


{kind=link}
{kind=link}
{kind=link}
{kind=link}
转 CP4-EPSPs基因大豆杂交后代对草甘膦的抗性水平与遗传背景的相关性
[孙如建1, 2, 3 , 孙宾成1 , 张琪1 , 胡兴国1 , 郭荣起1 , 郭兵福2 , 马岩松2, 4 , 于平1 , 张晓莉1 , 柴燊1 , 张万海1, *  , 邱丽娟
, 邱丽娟2, * ]
, 邱丽娟]
|
|


第一作者联系方式: E-mail: sunrujian321@126.com
第一代抗草甘膦转 CP4-EPSPs基因大豆GTS40-3-2是国际上应用时间最长、种植面积最大的转基因作物。本文以6份GTS40-3-2衍生的抗草甘膦转基因大豆新品系为亲本, 配制4个杂交组合, 利用抗性分级法和相对株高法鉴定杂交亲本及其F2:3子代对草甘膦的耐受性差异, 分析其抗性水平与遗传背景的相关性。结果表明, 以1230 g a.i. hm-2草甘膦喷施处理时, 转基因亲本及其F2:3子代的苗期生长受草甘膦抑制不显著, 而当喷施浓度提高至3690 g a.i. hm-2和4920 g a.i. hm-2时则抑制作用显著。供试的6个杂交亲本中以ZLHJ06-1568、ZLHJ10-713和ZLHJ06-698对草甘膦的耐受性相对较强, 而4个F2:3组合中以ZLHJ10-713 × ZLHJ06-698后代在草甘膦喷施后株高受抑制最小, 对草甘膦耐受性最强。不同组合后代对草甘膦的耐受性普遍优于其双亲, 呈现出杂种优势。各组合后代与亲本之间对草甘膦的耐受性均呈正相关, 但由于亲本间互作效应的不同, 导致后代抗性水平产生差异。本研究表明草甘膦抗性基因 CP4-EPSPs在大豆中的表达水平与其遗传背景相关联, 为利用转基因大豆新种质培育转基因大豆新品种过程中目标基因的定向选择提供了参考依据。
, QIU Li-Juan
The first Roundup Ready (RR) soybean (GTS40-3-2) carrying CP4-EPSPsis one of the transgenic crops that have the longest application time and largest planting acreage worldwide. However, it has not been commercialized and has less report in China. In this study, six RR soybean lines derived from GTS40-3-2 were hybridized to create four combinations. We used resistance grading and relative plant height in identifying the resistance to glyphosate of parents and F2:3 progeny, to investigate the correlation between resistance of the progeny and genetic background. The growth inhibition of transgenic parents and progeny at seeding stage was not significant when the glyphosate treatment concentration was 1230 g a.i. hm-2, while it was significant when the glyphosate concentration was 3690 g a.i. hm-2 and 4920 g a.i. hm-2. ZLHJ06-1568, ZLHJ10-713, and ZLHJ06-698 among the six parents had the more glyphosate resistance relatively. The progeny of ZLHJ10-713 × ZLHJ06-698 combination had the least inhibition of plant height showing highest resistance to glyphosate in all four F2:3 combinations. The progeny had better resistance to glyphosate than their parents showing better heterosis. The glyphosate resistance of progeny was positively correlated with that of parents. Difference of glyphosate resistance was found in progeny among different combinations because of the interaction between parents. Our study indicates that the expression of glyphosate resistance gene is closely related to the genetic background in soybean, providing a theoretical evidence for target gene selection in developing new varieties by using transgenic germplasm.
