



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
甘蓝型油菜烷羟化酶基因 MAH1的克隆与表达分析
[徐熠**  , 彭阳
, 彭阳** , 李帅, 赵秋棱, 张双娟, 李加纳, 倪郁* ]
, 彭阳]
|
|


第一作者联系方式: E-mail:714979637@qq.com, 327825919@qq.com
在植物蜡质合成途径中, 中链烷烃羟化酶(mid-chain alkane hydroxylase, MAH) 催化烷烃羟基化形成二级醇, 进一步氧化为酮。本研究以拟南芥P450依赖性酶CYP96A15/MAH1基因为探针, 采用电子克隆与RT-PCR技术, 获得2个甘蓝型油菜 MAH1的全长编码区序列, 分别命名为 BnMAH1-1和 BnMAH1-2(GenBank登录号分别为KT795344和KT795345)。二者ORF长度均为1491 bp, 无内含子, 核苷酸与氨基酸序列分别有92.4%与90.9%的一致性。根据编码区预测的BnMAH1-1和BnMAH1-2前体蛋白均为包含496个氨基酸残基的多肽链, 具有典型的P450蛋白家族保守结构P415xR417x、K螺旋(E359xxR362)、C末端的血红素结合域(F436xxGxRxCxG445) 以及氧结合带保守区域(A/G)G309x(D/E)T312(T/S)。NCBI BlastN、氨基酸序列多重比对与系统学分析表明, 两者与拟南芥 MAH1/ CYP96A15同源性最高。实时荧光定量PCR表明, BnMAH1-1与 BnMAH1-2主要在甘蓝型油菜茎、叶、花、及角果中表达, 其中在叶片中的表达量最高, 在根系中的表达量很低, 这与角质层蜡质主要沉积在植株地上部分相一致。 BnMAH1-1和 BnMAH1-2在无蜡粉材料茎、叶片中几乎不表达, 表明蜡质的减少与 MAH1的转录下调有关。 BnMAH1-1与 BnMAH1-2受SA、MeJA、ACC、ABA、NaCl及干旱胁迫诱导表达, 其中 BnMAH1-1可能在水分胁迫响应中起主要作用。
In wax biosynthesis, the mid-chain alkane hydroxylase (MAH) catalyzes the hydroxylation reaction from alkanes to secondary alcohols and further to corresponding ketones. In this study, using Arabidopsis P450-dependent enzyme CYP96A15/ MAH1 gene as a probe, the full-length coding sequences of two Brassica napus MAH1 genes, BnMAH 1-1 (GenBank accession number KT795344) and BnMAH1-2 (GenBank accession number KT795345), were isolated by in silico cloning and RT-PCR. The ORF lengths of BnMAH1-1 and BnMAH1-2 were 1491 bp, and no intron was included. BnMAH1-1 and BnMAH1-2shared 92.4% and 90.9% of sequence identity at nucleotide and amino acid level, respectively. The predicted BnMAH1-1 and BnMAH1-2 protein contained 496 amino acid residues, with typical P450 protein family conserved domains P415xR417x, K helix (E359xxR362), C-terminal hemopexin-like domain (F436xxGxRxCxG445) and oxygen binding zone (A/G)G309x(D/E)T312(T/S). NCBI BlastN, multiple alignment of amino acid sequence, and phylogenetic analysis showed that they were most homologous to A. thaliana CYP96A15/MAH1. Real-time quantitative PCR showed that BnMAH1-1 and BnMAH1-2 were mainly expressed in stem, leaf, flower, and silique. The highest expression level was found in leaf and the lowest was in root, which was consistent with the wax deposition on aerial part of plant. The expression of BnMAH1-1 and BnMAH1-2was barely detected in stem and leaf of wax deficient cultivar, suggesting that the reduced wax deposition is due to the down-regulation of MAH1 transcription. The expression of BnMAH1-1 and BnMAH1-2was significantly induced by exogenous application of SA, MeJA, ACC, and ABA and exposure to NaCl and drought, among which BnMAH1-1 may play a major role in response to water stress.
