



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻斑点叶突变体 splZ97的生理特性及其基因定位
[韦荔全1, ** , 罗延敏1, ** , 王文强1 , 池长程1 , 黄福灯2 , 向珣1 , 程方民1 , 潘刚1, *  ]
]
]
|
|


通过EMS诱变籼稻恢复系珍97获得一个稳定遗传的褐色斑点叶突变体 splZ97(spotted leaf Z97, splZ97)。大田条件下, 突变体 splZ97的斑点叶性状始于分蘖期, 此后由叶缘向叶中下部迅速扩散, 直至整个叶片, 严重时叶片部分或整体枯死, 从而致使突变体株高、每穗粒数及结实率极显著低于野生型对照。生理分析表明, 与野生型珍97相比, 孕穗期突变体 splZ97剑叶、倒二叶和倒三叶的叶绿素含量极显著降低, 而POD (peroxidase, POD)活性、O2܋含量及MDA (malondialdehyde, MDA)含量升高; 突变体 splZ97倒二叶和倒三叶的CAT (catalase, CAT)活性和可溶性蛋白含量极显著降低, 而SOD (superoxide dismutase, SOD)活性则极显著增加。组织化学分析进一步证实, 突变体 splZ97的叶片明显累积O2܋。此外, 突变体 splZ97苗期经盐胁迫处理后, 其株高及根长明显受到抑制。遗传分析表明, 突变体 splZ97的斑点叶性状受一对隐性核基因控制, 借助图位克隆技术将该基因定位于第12染色体长臂的RM28466与RM28485两个SSR标记之间, 物理距离为189 kb, 该结果为进一步克隆 SPLZ97基因并研究其功能奠定了基础。
A spotted-leaf mutant splZ97 was isolated from a mutant bank generated by EMS mutagenesis of indica restore line Zhen 97. Under field conditions, the brown lesion-mimics mutant splZ97 firstly displayed at the tip and edge of leaf blade at tillering stage, and then gradually spread to whole leaf, resulting in the death of the whole blade when the symptom was severe. At the same time, the major agronomic traits including plant height, grain number per panicle and seed-setting rate were markedly affected. Compared with the wild-type the flag leaf, the second leaf from top and the third leaf from top at heading stage, chlorophyll contents in the mutant splZ97 significantly decreased, while POD (peroxidase, POD) activity, O2܋ level and MDA (malondialdehyde, MDA) content increased. In addition, CAT (catalase, CAT) activity and soluble protein content of the second leaf from top and the third leaf from top of the mutant decreased as compared with the wild type; on the contrary, the SOD (superoxide dismutase, SOD) activity significantly increased. The histochemical analysis further indicated that O2܋ accumulated in the leaf blade of the mutant splZ97. In addition, under salt stress at seedling stage, the shoot length and root length of the mutant splZ97 were significantly shorter than these of the wild type. Genetic analysis and gene mapping showed that splZ97 was controlled by a single recessive nuclear gene, which was mapped to a region of 189 kb flanked by two SSR markers RM28466 and RM28485 on the long arm of chromosome 12. These results achieved in the present study would further facilitate the cloning and functional analysis of the gene SPLZ97.