大豆[Glycine max (L.) Merr.]是重要的粮油作物, 在全世界广为种植, 仅2014年全球大豆种植面积就高达1.185亿公顷[1]。随着人们生活水平的日渐提高, 我国大豆消费需求逐年攀升, 但大豆种植面积和年总产量却在逐年缩减, 其中种植成本高、经济效益低是造成我国大豆生产不景气, 供不应求的主要原因。为满足市场需要只能从国外进口大豆, 2015年进口总量高达8169万吨, 其中主要为抗草甘膦转基因大豆。抗草甘膦转基因大豆是世界上商品化最早、应用最广的转基因作物[2], 自1996年获得商业化生产许可以来, 其种植面积逐年扩大[3, 4, 5], 而我国虽已开展转基因大豆生物技术研究, 但仍未批准转基因大豆的商业化种植, 因此加速我国转基因大豆的研究进程具有重要的实践意义。
草甘膦是一种有机磷类除草剂, 具有低毒低残留、广谱灭生、内吸传导等特点[6, 7]。大豆对草甘膦的耐受性鉴定方法主要有田间喷洒法[8]、种子浸泡法、生物测定法[9]、和定量点施法[11]等, 其中又以田间喷洒法最为常用, 鉴定结果最为准确可靠[8]。草甘膦处理后的作用效果虽因植物种类、喷施剂量等的不同而存在差异, 但在植株茎顶端及叶片上出现的“ yellow flashing” 现象是最显著的标志性药害症状, 该现象在常规大豆中不可恢复, 但在抗草甘膦转基因大豆中是暂时的, 施药后期可恢复[12, 13, 14], 而高浓度的草甘膦则会抑制大豆早期(V1和V2)的株高生长[15]。
我国抗草甘膦转基因大豆仍处于研发阶段, 尚未产业化, 关于转EPSPs基因大豆杂交后代对草甘膦抗性的相关研究较少。刘文娟等[16]利用抗草甘膦转基因大豆GTS40-3-2, 在V2期茎叶喷施不同浓度的草甘膦, 收获的种子于第2年种植, 茎叶喷施相同浓度草甘膦后检测抗性, 结果表明成熟籽粒中外源基因的相对含量没有显著变化, 可见抗草甘膦特性可以在子代中稳定遗传。刘敏等[17]用抗除草剂转基因大豆与常规大豆杂交, 对杂种后代F1、F2、F3和BC1植株在2~3叶期喷施草甘膦, 遗传分析证实抗除草剂基因受一对显性核基因控制, 为杂交育种提供了理论依据。然而, 利用草甘膦抗性基因进行抗草甘膦转基因大豆新品种培育时, 其杂交后代对草甘膦的耐受性是否存在差异尚未见报道。
本试验利用抗草甘膦转基因大豆新品系进行有性杂交, 对亲本、F2和F2:3喷施不同剂量草甘膦, 调查植株叶片及株高的变化情况, 系统分析后代对草甘膦的抗性水平及其与亲本的相关性, 旨在明确转基因供体亲本和衍生后代间的抗性遗传特点, 为抗草甘膦大豆育种提供参考。
1.1.1 供试亲本及后代材料 抗草甘膦大豆品系中联HJ10-625 (ZLHJ10-625)、中联HJ11-140 (ZLHJ11-140)、中联HJ11-142 (ZLHJ11-142)、中联HJ10-713 (ZLHJ10-713)、中联HJ06-698 (ZLHJ06- 698)和中联HJ06-1568 (ZLHJ06-1568)等6个(表1), 均衍生于商业化推广的抗草甘膦转CP4-EPSPs基因大豆GTS40-3-2, CP4-EPSPs表达框在GTS40-3-2中为单拷贝插入(http://gmdd.shgmo.org/event/view/ 41)。2012年配制ZLHJ11-140 × ZLHJ10-625、ZLHJ11-142 × ZLHJ10-625、ZLHJ10-713 × ZLHJ06- 698和ZLHJ10-713 × ZLHJ06-1568等4个杂交组合, 2013年种植F1, 2014年种植F2, 单株收获后从每个杂交组合随机选择10个单株(10次重复)种植F2:3株行, 每个株行20株, 同时种植各组合亲本。
| 表1 杂交亲本相关信息 Table 1 Information of crossing parental |
1.1.2 喷雾器和除草剂 选用3WBD-18型背负式电动喷雾器, 由台州市吉达喷雾厂生产; 喷施所用Roundup的草甘膦异丙胺盐有效剂量为41%, 由美国孟山都公司生产。