植物地上部分表面覆盖的角质层蜡质是植物抵御外界环境胁迫的第一层屏障, 具有多种生理和生态功能, 例如限制植物非气孔性水分散失[1, 2]、防止外源病虫害侵入[3, 4]、阻挡紫外线辐射[5, 6]及降低灰尘、花粉和空气中的污染物在植物表面沉积等[7, 8]。开展植物角质层蜡质代谢、运输和相关调控网络的研究, 对通过育种和栽培手段调控角质层以提高植物抗逆性有重要意义。目前在水稻、番茄、亚麻荠等作物中已有一些利用蜡质相关基因提高作物产量和抗旱性的研究报道[9, 10, 11]。植物角质层蜡质主要是由碳原子数在20~34之间的超长链脂肪酸(VLCFAs)及其衍生物(包括烷烃、醇、醛、酮、酯等)组成, 其中一级醇、酯类等主要由VLCFAs经酰基还原途径生成, 而烷类、二级醇和酮类等组分主要由VLCFAs经脱羰基途径生成[12]。Kolattukudy等[13]对甘蓝叶片的体外前体物质同位素标记试验证实, 烷烃和二级醇能够转化为酮类物质。拟南芥中, 细胞色素P450依赖性酶CYP96A15可催化烷烃羟基化生成二级醇, 进一步氧化成酮, 因此被称为中链烷烃羟化酶(MAH)[14]。
油菜是全世界广泛种植的一种油料作物, 也是我国第五大作物, 具有典型的角质层结构。通常情况下, 甘蓝型油菜叶和茎表皮均有蜡粉覆盖。甘蓝型油菜光叶突变体GL中BnCER1与其他蜡质合成基因表达均下调[15]。白菜型油菜脂质转移蛋白基因BraLTP1在甘蓝型油菜中超表达导致叶片蜡质沉积减少, 并影响细胞分裂和花的发育[16]。MeJA与ACC处理显著诱导甘蓝型油菜中双11角质层蜡质总量与蜡质晶体密度的增加, 蜡质组分中烷类、次级醇类、酮类显著增加[17]。有关油菜蜡质基因功能研究鲜见报道, 本研究克隆甘蓝型油菜中链烷烃羟化酶基因MAH1, 并进行生物信息学分析与基因表达分析, 以期为通过育种和栽培手段调控角质层、提高油菜逆境胁迫的耐受性奠定基础。
甘蓝型油菜中双11与无蜡粉材料NoWax, 由西南大学油菜工程中心提供。将其按常规方法种植, 采集中双11根、茎、叶、花、角果, 储存于-80℃, 用于基因组DNA与RNA的提取。采集NoWax材料的茎、叶用于RNA的提取。大肠杆菌(Escherichia coli)菌株DH5α 感受态购自北京全式金生物技术有限公司。
采用溶液培养(以1/10的MS溶液配制培养液), 待中双11水培幼苗长至5片真叶时, 分别进行激素、盐胁迫与低温胁迫处理。激素处理是分别用200 μ mol L-1水杨酸(SA)、100 μ mol L-1茉莉酸甲酯(MeJA)、20 μ mol L-1脱落酸(ABA)及200 μ mol L-1乙烯合成促进剂1-氨基环丙烷-1-羧酸(ACC)处理6 h。盐胁迫处理是用100 mmol L-1 NaCl溶液处理6 h。低温胁迫是幼苗在4℃下培养24 h。干旱胁迫处理是采用盆栽试验, 待幼苗长至5片真叶时连续7 d干旱, 对照组正常浇水。处理结束后, 取上述材料的叶片保存于-80℃, 用于RNA提取。
采用CTAB法提取基因组DNA。采用TransZol RNA提取试剂盒(TaKaRa)提取总RNA。用DNase I (TaKaRa)消化去除基因组DNA。依照TransScript First-Strand cDNA Synthesis SuperMix反转录试剂盒(北京全式金生物技术有限公司)进行样品mRNA的反转录。
以拟南芥CYP96A15为信息探针, 对甘蓝型油菜基因组数据库(http://www.ncbi.nlm.nih.gov/genome/203)进行同源性Blast检索分析。对检索到的甘蓝型油菜相关序列进行人工拼接与计算机软件预测, 获得包括完整ORF在内的相应基因的序列。
