积累阶段对提高粒重[31, 32]、改善品质[33, 34]尤为重要。本研究结果表明, 粒重最大生长速率一般出现在开花后18~21 d、快速增重时期为开花后12~26 d, 蛋白质最大积累速率一般出现在开花后21~24 d、快速积累时期为开花后13~32 d, 可以看出, 干物质和蛋白质同时快速积累时期为开花后13~26 d, 因此提高此阶段灌浆速率可能是产量和品质协调提高的有效途径。
本研究选用的济麦22为目前年种植面积最大的高产中筋广适型品种, 石4185为国家黄淮北片多年对照品种, 济麦20曾为年推广面积最大的优质面包小麦品种, 济麦6487为优质面包小麦品系。选用上述4个品种, 拟通过分析小麦穗粒位籽粒形成过程中粒重和蛋白质含量变化动态及蛋白质积累规律差异, 探寻高产和优质面包小麦的穗型契合点, 为高产优质小麦新品种培育提供参考依据。从本试验结果来看, 强势粒产量和品质均明显优于弱势粒、且更加稳定, 下部弱势粒在灌浆盛期灌浆速率大, 且后期蛋白质含量回升速度快, 强势粒粒重与蛋白质含量相关不显著, 而成熟期中部弱势粒与之显著正相关。因此, 在选育高产优质小麦品种过程中, 籽粒不宜过大, 尽量减少小花位数, 适当增加中、下部籽粒[10, 24], 提高开花后13~26 d的灌浆速率, 加强耐热性选择, 这可能是产量和品质进一步协调提高的有效途径。然而, 本试验仅分析了开花后小麦穗粒位粒重和蛋白质含量的变化动态, 实际蛋白质组分与各组分的分布对小麦加工品质也具有重要影响[26], 对此今后工作中还需进一步研究。
小麦籽粒的着生部位对开花后各时期的粒重、蛋白质积累量和蛋白质含量有重大影响, 其变异大于年份和基因型变异。大粒品种较小粒品种更容易受环境影响, 稳定性差。籽粒干物质和蛋白质积累量均表现为强势粒大于弱势粒。强势粒前、中期积累速率快、量多, 受气候影响小, 产量和品质更加稳定。成熟期, 中部强势粒、下部强势粒和下部弱势粒蛋白质含量较高, 对加工品质非常关键。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|