在植株生长至第一片三出复叶完全展开(V2期)时, 对亲本和F2:3株系分别喷施清水(T1)和1230 (T2)、3690 (T3)、4920 g a.i. hm-2(T4)的草甘膦, 即0倍、1倍、3倍和4倍的生产推荐浓度(0、3000、9000和12 000 mL hm-2), 于喷药7 d后调查株高变化及叶片药害等级。
1.3.1 相对株高法[18] 调查每个单株喷药前及喷药7 d后的株高, 得到株高平均变化值Δ h1、Δ h2、Δ h3和Δ h4, 并计算T2、T3、T4与T1 (CK)的比值。
以T2为例, 相对株高变化(Hx)=Δ h2 /Δ h1× 100%, 该值越大表明株高受抑制越轻, 抗性越强。
1.3.2 抗性分级法[19]X(%) = [∑ (N× S) / (T× M)] × 100, 式中X为药害指数, N为同级受害株数, S为药害级别数, T为总株数, M为最高级别。
即药害指数(%) = (1× 1级药害株数+2× 2级药害株数+3× 3级药害株数+4× 4级药害株数)/(总株数× 最高药害级)× 100, 其中最高药害级为4。
1.3.3 药害等级划分标准[20] 0级为植株生长正常, 无任何受害症状, 与对照(T1)相比, 无明显差异; 1级为0~25%叶片产生暂时性的接触性药害斑、皱缩或生长受轻微抑制; 2级为25%~50%叶片产生较重的连片药害斑, 褪绿、皱缩、畸形或者生长明显受抑制, 但可以恢复; 3级为50%~75%叶片产生药害, 或造成生长点死亡, 生长严重受抑制; 4级为75%以上的叶片产生药害, 或部分或全部植株死亡。
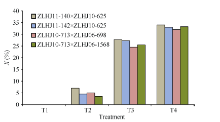
利用4种浓度草甘膦对6份亲本材料喷施。用抗性分级法测定受害程度表明, 各亲本在T2 (1230 g a.i. hm-2)下平均药害指数为6.96%, 药害较轻, 叶片无明显变化, 而在T3 (3690 g a.i. hm-2)和T4 (4920 g a.i. hm-2)下叶片受害相对较重, 平均药害指数分别为29.63%和39.58%, 与T2差异显著(图1)。
 | 图1 6个亲本对草甘膦的药害指数Fig. 1 Glyphosate phytotoxicity index of six parents |
方差分析(表2)及多重比较(表3)表明, ZLHJ06- 1568、ZLHJ10-713和ZLHJ06-698的平均药害指数分别为24.41%、24.58%和24.64%, 显著低于ZLHJ11-140 (27.83%)和ZLHJ11-142 (27.33%), 对草甘膦表现出较强的耐受性, 但总体而言, 各亲本均表现出较强的耐受性, 植株在高浓度处理后虽有药害症状表现, 但经过一段时间均能恢复正常生长。
| 表2 亲本对草甘膦抗性的方差分析 Table 2 Variance analysis of parents' resistance to glyphosate |
| 表3 亲本对草甘膦抗性的多重比较 Table 3 Multiple comparison of parents' resistance to glyphosate |
对4个组合F2群体喷施生产推荐剂量1230 g a.i. hm-2的草甘膦, 植株生长正常且未发现明显的药害症状, 表明各亲本携带的草甘膦抗性基因均已通过杂交的方法转移至F2群体中。
2.3.1 抗性分级法 在不同浓度草甘膦处理下, 不同组合F2:3株系的叶片受害程度表现出相同的变化趋势(图2)。
 | 图2 4个杂交组合F2:3叶片不同处理下药害表型Fig. 2 Injured phenotype of F2:3 leaves of four combinations at each concentration |
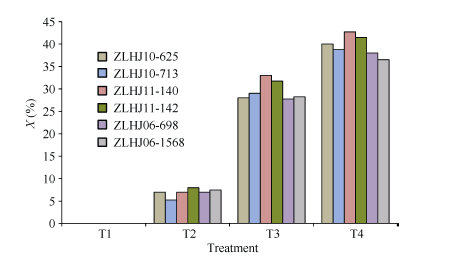
