
{kind=link}
{kind=link}
{kind=link}
套作荫蔽对苗期大豆叶片结构和光合荧光特性的影响
[范元芳 , 杨峰
, 杨峰* , 刘沁林, 谌俊旭, 王锐, 罗式伶, 杨文钰* ]
, 杨峰, 刘沁林, 谌俊旭, 王锐, 罗式伶, 杨文钰]
|
|


第一作者联系方式: E-mail: 851301600@qq.com
间套作系统中荫蔽是影响低位作物生长发育的主要因素。本研究采用玉米-大豆套作种植模式, 以南豆12大豆品种为研究对象, 设置2个行比配置, 即1:1 (A1处理; 1行玉米间隔1行大豆, 间距50 cm)和2:2 (A2处理; 2行玉米间隔2行大豆, 玉米和大豆各自的行距均为40 cm, 玉米和大豆间距为60 cm), 净作大豆为对照(CK), 研究玉米和大豆不同行比套作配置下荫蔽对大豆叶片光合荧光特性、解剖结构及超微结构的影响。结果表明, 套作处理A1、A2的光合有效辐射(PAR)分别比CK显著降低91.2%、66.8%。叶绿素 a含量分别降低50.2%和27.9%, 净光合速率分别降低63.2%和37.8%。套作大豆功能叶片荧光参数 Fv'/ Fm'和 Fq'/ Fm显著高于净作大豆, 而 Fv/ Fm、 qP和NPQ低于净作。对于叶片结构, 套作大豆叶片厚度、栅栏组织厚度、海绵组织厚度均低于CK, 且净作大豆叶片厚度分别是A1和A2处理下的2.15倍和1.69倍; 套作大豆叶片的叶绿体结构比较完整, 无破碎现象, 含淀粉粒稀少, 随着荫蔽程度的加重, 嗜锇颗粒、基粒片层和基质片层增多, 叶绿体减小但其数目增多, 而净作大豆叶片叶绿体中淀粉粒较多且色泽亮白, 基粒片层和基质片层稀少。因此, 玉米-大豆套作种植中不同行比配置导致低位作物大豆冠层光环境差异, 直接影响大豆叶片结构特征和光合荧光特性。
Shading in relay intercropping system is a major factor affecting the low growing of crop growth and development. This study attempted to compare leaf structure and photosynthetic characteristics of cultivar Nandou 12 under maize-soybean relay intercropping systems. Three treatments were designed including one row maize to one row soybean with 50 cm of row space between maize and soybean (A1); two rows maize to two rows soybean with 40 cm of row space for maize or soybean and 60 cm of row space between maize and soybean (A2), sole cropping of soybean. The result indicated that shading caused the changes of both anatomical structure of leaf and photosynthetic characteristics in maize/soybean relay strip intercropping systems. During the V5 stage of soybean, the PAR density in treatments (A1 and A2) were lower than that in CK by 91.2% and 66.8%, respectively. The content of Chl a of treatments (A1 and A2) was lower than that in CK by 50.2% and 27.9%. The Pn of treatments (A1 and A2) was lower than that in CK by 63.2% and 37.8%. The Fv/ Fm, qP, NPQ in leaf of treatments (A1 and A2) were lower than there in CK significantly, Fv'/ Fm' and Fq'/ Fm' of treatments (A1 and A2) were higher than that in CK. Under light microscope, compared with CK, A1 and A2 treatments decreased palisade tissue thickness and spongy tissue thickness of leaf. The leaf of treatment CK was 2.15 and 1.69 times as thick as that of treatments A1 and A2. In A1 and A2 treatments leaf chloroplast morphology was normal, the number of chloroplasts, granule lamella and osmophilic globule in chloroplast increased; the structure of chloroplast was not damaged; There were few starch grains in chloroplasts; and the structure of cell organelles was normal. Therefore, the different spatial patterns of maize-soybean intercropping system result in different light environments above the soybean canopy, and directly affect soybean leaf structure and photosynthetic fluorescence characteristics.
