


{kind=link}
{kind=link}
{kind=link}
{kind=link}
利用普通六倍体小麦和西藏半野生小麦杂交衍生的重组自交系定位小麦芒长QTL
[宫希1  , 蒋云峰
, 蒋云峰2 , 徐彬杰2 , 乔媛媛2 , 华诗雨1 , 吴旺1 , 马建2 , 周小鸿2 , 祁鹏飞2, * , 兰秀锦2 ]
, 蒋云峰, 兰秀锦|
|


第一作者联系方式: E-mail: 360035030@qq.com
芒长是普通小麦的重要农艺性状, 受多个基因控制。本研究利用长芒的普通小麦郑麦9023与无芒的西藏半野生小麦Q1028构建一个重组自交系群体(186个株系); 采用SSR和DArT分子标记, 构建覆盖小麦全基因组的遗传图谱(2597 cM)。基于重组自交系群体两年芒长表型数据, 采用ICIM作图法对小麦芒长性状进行QTL定位分析。共检测到2个与芒长相关的QTL, 即 Qwa.sau-4AS和 Qwa.sau-5AL。它们分别位于4AS和5AL染色体上, 可分别解释7.4%和27.3%的表型变异。这2个QTL效应可能分别来源于钩芒基因 Hd与抑芒基因 B1。利用连锁标记进行基因型分析, 表明 Qwa.sau-4AS与 Qwa.sau-5AL对芒长的抑制效果具有累加效应, 且 Qwa.sau-5AL效应强于 Qwa.sau-4AS。本研究将为精细定位及克隆这2个QTL奠定基础。
, JIANG Yun-Feng, LAN Xiu-JinAwn length is an important agronomic trait in hexaploid wheat and controlled by multiple genes. A recombinant inbred line (RIL) population containing 186 lines were developed by crossing Tibetan semi-wild wheat accession Q1028 (female) and common wheat variety Zhengmai 9023, and a genome-wide genetic map (2597 cM) was constructed using SSR and DArT markers. QTLs controlling awn length were identified with two-year phenotypic data and the genetic map by using the Inclusive Composite Interval Mapping (ICIM) method. Two QTLs associated with awn length, designated Qwa.sau-4AS and Qwa.sau-5AL, were detected on chromosomes 4A and 5A, which explained 7.4% and 27.3% of phenotypic variations, respectively. According to genetic locations, effects of the two QTLs might be from Hdand B1 genes that could reduce awn length, respectively. Genetic analysis showed that Qwa.sau-4ASand Qwa.sau-5ALhad cumulative effects, with stronger inhibiting effect in Qwa.sau-5AL than in Qwa.sau-4AS. These results are valuable for fine mapping and cloning target genes controlling awn length in the future.
小麦有芒品种通常比无芒品种的产量高10%以上[1, 2], 因此芒也成为高产育种的考察性状之一。芒作为小麦穗器官的组成部分, 是植物长期进化、适应环境的结果[3], 是小麦光合作用和蒸腾作用的重要器官。芒的绿色细胞富含叶绿体与线粒体, 含量较颖片和外稃更高, 其形态和结构与叶片相似, 具有光合功能[4], 在小麦灌浆后期, 其光合活性高于旗叶[5]。叶片干枯后, 几天之内水分仍可运输到芒, 继续进行光合作用, 光合产物就近转运, 增加籽粒干物质积累[6, 7]。