利用叶片受害表型鉴定转基因大豆F2:3对草甘膦处理的抗性反应, 结果表明在相同浓度草甘膦处理下, 各组合间的抗性差异不显著, 其中ZLHJ10- 713 × ZLHJ06-698在T3和T4下的药害指数分别为24.5%和32.0%, 对草甘膦的抗性较强, ZLHJ11-140 × ZLHJ10-625抗性较差, 药害指数分别为27.8%和34.0%。不同浓度草甘膦处理间相比, 低浓度的T2与T1相比药害症状不明显, 平均药害指数仅为5.0%, 而较高浓度的T3 (26.3%)和T4 (33.1%)下药害症状较重, 与T1、T2相比差异显著(图3)。
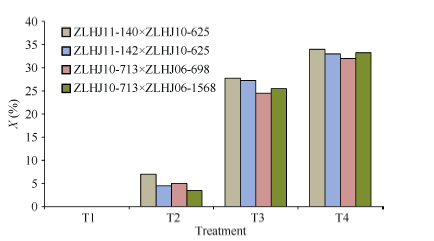 | 图3 4种不同草甘膦处理后4个杂交组合F2:3的药害指数比较Fig. 3 Comparison of Phytotoxicity rating in F2:3 generation of four crosses treated with four glyphosate concentrations |
2.3.2 相对株高法 同一浓度下F2:3群体不同组合间对草甘膦的耐药性存在差异(表4)。T2水平下
各组合间的差异不显著; 在T3水平下, ZLHJ10-713 × ZLHJ06-698的相对株高变化最大, 达到83.28%, 抗性最强, 与ZLHJ10-713 × ZLHJ06-1568和ZLHJ11-140 × ZLHJ10-625差异达到极显著水平; 在T4水平下, ZLHJ10-713 × ZLHJ06-698的抗性同样最强(76.45%), 与ZLHJ10-713 × ZLHJ06-1568及ZLHJ11-140 × ZLHJ10-625差异达到显著水平。
4个组合不同处理间株高变化呈现一致的变化规律, 其中T1的相对株高值最大, 与T2差异不显著, 但与T3、T4间差异均达到极显著水平; T2与T3、T4间除ZLHJ10-713 × ZLHJ06-698外差异均达到极显著水平, 可见生产推荐浓度的草甘膦对转基因大豆后代的苗期株高影响极小; 而T3和T4浓度下株高受抑制显著, 且两处理间差异不显著。
| 表4 不同处理、不同组合的株高相对变化 Table 4 Relative variation of plant height for each treatment combination |
利用SPSS分析相对株高法和抗性分级法两种抗性测定结果, 二者相关系数为0.821, 达到极显著相关。次年在大田种植并喷药, 用同样方法重复鉴定F2:3抗性, 与之前结果相关系数为0.409, 呈显著相关, 表明杂交转育后代对于草甘膦的抗性稳定性较高, 试验重复性较好。说明两种方法均可用于对大豆抗草甘膦特性的评价, 可以根据实际情况选用适宜的鉴定方法。
各组合亲本与F2:3在不同浓度下表现出的抗性水平规律相似, F2:3抗性普遍优于双亲, 这是由于F2:3具有一定的杂种优势。亲本抗性较高的材料, 其后代大部分抗性也较高, 但由于亲本之间存在互作效应, 后代的抗性并不是双亲的简单累加, 会出现双亲高抗但杂交后代抗性相对较低的情况, 例如T4下的ZLHJ10-713 × ZLHJ06-698和ZLHJ10-713 × ZLHJ06-1568等(图4)。
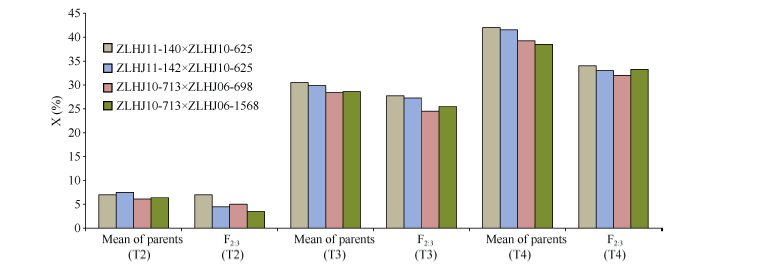 | 图4 不同草甘膦处理条件下亲本与F2:3药害指数Fig. 4 Phytotoxicity index of parents and F2:3 in treatments with different glyphosate concentrations |
亲本与后代的抗性均呈正相关: 在亲本和F2:3均为1230 g a.i. hm-2浓度下处理时, 亲本与F2:3的表型变化不明显, 抗性相关系数也最小(0.251)。其他浓度处理下, 亲本与F2:3之间的抗性相关系数都大于0.5, 其中在亲本和F2:3均为3690 g a.i. hm-2浓度下处理时, 亲本与F2:3的相关性最大(0.979), 达到显著水平(表5)。