光合作用是植物生长发育和产量的基础, 叶片是植物光合作用的主要器官[1], 植物体的形态、结构与生理功能是统一的, 光合作用的差异与植株叶片光合器官的解剖学特征紧密相关[2]。覃凤飞等[3]研究表明紫花苜蓿的叶片厚度、栅栏组织厚度和海绵组织厚度分别与叶片的气体交换、蒸腾作用、光能捕获结构和功能相关。甄伟等[4]、马慧丽等[5]和黄俊等[6]发现弱光影响光合机构和活性, 但不破坏光合机构。植物的叶片解剖结构反映植物对环境变化的响应, 是植物进化过程中对环境变化较敏感且可塑性较大的器官, 叶片结构可塑性已成为衡量植物对光胁迫环境适应能力的重要指标[7]。一些研究表明弱光使苹果[5]、桃叶[8]、三叶爬山虎[9]、番茄[10]、高粱[11]等叶片的栅栏组织、海绵组织厚度和叶片总厚度降低, 但是叶肉细胞内的叶绿体数增多, 叶绿体基粒明显变厚, 基粒数和基粒片层数增加, 淀粉粒数降低。大豆[Glycine max(L.) Merr.]是世界上重要的豆类作物之一, 同时也是我国主要的粮食和经济作物[12]。因此, 明确玉米-大豆套作下不同行比配置的大豆功能叶片解剖结构、超微结构及光合荧光特性将为大豆高产育种栽培研究提供理论依据。
试验地的土壤为紫色黏土, pH为6.8。玉米品种为川单418, 由四川农业大学玉米所提供。大豆品种为南豆12, 由四川省南充市农业科学研究所提供。
2014年和2015年的4月至11月在四川省仁寿县珠嘉乡的四川现代粮食生产示范基地采用单因素完全随机区组设计, 小区面积7 m × 6 m, 3次重复。设置2个处理(图1), 处理A1 (1行玉米, 1行大豆, 间距50 cm)和A2 (2行玉米, 2行大豆, 玉米、大豆各自的行距均为40 cm, 玉-豆间距60 cm); 以净作大豆(CK)为对照, 行距70 cm。玉米株距16.7 cm, 大豆株距10 cm, 净作大豆与套作种植密度一致。2014年, 玉米3月27日育苗, 4月10日移栽定植, 8月10日收获; 大豆6月17日播种, 10月21日收获。2015年, 玉米3月29日育苗, 4月13日移栽定植, 8月13日收获; 大豆6月19日播种, 10月22日收获。各小区施肥水平一致, 其他按照大田常规管理进行。
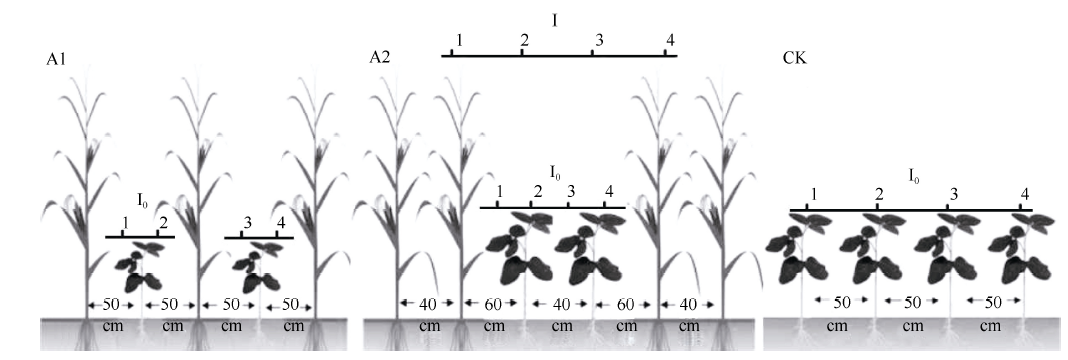 | 图1 PAR传感器在玉米-大豆带状套作组合中的摆放位置示意图Fig. 1 Diagram of maize-soybean relay strip intercropping patterns and layout of photo synthetically active radiation sensors |
1.3.1 大豆冠层光合有效辐射 参照Fehr等[13] 划分的生育时期测定, 大豆取样时间为V5期(五节期)。在玉米和大豆共生阶段, 于大豆V5期在晴天的11:00— 13:00, 利用美国LI-COR公司的LI-1400光量子仪, 在各处理的小区中间区域, 移动探测杆, 记录瞬时PAR量子通量读数[14]。沿大豆冠层顶部5 cm高度由西向东在测点1、2、3、4记录一次PPFD读数(图1), 计算平均值, 玉米冠层作为I, 在各处理的测点1记录一次PPFD读数, 作为I0。并采用如下公式进行透光率计算[15, 16]。透光率= I0/ I, 式中I0为各处理大豆冠层顶部的辐射强度, I为玉米冠层顶部的辐射强度[17]。