芒在逆境条件下有助于提高产量[8, 9]。小麦的芒有较多气孔和较高的蒸腾作用, 在干旱条件下可加快水分向麦穗的运输, 使其吸收更多养分, 保证籽粒发育。此外, 小麦芒在种子的传播, 抗御虫害以及防止脱粒等方面也具有重要的作用。白天空气干燥时芒保持弯曲, 而夜晚空气湿度增大引起芒的纤维排列变化, 使芒变直, 每日循环的空气湿度成为芒推动种子进入地面的能量来源, 证明芒在小麦繁殖进化的过程中起了重要的作用[10]。芒表面有坚硬的硅质毛, 不利于害虫的飞落, 降低害虫在麦穗上产卵的机会, 有利于抗御鸟雀和虫害[11]。芒的弹性又可防止穗间相互撞击, 避免籽粒脱落。
小麦芒长由多个基因控制, 其中效应较强且报道最多的是3个基因位点B1、B2和Hd。B1和B2是芒抑制基因, 其中B1抑制作用最强, 可导致穗基部和中部芒极短或无芒[9, 12]。B1位于5AL染色体末端, 与标记Xgwm291的遗传距离为4.99 cM [9]。B2会造成整个穗部的短芒, 它被定位在中国春6BL-5和6BL-6缺失片段末端之间, 与分子标记Xwmc539、Xgpw5130和Xwmc748临近[13]; Hd基因控制钩芒的产生[14], 也会对芒长产生抑制作用, 被定位于4AS染色体Xfba78标记附近[15]。
本研究构建了一个重组自交系群体, 利用两种分子标记构建了遗传图谱, 结合2年芒长表型数据, 定位了2个QTL, 为进一步精细定位芒长相关基因奠定了基础。
郑麦9023 (Triticum aestivum, AABBDD)是一个有芒普通小麦品种, 西藏半野生小麦Q1028 (Triticum aestivum, AABBDD)是一个无芒六倍体材料。它们的芒长差异显著(图1)。以郑麦9023为母本与Q1028杂交, F1代起连续自交, 通过单粒传法获得重组自交系(RIL)。
 | 图1 郑麦9023(有芒)和Q1028(无芒)的穗芒形态Fig. 1 Awn feature of Zhengmai 9023 (with awn) and Q1028 (without awn) |
将186个RIL株系及其亲本分别于2011— 2012 (F11)和2012— 2013 (F12)小麦生长季, 在四川农业大学小麦研究所温江试验田进行表型观测。试验田肥力中等、地力均匀。采用单粒播种, 每系种1行, 行长2 m, 行距0.3 m, 株距0.1 m。10月12日至15日播种, 分蘖前均苗, 使苗与苗之间实际间距大于0.08 m, 抽穗期单穗套袋自交。其他田间管理与大田生产一致。
小麦成熟后, 每株随机取5穗, 用游标卡尺测量侧芒的长度, 取平均值。
取亲本及RIL系(F12)的幼苗叶片, 利用CTAB法提取小麦基因组DNA。根据Rö der等[16]和Somers等[17]发表的小麦遗传图谱, 选取1089对SSR引物用于亲本间全基因组多态性筛选, 其中107对引物在双亲间具有多样性, 用于检测186个RIL系。引物序列信息来源于http://wheat.pw.usda.gov/GG2/index. shtml, 引物由上海英俊生物有限公司合成。PCR扩增程序为94℃变性5 min; 94℃变性1 min; 60℃退火(根据引物不同而调整) 1 min; 72℃延伸1.5 min共40个循环; 72℃延伸7 min后降温到12℃。扩增产物经8%聚丙烯酰胺凝胶电泳检测, 银染显带。DArT芯片由Diversity Arrays Technology Pty. Ltd. (http://www. diversityarrays.com/)开发, 分析检测方法参照Wenzl等[18]的报道。
郑麦9023侧芒平均为6.2 cm, 顶芒为4.0 cm; Q1028无芒。RIL群体不同株系芒长差异巨大。两年的表型数据均呈现明显的双峰分布, 短芒株系的芒长为0.4~1.2 cm; 长芒株系的芒长为2.8~5.2 cm; 极端长芒株系较少, 这表明本群体中芒长性状应该由多基因控制并存在一个效应更强的位点(图2)。
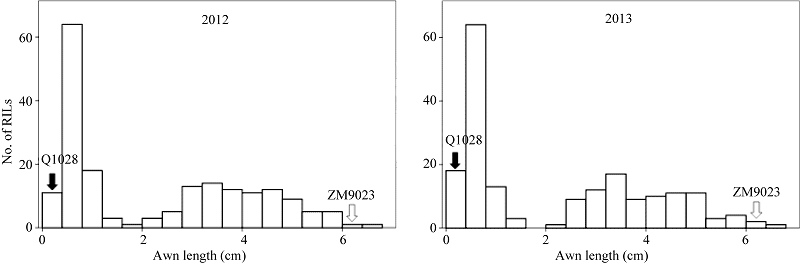 | 图2 重组自交系群体两年芒长直方图Fig. 2 Frequency distribution of awn length in the RIL population for two years |
共检测到2个芒长QTL, 分别位于4AS和5AL染色体, 两年平均可分别解释表型变异的7.4%和27.3% (表1)。位于4AS的QTL被命名为Qwa.sau-4AS。两年的表型数据均将其定位于Xgpw4448附近(图3-a), 相距1.3 cM。位于5AL染色体的QTL被命名为Qwa.sau-5AL, 其贡献率高于Qwa.sau-4AS (表1), 两年的数据均将其定位于wPt-1038附近, 相距1.1 cM (图3-b)。