根据电子克隆序列设计PCR引物BnMAH1- 1F/BnMAH1-1R、BnMAH1-2F/BnMAH1-2R (表1), 以中双11的cDNA与基因组DNA分别为模板扩增, PCR体系含10× PCR缓冲液(Mg2+) 5 μ L、2.5 mmol L-1 dNTPs 4 μ L、10 μ mol L-1上下游引物各1.5 μ L、5 U μ L-1Taq酶0.8 μ L、模板2.5 μ L、加ddH2O 34.7 μ L补足50 μ L。其程序为94℃ 2 min; 94℃ 30 s, 55℃ 30 s, 72℃ 1 min, 35个循环; 72℃ 10 min。利用琼脂糖凝胶电泳检测, 回收目的片段, 与载体(pMD-19T Vector)连接, 转化DH5α 感受态, 筛选阳性克隆并测序。
| 表1 甘蓝型油菜MAH1基因克隆引物与荧光定量PCR引物 Table 1 Primers used for B. napus MAH1gene cloning and Real-time fluorescence quantitative PCR |
利用Vector NTI Advance 11. 5软件进行序列比对、翻译及绘制进化树, 在NCBI (http://www.ncbi. nlm.nih.gov/)网站上搜索GenBank Blast, 在http:// bip.weizmann.ac.il/和http://www.expasy.org/进行蛋白质结构、保守域、功能域分析。
根据实际克隆获得的序列设计荧光定量PCR引物BnMAH1-1Fq/BnMAH1-1Rq、BnMAH1-2Fq/ BnMAH1-2Rq (表1), 在qTOWER2.2荧光定量PCR仪(德国耶拿)上扩增。按照ChamQ SYBR qPCR Master Mix试剂盒(Vazyme)说明配制反应体系, 含2× ChamQ SYBR qPCR Master Mix 10.0 μ L、10 μ mol L-1上下游引物各0.4 μ L、cDNA 2.0 μ L、ddH2O 7.2 μ L。扩增程序为95℃ 30 s; 95℃ 5 s, 56℃ 30 s, 40个循环。Actin7作为内参基因, 设置3个生物学重复, 2个技术重复。
参考倪郁等[17]方法采集甘蓝型油菜展开叶片, 于含氯仿的大试管内浸提1 min, 氯仿中预先加入已知浓度的C24烷作为内标。用氮吹仪蒸干浸提液, 再于70℃下用20 μ L BSTFA与20 μ L吡啶衍生45 min, 再次用氮吹仪蒸干, 将产物溶于200 μ L氯仿中。用气相色谱仪(福立9790II, 浙江)分析测定蜡质组分及含量。气相色谱的毛细管柱长30 m, 直径为0.32 mm, 液膜厚度0.25 μ m; 以氮气作为载气; 使用FID (火焰离子化检测仪)为检测器; 柱膜和FID检测器的温度分别为300℃和320℃; 使用的升温程序为初温80℃, 保持0.1 min, 后以15℃ min-1升至245℃, 保持5 min, 再以19℃ min-1升至325℃, 保持12 min。根据FID的峰面积确定蜡质的量, 利用GC-MS (岛津GC2010MS, 日本)鉴定蜡质组分, 参考内标正二十四烷计算各蜡质组分的实际含量(μ g cm-2)。利用数字化扫描仪(EPSON V750)和WinFOLIA叶片专业图像分析软件(Regent Instrument Inc, Canada)测定并记录叶面积。每处理4个重复。
将叶片干燥后用碳导电胶布固定在载物台上, 放入金属离子溅射仪(E1010, Hitachi)内喷镀导电膜, 于扫描电子显微镜(S3000-N, Hitachi)下观察镀金材料的形态。每处理4个重复。
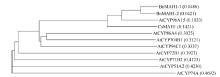