1.3.2 光合色素含量 在大豆V5期每个小区中间位置连续取长势一致大豆植株5株的功能叶片(倒三复叶的中间小叶), 用直径为1.2 cm的打孔器将每张叶片打4个孔, 放入80%丙酮溶液15 mL中, 在20℃下暗处浸提24 h, 过滤后在波长663、645和470 nm处测定光密度, 然后计算叶绿素a、叶绿素b和类胡萝卜素含量。每个小区均以3次测定的平均值作为其色素含量[18]。
1.3.3 叶片解剖结构 在大豆V5期, 选择各小区中间位置具有代表性大豆植株5株的功能叶片(倒三复叶的中间小叶), 在靠近主叶脉基部的1/3处取样(5 mm × 5 mm左右), 用标准固定液(FAA)固定保存, 用酒精和正丁醇系列脱水, 石蜡包埋, 采用德国产的Leica切片机切片, 横切片厚度为10 μ m, 用松节油和酒精系列脱蜡、复水后进行番红染色, 中性树胶封片, 采用Nikon eclipse50i型显微镜观察切片组织和细胞排列情况, 并用ACT-2U成像系统显微照相及分析[19]。
1.3.4 叶绿体超微结构 在大豆V5期, 选择各小区中间位置具有代表性大豆植株5株的功能叶片(倒3复叶的中间小叶), 在靠近主叶脉基部的1/3处取样(5 mm× 5 mm左右), 经3%戊二醛预固定, 1%四氧化锇再固定, 丙酮逐级脱水, Epon812包埋, 半薄切片光学定位, 超薄切片, 醋酸铀及枸橼酸铅双重染色, 日立H-600IV型透射电镜观察拍照。每个处理观测15个视野。
1.3.5 光合特性 在大豆V5期分别选取各小区中间位置长势均匀的大豆植株5株, 在晴天的9:00— 11:00用美国的LI-COR公司生产的LI-6400便携式光合仪在田间测定功能叶片(倒三叶中间小叶)的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr), 3次重复[20, 21]。
1.3.6 叶绿素荧光特性 采用英国Technologica公司的叶绿素荧光成像系统(CFI)测定Fv/Fm(PSII最大光化学量子产量)、Fq'/Fm' (PSII实际光化学效率)和qP (光化学荧光猝灭系数)。测定过程中, 让大豆叶片充分暗适应15 min后获得暗处理数据, 随后选择光强为800 mol m-2 s-1, 让叶片充分光适应5 min后获得光处理数据。每个小区参数均为3次测定的平均值[22, 23]。
采用Microsoft Excel 2013软件整理数据, SPSS17.0软件统计分析。
由表1可知, 在玉米-大豆共生期(V5期), 不同套作荫蔽条件下, 大豆群体PAR和透光率存在显著差异, 且均显著低于CK (P< 0.05)。处理A1、A2的PAR分别比CK显著降低91.2%、66.8%。处理A1、A2的透光率分别为CK的8.7%、31.1% (2014年和2015年平均值)。
| 表1 玉米-大豆套作不同行比配置下大豆冠层光分布 Table 1 Light distribution of soybean canopy in different maize-soybean relay intercropping systems |
由表2可知, 与CK相比, 处理A1、A2的单位面积光合色素含量均显著降低。处理A1、A2大豆功能叶片的叶绿素a含量分别比CK降低了50.2%和27.9%, 叶绿素b含量分别降低了46.4%和13.7%, 类胡萝卜素含量分别降低了49.9%和33.6%; 叶绿素a含量与叶绿素b含量的比值分别比CK降低了6.9%和16.3%。套作荫蔽越严重的处理A1光合色素下降程度越大, 但是A1的叶绿素a含量与叶绿素b含量的比值下降的程度小于A2 (2014和2015两年平均值)。