| 表5 不同草甘膦处理条件下亲本与F2:3对草甘膦抗性的相关系数 Table 5 Correlation coefficient of glyphosate resistance between parents and F2:3 |
F2:3各株系与亲本材料相比抗性普遍较强, 其中平均抗性排名前10位的, 均来源于ZLHJ10-713× ZLHJ06-698和ZLHJ11-142× ZLHJ10-625组合, 在这2个组合后代中, 有较高机率筛选到高抗材料(表6)。
| 表6 基于株高相对变化的草甘膦抗性排序前10位F2:3株系 Table 6 Top 10 F2:3 lines with high glyphosate resistance based on the relative height change |
为避免杂交亲本因外源基因有无对后代抗性产生影响, 本研究选择的杂交亲本均含有草甘膦抗性基因CP4-EPSPs。抗性分级法是用于鉴定转基因材料对草甘膦耐受性最经典最常用的方法, 本试验在采用抗性分级法的同时, 以株高作为指标鉴定抗性, 蒋凌雪等[21]对草甘膦处理后大豆顶芽的全基因表达分析表明, 草甘膦抑制大豆顶芽生长的分子机制为抑制莽草酸途径中色氨酸的生物合成, 只有色氨酸迅速下降并维持较低水平时才能抑制芽伸长, 而色氨酸也是唯一有效的芳香族氨基酸, 说明这一方法具有一定的理论基础。
本研究发现, 低浓度(1230 g a.i. hm-2)处理的药害不明显, 而高浓度处理(3690 g a.i. hm-2, 4920 g a.i. hm-2)与清水对照相比存在极显著差异, 证明高浓度草甘膦对转基因大豆的苗期生长有显著的抑制作用, 与前人报道相同[18]。同时, F2:3对草甘膦的耐受性普遍高于亲本, 这与F2:3具有一定的杂种优势有关, 后续世代耐受性有降低的可能, 但仍需通过试验进一步验证。各亲本中ZLHJ06-1568、ZLHJ10-713和ZLHJ06-698的抗性相对较强, 而各组合F2:3中以ZLHJ10-713 × ZLHJ06-698对草甘膦的抗性较强, 说明利用抗性较高的亲本杂交, 获得高抗后代的几率较大, 但后代抗性并不是双亲的简单累加, 表明抗草甘膦特性这一目标性状的基因表达与遗传背景密切相关, 双亲遗传背景的差异影响着后代遗传倾向, 这一发现对于杂交育种中的亲本选配具有一定的借鉴意义。
本试验中3690 g a.i. hm-2的草甘膦对大豆苗期产生明显药害, 而1230 g a.i. hm-2无明显药害。但有研究发现, 抗草甘膦大豆对1倍、2倍、4倍推荐剂量草甘膦的耐受性均较好, 不同剂量和不同施药时期对大豆产量影响不显著[18], 这可能与其利用莽草酸及产量作为主要指标进行分析、且喷药时期不同有关。草甘膦的推荐用药浓度为1230 g a.i. hm-2, 而在实际生产实践中, 农民为提高除草效率, 往往采用加倍剂量喷施, 有报道表明, 用3倍和4倍生产剂量草甘膦喷施, 对抗草甘膦大豆具有明显的抑制作用[22]。因此, 生产上为保证除草效果, 同时避免除草剂对抗草甘膦大豆造成药害, 建议将草甘磷施用剂量控制在2460 g a.i. hm-2以下。
转基因大豆亲本及其F2:3植株在第一片三出复叶完全展开(V2期)时喷施浓度为1230 g a.i. hm-2的草甘膦, 大豆苗受影响不显著, 而浓度为3690 g a.i. hm-2和4920 g a.i. hm-2的草甘膦则对苗期生长具有显著的抑制作用。不同组合F2:3抗性普遍高于其双亲, 表现出一定的杂种优势。6个亲本中以ZLHJ06- 1568、ZLHJ10-713和ZLHJ06-698的抗性相对较强, 而4个杂交组合F2:3中以ZLHJ10-713× ZLHJ06-698对草甘膦的抗性较强。亲本与其杂交子代对草甘膦的耐受性呈正相关, 利用抗性较高的亲本进行杂交, 获得高抗后代的几率较大, 但后代抗性并不是双亲的简单累加, 由于亲本间互作效应不同, 导致后代抗性水平产生差异, 表明抗草甘膦这一目标基因在大豆中的表达与其遗传背景密切相关。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|