克隆到的2条序列ORF长度均为1491 bp, 无内含子, 编码496个氨基酸, 被分别命名为BnMAH1-1 (GenBank登录号为KT795344)和BnMAH1-2 (GenBank登录号为KT795345)。二者的核苷酸序列一致性为92.4%, 氨基酸序列一致性为90.9%。GenBank Blast搜索表明, BnMAH1-1和BnMAH1-2与拟南芥CYP96A15最同源。将推导的BnMAH1-1与BnMAH1-2蛋白与拟南芥P450家族成员以及亚麻荠(Camelina sativa) MAH1蛋白进行氨基酸序列多重比对(图1)。BnMAH1-1、BnMAH1-2与拟南芥CYP96A15最同源, 分别有77.7%/85.7%、80.7%/ 88.8%的序列一致性/相似性, 其次是亚麻荠CsMAH1, 序列一致性/相似性为66.8%/77.0%、67.2%/76.8%。系统进化分析(图2)表明, BnMAH1-1与BnMAH1-2首先与拟南芥CYP96A15聚在一起形成一个亚组, 然后与亚麻荠MAH1聚在一起, 之后依次与拟南芥CYP86家族的其他亚家族成员、CYP其他多家族成员聚在一起, 最后与CYP单家族成员聚集。蛋白质序列同源比对发现, BnMAH1-1、BnMAH1-2具有典型的P450蛋白家族保守结构域, 即P415xR417x结构域、K螺旋(E359xxR362)以及C末端的血红素结合域(F436xxGxRxCxG445), 同时还具有高度保守的(A/G)G309x(D/E)T312(T/S)序列, 是一个含苏氨酸(T)的氧结合带保守区域(图1)。
 | 图1 甘蓝型油菜BnMAH1-1、BnMAH1-2推测的氨基酸序列与其他植物的P450家族氨基酸序列比对参加比对的有拟南芥P450家族成员AtCYP96A15(AT1G57750)、AtCYP86A4(AT1G01600)、AtCYP94C1(AT2G27690)、AtCYP72B1 (AT2G26710)、AtCYP71B2(AT1G13080)及亚麻芥CsMAH1(KJ461887)。图中完全相同、基本相同与相似程度低的氨基酸残基分别用黑色、深灰色与浅灰背景表示, 不相似氨基酸残基无背景色; 下画线加着重号表示的是细胞色素P450酶系中的保守结构域。Fig. 1 Alignment of deduced amino acid sequences of BnMAH1-1 and BnMAH1-2 with other P450 proteinsThe P450 proteins (accession numbers in parentheses) AtCYP96A15 (AT1G57750), AtCYP86A4 (AT1G01600), AtCYP94C1 (AT2G27690), AtCYP72B1 (AT2G26710) and AtCYP71B2 (AT1G13080) from A. thaliana, and MAH1 (KJ461887) from C. sativaare aligned. In the panel, identical, conservative, weakly similar and non-similar residues are denoted with the background of dark, dark gray, light gray and white, respectively. In consensus line the conserved residues and domains are underlined and doted. |