| 表2 玉米-大豆套作不同行比配置下大豆叶片光合色素含量 Table 2 Photosynthetic pigments of soybean leaf in different maize-soybean relay intercropping systems (mg dm-2) |
由表3可知, 玉米-大豆共生期, 套作荫蔽条件下大豆叶片的净光合速率(Pn)均显著低于CK (P< 0.05), 且套作荫蔽程度越重, 大豆功能叶片的净光合速率降低幅度越大, 表现为A1< A2< CK。与CK相比, 处理A1、A2的净光合速率分别显著降低63.2%和37.8%; 胞间CO2浓度显著增加7.3%和5.4%; Gs和Tr与Pn变化趋势相同(2014和2015两年均值)。
| 表3 玉米-大豆套作不同行比配置下大豆光合特性 Table 3 Photosynthetic characteristics of soybean in different maize-soybean relay intercropping systems |
由表4可见, 与净作相比, 套作荫蔽条件下大豆功能叶片的PSII有效光化学量子产量(Fv'/Fm')和实际光化学效率(Fq'/Fm')均呈增加趋势。与CK相比, 处理A1显著降低了14.5%, A2与CK差异不显著。套作荫蔽条件下大豆叶片的PSII最大光化学量子产量(Fv/Fm)、光化学荧光猝灭系数(qP)、非光化学荧光猝灭系数(NPQ)均减小, 其中处理A1的非光化学荧光猝灭系数与CK达到显著差异水平, 均显著低于CK 18.7%, A2与A1和CK均无显著差异。
| 表4 玉米-大豆套作不同行比配置下大豆叶绿素荧光参数 Table 4 Fluorescence parameters of soybean in different maize-soybean relay intercropping systems |
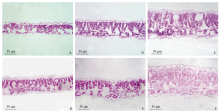
大豆在处理A1、A2和CK处理下的叶片解剖结构(图2)和观测结果(表5)表明, 大豆叶片类型属于异面叶, 叶肉组织分化为明显的栅栏组织和海绵组织, 但在3种配置下存在明显的差异。如图2所示, 各处理的大豆叶片厚度、栅栏组织厚度、海绵组织厚度和表皮细胞厚度均低于CK, 2种套作配置对其影响依序为A1< A2< CK。CK的功能叶片的上、下表皮细胞、栅栏组织叶肉细胞和海绵组织细胞略大, 且排列致密; 处理A1、A2的叶片栅栏组织叶肉细胞较小, 排列疏松, 细胞间隙较大, 下表皮细胞变薄。进一步量化可见(表5), 净作下大豆功能叶的叶片厚度分别是处理A1、A2的2.15倍、1.69倍; A2的上表皮细胞厚度和栅海比与CK均未达到差异显著水平; A1与A2的海绵组织厚度差异不显著; A1、A2的栅栏组织厚度分别显著比CK降低了68.7%、47.9%, 海绵组织厚度分别降低了46.6%、24.1%。不同玉米-大豆空间配置下荫蔽对叶片解剖结构的影响存在一定的差异。
 | 图2 玉米-大豆套作不同行比配置下大豆叶片解剖结构 A~C: A1、A2处理和CK (2014年); D~F: A1, A2处理和CK (2015年) (× 400)。Fig. 2 Anatomical structure of soybean leaf in different maize-soybean relay intercropping systems A-C: A1, A2 treatments and CK (2014); D-F: A1, A2 treatments and CK (2015) (× 400). |