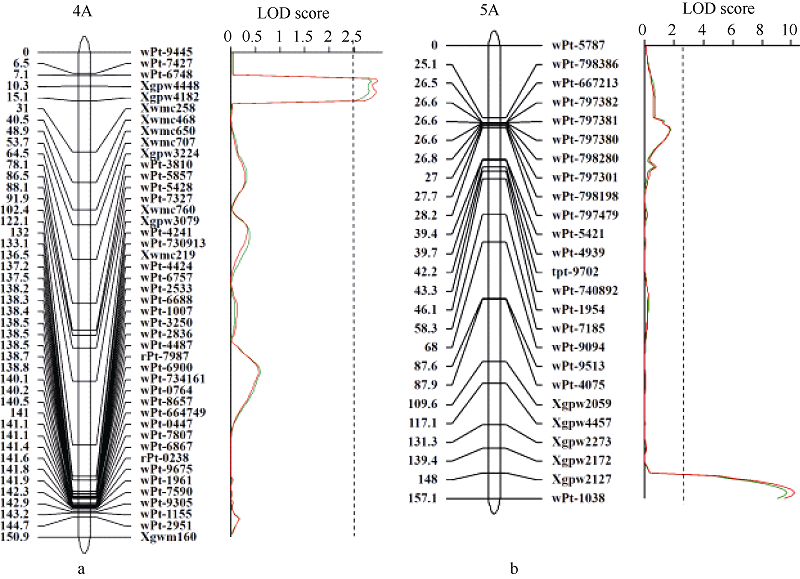 | 图3 4A染色体(a)和5A染色体(b)遗传图谱及芒长性状分析的LOD曲线图 连锁图谱右边是标记名称, 左边是标记位置(cM)。红色和绿色曲线分别为2012和2013年数据; 虚线指示LOD阈值(LOD = 2.5)。Fig. 3 Genetic maps of chromosomes 4A (a) and 5A (b) and the LOD plots of awn length In the linkage map, marker names and positions (cM) are on the right and left, respectively. LOD scores in 2012 and 2013 are shown with the red and green curves, respectivley. The vertical dashed line is the threshold of significance (LOD = 2.5). |
| 表1 郑麦9023 × Q1028群体的芒长QTL Table 1 QTLs for awn length identified in the population of ZM9023 × Q1028 |
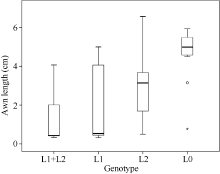
利用与2个QTL连锁的分子标记, 对186个RIL株系进行基因型筛选。去除83个在QTL区间发生重组的株系, 共得到26个同时含有2个QTL的株系, 其平均侧芒长为1.21 cm; 得到24个不含芒长QTL的株系, 其平均侧芒长为4.43 cm; 17个仅含Qwa.sau-5AL位点的株系, 其平均侧芒长为1.71 cm; 37个株系仅含Qwa.sau-4AS位点的株系, 其平均侧芒长为2.97 cm (图4)。
 | 图4 RIL群体不同基因型的芒长箱型分布图Fig. 4 Box plot distribution of awn length in different genotypes of the RIL population L1: Qwa.sau-5AL; L2: Qwa.sau-4AS ; L0: without QTL. |
本研究基于郑麦9023和Q1028构建的RIL群体及其全基因组遗传图谱, 共检测到2个与芒长相关的QTL。其中Qwa.sau-4AS被定位于染色体4AS上的Xgpw4448附近, 与Sourdille等[15]的定位结果相比较, Xgpw4448与Xfba78之间的遗传距离接近, Qwa.sau-4AS应与之前报道的Hd基因位于同一区域, 因此推测该QTL效应来源于钩芒基因Hd。Qwa.sau-5AL与wpt-1038连锁, 与杜斌等[9]的定位结果相比较, wpt-1038与Xgwm291都位于5AL的末端区域, 与B1基因在同一区域, 并表现出同样强烈的芒抑制作用, 因此该QTL的效应应该来源于强抑芒基因B1。
2个抑芒位点均来源于无芒亲本Q1028, 对芒长具有显著抑制, 并存在累加效应。通过比较不同基因型株系的芒长数据发现, 只携带Hd基因的株系平均芒长为2.97 cm, 而只携带B1基因的株系平均芒长为1.71 cm, 说明B1对芒长的抑制效果强于Hd基因。而当同一株系上同时存在这2个基因时, 平均芒长最小。由此可见, B1基因与Hd基因对芒长的抑制效果具有累加效应。这2个位点累计可解释33.9%~35.5%的表型变异, 推测存在未检测出的QTL。
无芒小麦中国春作为一个普通小麦的模式植物, 对其芒长性状已经有较深入研究。普遍认为其无芒特性是由Hd、B1和B2联合作用所致[15]。本试验所采用的无芒亲本为西藏半野生小麦, 许多系统分类及聚类研究已经证明其与中国春具有密切的亲缘关系[22, 23, 24]。未被检测的QTL效应可能源于6BL染色体的B2, 我们采用连锁图谱的6B染色体包含着与B2基因连锁的标记Xwmc182, 但该位点及其所在区域与芒长无显著连锁, 因此未检测到预期的QTL。其原因可能是所采用的2个亲本之间B2基因型相同。
本研究结果为Hd与B1基因后续精细定位与克隆提供了分子标记基础。我们已经利用与钩芒基因Hd连锁的分子标记, 从F8的自交系群体中筛选到杂合单株构建近等基因系群体, 正在进行精细定位相关工作。
共定位2个与芒长性状相关的QTL, 分别位于4AS和5AL染色体, 共解释34.7%的表型变异。这2个QTL应是钩芒基因Hd和抑芒基因B1。2个基因对芒长的抑制作用具有累加效应, 且B1的抑制效果强于Hd。本研究为后续筛选近等基因系及基因精细定位奠定了基础。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|