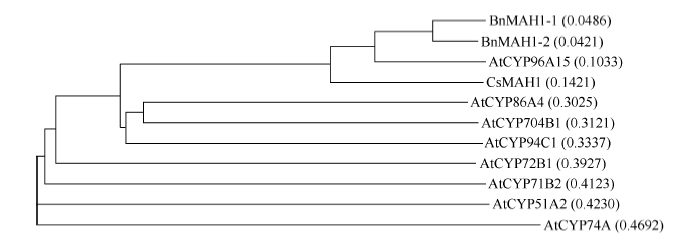 | 图2 BnMAH1基因的系统发生树AtCYP96A15(AT1G57750)、AtCYP86A4(AT1G01600)、AtCYP94C1(AT2G27690)、AtCYP72B1(AT2G26710)、AtCYP71B2(AT1G13080)、AtCYP51A2(AT1G11680)、AtCYP74A(AT5G42650)、AtCYP704B1(AT1G69500)来自拟南芥P450家族, CsMAH1(KJ461887)来自亚麻芥。Fig. 2 Phylogenetic tree of BnMAH1AtCYP96A15 (AT1G57750), AtCYP86A4 (AT1G01600), AtCYP94C1 (AT2G27690), AtCYP72B1 (AT2G26710), AtCYP71B2 (AT1G13080), AtCYP51A2 (AT1G11680), AtCYP74A (AT5G42650), and AtCYP704B1 (AT1G69500) are from P450 proteins of A. thaliana, and CsMAH1 (KJ461887) from C. sativa. |
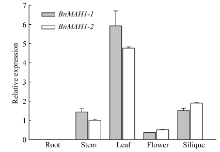
实时荧光定量PCR结果显示(图3), BnMAH1-1与BnMAH1-2主要在甘蓝型油菜茎、叶、花及角果中表达, 其中在叶片中的表达量最高, 其次是在角果和茎中, 而在根系中的表达量最低。BnMAH1-1的表达量在茎和叶片中高于BnMAH1-2, 在花和角果中略低于BnMAH1-2。
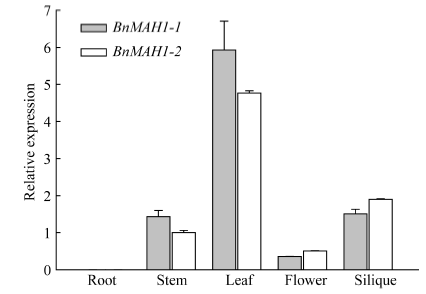 | 图3 BnMAH1-1与BnMAH1-2在甘蓝型油菜不同组织器官中的表达分析Fig. 3 Tissue-specific expression ofBnMAH1-1 and BnMAH1-2 in various organs of B. napus |
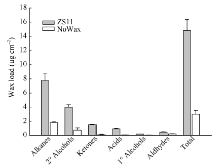
甘蓝型油菜中双11叶表皮蜡质晶体结构以杆状、颗粒状(小型片状)为主, 而无蜡质材料NoWax叶表面无蜡质晶体结构覆盖(图4)。GC-MS分析结果表明, 与中双11相比, NoWax材料叶角质层蜡质各组分含量以及蜡质总量均显著下降, 其中酮含量下降92.87% (图5)。由图6可知, BnMAH1-1和BnMAH1-2在ZS11茎、叶片中均有表达, 而在NoWax材料茎中表达量很低, 在叶片中几乎不表达。
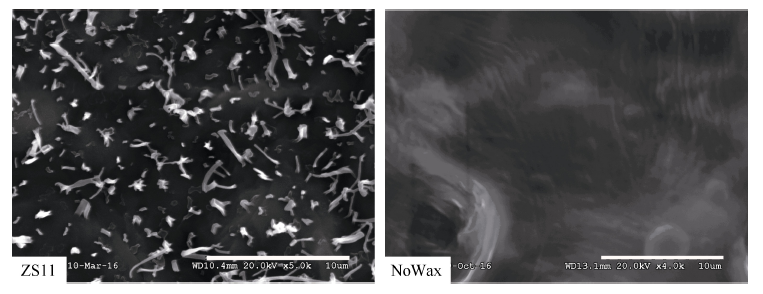 | 图4 表皮蜡质的扫描电镜分析(标尺=10 μ m)Fig. 4 Scanning electron microscopy analysis of epicuticular wax deposition on leaves (bar=10 μ m) |
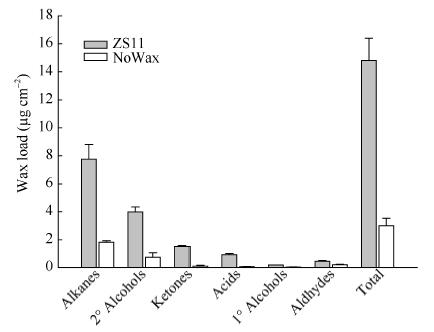 | 图5 ZS11与NoWax叶片角质层蜡质的组成与含量Fig. 5 Cuticular wax amount and composition in leaves of ZS11 and NoWax |
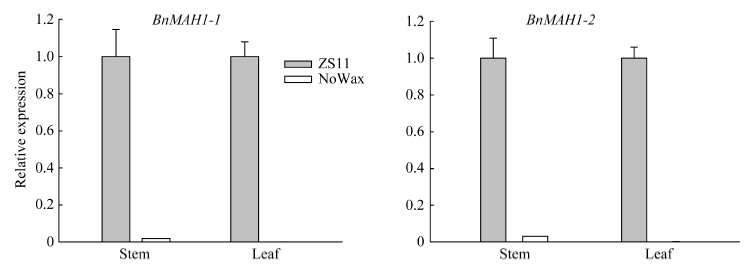 | 图6 BnMAH1-1与BnMAH1-2在有蜡粉材料ZS11与无蜡粉材料NoWax中的差异表达Fig. 6 Differential expression of BnMAH1-1 and BnMAH1-2 in ZS11 and NoWax |