| 表5 玉米-大豆套作不同行比配置下大豆叶片解剖结构 Table 5 Anatomical structure parameter of soybean leaf in different maize-soybean relay intercropping systems |
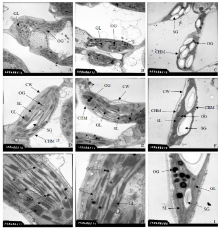
图3所示, 与对照相比, 套作荫蔽处理条件下的大豆叶片的叶绿体结构完整, 没有发生破碎现象, 不同程度荫蔽环境下的大豆叶片的基粒片层和基质片层受到影响程度不同, 处理A1、A2的叶绿体的超微结构发生明显变化, 部分叶绿体形状呈不规则的长椭圆形, 中间部分背向细胞壁凸起。各荫蔽处理均降低了单个细胞和叶绿体的大小, 但是叶绿体的数目却明显增多, 处理A1的叶绿体数目最多但是个体较小(图3-A)。套作荫蔽条件下大豆叶片叶绿体中的淀粉粒稀少, 嗜锇颗粒增多; CK叶绿体中的淀粉粒较多且色泽亮白, 基粒片层稀少(图3-I); 套作处理A1、A2的大豆叶片的叶绿体中的基粒片层和基质片层分布密集。基粒片层数目的增加增强了叶绿体的光能吸收、传递和转换功能, 利于光合产物的形成和积累。
 | 图3 玉米-大豆套作不同行比配置下大豆叶片超微结构 A~C: 叶绿体分布(× 6000); D~F: 叶绿体超微结构(× 12 000); G~J: 叶绿体基粒片层(× 25 000); CW:细胞壁; CHM: 叶绿体膜; GL: 基粒片层; SL: 基质片层; SG: 淀粉粒; OG: 嗜锇颗粒; A, D, G: A1处理; B, E, H: A2处理; C, F, I: CK。Fig. 3 Chloroplast microstructure of soybean leaf in different maize-soybean relay intercropping systems A-C: chloroplast distribution (× 6000); D-F: chloroplast ultrastructure (× 12 000); G-I: grana layer in chloroplast (× 25 000); CW: cell wall; CHM: chloroplast membrane; GL: grana lamellae; SL: stroma lamellae; SG: starch grain; OG: csmiophilic granule; A, D, G: A1 treatment; B, E, H: A2 treatment; C, F, I: CK. |
光合有效辐射(PAR)是植物生命活动、有机物质合成和产量的能量来源, PAR透光率是冠层内指定高度的PAR与冠层顶部PAR的比值[24]。光合有效辐射直接影响植物的生长发育、产量和品质, 在玉米-大豆套作不同配置下, 套作处理A1、A2的PAR分别为CK的10%、30%左右。套作群体中高位作物玉米的冠层分布直接导致了大豆群体的光能截获量的差异, 形成了不同程度的荫蔽, 从而影响大豆植株的生长。崔亮等[14]研究表明不同玉米-大豆带状套作组合形成不同的大豆条带生长光环境, 该系统下大豆生长带内的PAR和透光率均显著下降, 本研究结果与崔亮等研究结果基本一致。