荧光定量PCR结果表明(图7), SA、MeJA、ACC以及ABA均不同程度地诱导BnMAH1-1和BnMAH1-2的表达, 其中MeJA、ABA诱导BnMAH1-1表达上调10倍以上。NaCl及干旱胁迫下, BnMAH1-1和BnMAH1-2表达均显著上调, 而低温胁迫下BnMAH1-1和BnMAH1-2表达均显著下调。
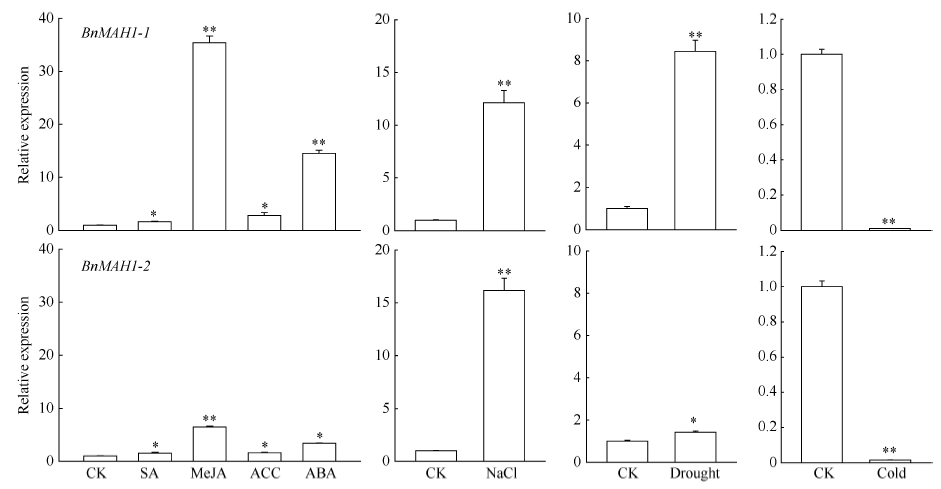 | 图7 激素与逆境胁迫对BnMAH1-1与BnMAH1-2基因表达的影响数据柱上* 与* * 分别表示在P< 0.05或P< 0.001水平上差异显著。Fig. 7 Effects of hormones and stresses on the expression of BnMAH1-1 and BnMAH1-2 in B. napusAsterisk and double asterisks above data bars indicated significance at P< 0.05 and P< 0.001, respectively according to Student’ s t-test. |
植物角质层是植物与外界环境的第一接触面, 是植物应对生物和非生物逆境的重要屏障, 其中的角质层蜡质是由堆积在角质层最外层有独特三维结构的表皮蜡质和填充于角质中的内部蜡质组成, 具有多种生理和生态学功能。角质层蜡质主要由VLCFAs、烷烃、一级醇、二级醇、酮、醛、酯等组成。前人利用反向遗传学的方法在拟南芥茎中发现了一个与二级醇和酮合成相关的细胞色素P450依赖性酶CYP96A15, 称之为中链烷烃羟化酶(MAH1)[14]。P450基因是一个古老的超基因家族, 基于蛋白序列的相似性和亲缘关系, 陆生植物P450可分为11簇, 归为单家族簇(CYP51、CYP74、CYP97、CYP710、CYP711、CYP727、CYP746)和多家族簇(CYP71、CYP72、CYP85、CYP86) 2类。拟南芥中发现有272个P450基因, 其中CYP86家族簇包含33个基因, 分别聚类在CYP86、CYP94、CYP96和CYP704家族中[18, 19]。拟南芥CYP96家族中包含8个亚家族, 分别为CYP96A、CYP96B、CYP96C、CYP96D、CYP96E、CYP96F、CYP96G和CYP96H, 其中CYP96A亚家族包含15个基因成员, 到目前为止, 只有CYP96A15被发现与角质层蜡质合成有关, 最终被鉴定为MAH。本研究以拟南芥CYP96A15基因为探针, 从甘蓝型油菜中克隆了2个MAH1基因BnMAH1-1和BnMAH1-2。在核苷酸水平与氨基酸水平上, 这2个基因与拟南芥CYP96A15同源性最高、与亚麻荠MAH1其次。多重比对及系统发生树结果表明, BnMAH1-1和BnMAH1-2首先与CYP96A15聚在一起, 其次与CYP86, 再次与其他P450家族成员聚在一起。前人研究表明, 细胞色素的三维结构非常保守, 其中保守的血红素结合域FxxGxRxCxG序列是鉴定P450类蛋白的主要特征[20, 21]。BnMAH1-1和BnMAH1-2氨基酸序列结构分析表明, 它们在第436至第445个氨基酸处均含有血红素结合域FxxGxRxCxG (图1)。该结构域中存在与铁元素形成硫醇盐离子键的绝对保守的半胱氨酸(C)残基。此外, BnMAH1-1和BnMAH1-2中还含有P450的其他保守结构域P415xR417x、K螺旋(E359xxR362)及氧结合带保守序列(A/G)G309x(D/E)T312(T/S), 它们可能在蛋白质的折叠、组装中起关键作用[18, 22]。这些结果表明, 本研究所克隆到的BnMAH1-1和BnMAH1-2序列是编码中链烷烃羟化酶的基因序列, 是对应于拟南芥CYP96A15的2个垂直同源基因。