叶绿素含量对叶绿体的光合活性具有重要意义, Chl b/a比值增大时, 叶绿体对2, 6-二氯-靛酚的还原能力增强, 叶绿体的光合磷酸化活性提高, 利于提高光合速率[3, 25]。本研究中发现, 套作荫蔽显著降低了大豆功能叶片的单位面积叶绿素含量, 但是处理A1的Chl b/a的比值下降的程度小于A2。说明大豆功能叶片在严重的荫蔽环境中通过调节自身叶绿素a与叶绿素b的比值来提高光合作用, 这与黄俊等[6]、姚允聪等[8]研究结果类似。叶绿素荧光作为光合作用研究的灵敏探针, 能够很好地反映逆境因子对光合作用的影响[26, 27]。在本试验中, 套作荫蔽增加了大豆功能叶片的Fv'/Fm'和Fq'/Fm', 降低了Fv/Fm、qP和NPQ, 说明套作荫蔽导致大豆幼苗叶片PSII反应中心光合电子传递速率下降, 使累积在PSII反应中心的光能过剩, 为了维持较高的能量代谢水平, 大豆叶片通过降低热耗散来提高的光化学反应效率。本研究测定光合参数发现套作荫蔽显著降低了大豆叶片的Pn、Gs和Tr, 且套作荫蔽程度越严重, 其降低幅度越大。这与崔亮等[14]和苏本营等[20]研究结果相似。
植物体的形态、结构和生理功能是统一的, 光合速率的差异也必然与叶片光合作用器官的解剖学特性紧密相关[28], 叶片解剖结构能够反映植物对环境变化的响应, 厚叶结构、发达的栅栏组织、排列紧密平直的表皮细胞是阳生叶的典型特征, 而发达的海绵组织、凸透形的表皮细胞则是叶片对于弱光、水分充足的一种适应特性[9, 10]。刘凡值等[29]研究表明, 间作荫蔽使大豆叶片的表皮细胞增大且排列疏松, 栅栏组织厚度由于细胞层数的减少而低于净作, 海绵组织间隙大、呈椭圆形。本研究中, 大豆叶片厚度随套作荫蔽增强而变薄, 造成该差异的主要原因是套作大豆叶片栅栏组织厚度的减小, 但是栅栏组织细胞层数并未发生变化, 均为两层, 随着荫蔽程度的加重, 栅栏细胞排列呈现出由紧密向疏松变化的趋势, 形状由长棒状趋于短锥状; 表皮细胞逐渐减小(图1), 海绵组织细胞排列变得稀疏。叶绿体是光合作用的场所, 叶绿体数目多则叶绿素含量和光合速率高[30, 31]。本试验中, 套作荫蔽降低了单个细胞和叶绿体的大小以及单个叶绿体中的淀粉粒数, 嗜锇颗粒、叶绿体数、基粒数, 基粒片层数目普遍高于对照, 基粒片层数目的增加增强了叶绿体的光能吸收、传递和转换功能, 利于光合产物的形成和积累, 这与冯乃杰等[2]、姚允聪等[8]的研究结果相一致。一些相关研究指出, 叶绿体根据光线的强弱发生形态调整, 甚至发生结构变化, 以增强对弱光的吸收利用能力[26, 27]。套作荫蔽条件下, 大豆叶绿体的基粒数和基粒片层数增加, 但其光合速率显著降低, 这与前人在黄瓜、人参、姜等作物上的研究结果相似[28, 30, 31]。基粒片层的垛叠程度越高, 说明捕获光能机构高度密集, 同时作为酶的排列支架, 有利于形成一个长的代谢传送带, 促进光能的吸收传递和转换[32], 这可以在一定程度上解释套作荫蔽下光化学效率增加的现象。甄伟等[4]研究表明, 弱光处理7 d后, 黄瓜叶片的光合产物运输受阻, 叶绿体内淀粉粒积累。艾希珍等[28]认为淀粉粒的多少可能与弱光处理时间长短有关, 长时间弱光使黄瓜植株对弱光逐渐适应, 光合产物的运输逐渐协调, 淀粉粒减少。弱光下叶绿体内的淀粉粒数的多少, 前人因试验材料、光照时间和光照强度的不同而不同, 本研究结果与艾希珍等研究结果一致。
玉米-大豆套作系统中, 荫蔽降低了大豆叶片单位面积的光合色素含量、Fv/Fm、qP和NPQ, 增加了大豆功能叶片的Fv'/Fm'和Fq'/Fm'。大豆叶片厚度随套作荫蔽增强而变薄, 细胞排列由紧密向疏松变化, 细胞间隙增大。套作荫蔽同时也降低了叶绿体的大小、淀粉粒数; 增加了叶绿体、嗜锇颗粒、基粒和基质片层的数目, 这利于光合产物的形成和积累。大豆在适当的荫蔽环境下表现出良好适应性。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|