陆生植物地上大部分器官都有角质层蜡质覆盖, 蜡质是植物功能器官的重要结构。本研究结果表明, BnMAH1-1与BnMAH1-2基因主要在甘蓝型油菜茎、叶、花、角果中表达, 其中在叶片中的表达量最高, 在根系中表达量很低(图3)。这与角质层蜡质主要沉积在植物地上部分的特性相一致。BnMAH1-1和BnMAH1-2在无蜡质材料茎、叶片中几乎不表达, 暗示了蜡质的减少与BnMAH1的转录下调有关。与中双11相比, NoWax材料叶角质层蜡质中包括酮类在内的各组分含量以及蜡质总量均显著下降(图4和图5), 而在蜡质合成基因表达方面, 除MAH1外, 负责烷类合成的CER1、负责一级醇合成的CER4等的转录水平也都下调。由BnMAH1、BnCER1、BnCER4等基因各成员启动子同时发生突变而导致这些蜡质合成基因转录水平下调的可能性几乎为零, 因此很有可能是调控蜡质合成基因表达的上游信号基因发生改变, 进而抑制蜡质合成基因表达。BnMAH1-1和BnMAH1-2可能活跃地参与了甘蓝型油菜地上部分, 尤其是叶片角质层蜡质的沉积。
角质层蜡质的沉积除受发育状况的控制外, 还对水分、光强、寒冷、季节变化等环境信号响应[23]。拟南芥叶片角质层蜡质在盐、干旱胁迫以及施用外源ABA后增加[24], 而在黑暗中减少[25]。当玉米四叶期被低温处理7 d时, 其第三片叶角质层蜡含量减少29%[26]。进一步在分子水平上的研究发现, 拟南芥蜡质合成基因KCS、CER10、KCR1在盐、水分胁迫下转录水平上调, 而在低温与黑暗条件下转录水平下调[27]。本实验中, NaCl与干旱胁迫显著诱导了BnMAH1-1和BnMAH1-2的表达水平上调, 低温胁迫则显著下调了BnMAH1-1和BnMAH1-2的表达, 表明BnMAH1是甘蓝型油菜响应逆境胁迫的蜡质基因。
植物在遭受各种生物和非生物胁迫时, 体内激素水平会发生变化以启动和调节某些与逆境适应相关的生理生化过程来诱导抗(耐)逆性的形成。ABA、ETH、SA、JA都是与植物逆境胁迫相关的激素。本试验表明, ABA、SA、MeJA以及ACC不同程度地诱导了BnMAH1-1和BnMAH1-2的转录上调, 其中MeJA与ABA的诱导效果最显著。前人研究报道, ABA可诱导独行菜叶片中大于C26的脂肪族物质含量的增加, 其耐旱性随之增强, 这可能是由于ABA对蜡质合成基因KCS6的诱导[28]。拟南芥在水分胁迫下, ABA通过上调转录因子MYB96的表达而促进蜡质合成基因KCS1、KCS2、KCS6与KCR1的表达, 最终促进角质层蜡质的积累[29]。这些结果表明, 植物激素作为信号分子可以调控蜡质合成基因的表达从而影响蜡质沉积, 最终影响植物对逆境胁迫的响应。本试验中, 干旱胁迫与ABA极大地诱导了BnMAH1-1的表达上调, 表明BnMAH1-1可能在响应水分胁迫中起主要作用。
克隆了2个甘蓝型油菜中链烷烃羟化酶基因BnMAH1-1和BnMAH1-2, 它们是对应于拟南芥CYP96A15/MAH1的垂直同源基因; BnMAH1-1与BnMAH1-2主要在甘蓝型油菜地上部分组织器官中表达, 在叶片中的表达量最高, 在无蜡粉材料茎、叶片中几乎不表达, 暗示蜡质的减少与MAH1的转录下调有关, BnMAH1-1和BnMAH1-2可能活跃地参与甘蓝型油菜地上部分, 尤其是叶片角质层蜡质的沉积。BnMAH1-1与BnMAH1-2受SA、MeJA、ACC、ABA、NaCl及干旱胁迫诱导表达, 其中BnMAH1-1可能在水分胁迫响应中起主要作用。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|