植物斑点叶(spotted-leaf)、类病变(lesion mimic)或类病斑(lesion simulating disease)突变体是指突变体植物在没有受到明显逆境、机械和农药损伤或病原菌侵染的条件下, 植物叶片、叶鞘甚至茎秆等部位自发形成类病原菌侵染或类过敏反应的程序性细胞死亡现象[1, 2]。研究表明, 斑点叶突变体还与植物抗病防卫反应密切相关[3]。因此, 斑点叶突变体是研究植物程序性细胞死亡及病害响应机理的理想材料[2, 3]。
关于斑点叶的形成机理, 基于不同突变体的分子生理生化研究, 认为斑点叶的形成不仅受外因, 如光[4]和温度[5]的诱导; 更受内因, 如激素和ROS (reactive oxygen species, ROS)等信号因子[6, 7, 8], 以及基因的精细调控[7, 8]。利用正向和反向遗传学手段, 迄今已从水稻中克隆并经遗传转化或T-DNA插入突变体验证的叶片类病变形成的相关基因有18个, 根据基因功能可以将其分成6类, 包括转录因子, 如SPL7[9]和OsLSD1[10]; 蛋白酶或激酶基因, 如OsMAPKKK1/ OsEDR1/OsACDR1/SPL3[11]和NRKe[5]; 参与代谢加工降解的基因, 如SGR[12]、UAP1/SPL29[13]、CYP71P1/SL[14]、SPL11[15]、OsAT1/SPL18[16]、RLS1[17]、SPL5[18]、LLB[19]等; 物质转运相关基因, 如OsPLS1[20]、SPL28[21]、LMR[22]等; 蛋白或RNA结合蛋白基因, 如GE14e[23]和OsLMS[24]; 生物合成相关基因, 如RLIN1[4]。
本课题组利用EMS诱变籼稻恢复系珍97, 获得一个隐性斑点叶突变体, 暂命名为splZ97 (spotted leaf Z97, splZ97)。该突变体斑点叶表型始于水稻分蘖期(约6~7叶期), 之后随着叶龄的增加, 除上部1~2片展开叶及心叶外, 其他叶片均不同程度出现斑点叶性状; 抽穗开花期除剑叶外所有叶片均出现斑点叶性状。本文对突变体splZ97基本表型、生理变化及基因定位等方面的研究, 为进一步克隆该基因并揭示其斑点叶分子生理机理奠定了基础。
EMS诱变籼稻恢复系珍97, 从诱变后代中获得斑点叶突变体splZ97, 经浙江杭州和海南连续6代回交和自交, 获得斑点叶性状稳定株系。之后以splZ97为母本, 分别与原始野生型对照珍97、粳稻品种02428和秀水110杂交获得F1, F1自交获得的F2群体用于遗传分析及基因定位。所有材料均种植于浙江大学紫金港农业试验站。2014年和2015年在水稻成熟后, 分别取splZ97与珍97各20株, 调查其株高、穗长、有效穗数、每穗粒数、结实率和千粒重等主要农艺性状。2015年在水稻生长至孕穗期, 取突变体splZ97及其野生型珍97的剑叶、倒二叶和倒三叶, 测定有关生理指标; 同时, 对F1及其F2群体分单株观察其斑点叶表型并取样, 提取DNA用于后续基因定位分析。
选取突变体splZ97及其野生型对照珍97的成熟种子并脱壳, 经75%酒精消毒1 min及10%次氯酸钠溶液消毒20 min, 之后灭菌水洗涤5次, 每次2 min。将消毒好的种子接种在含0、100和200 mmol L-1 NaCl的1/2 MS[25]固体培养基上, 于水稻组织培养室培养6 d后统计根长及株高并拍照。培养条件为光照16 h, 黑暗8 h, 温度为28℃。
选取突变体splZ97及其野生型对照珍97各10株处于孕穗期(剑叶叶枕与倒二叶叶枕平齐的分蘖)的剑叶、倒二叶和倒三叶, 分别测定叶绿素、可溶性蛋白、过氧化氢(H2O2)、超氧阴离子(O2܋)、MDA含量以及CAT、SOD和POD活性。用80%丙酮于黑暗条件下浸泡叶片至发白, 将浸提液稀释5倍, 以80%丙酮为空白, 在波长663 nm和646 nm下测定其光密度, 计算叶绿素含量; 采用考马斯亮蓝G-250染色法测定可溶性蛋白; 采用碧云天生物技术有限公司的过氧化氢检测试剂盒(s0038)测定H2O2含量; 参考《植物生理学实验技术》的相关方法测定其他生理指标[26]。采用日本岛津公司UV-2450紫外分光光度计, 每个指标测定5个生物学重复。
选取分蘖盛期突变体splZ97及其野生型对照珍97主茎倒二叶中上部3~5 cm长叶片, 分别参考Kariola等[27]和Mahalingam等[28]的方法进行NBT及DAB染色。每个试验重复3次。
大田栽培条件下, 首先分别观察splZ97/珍97、splZ97/02428和splZ97/秀水110的杂种F1叶片表型, 确定控制突变体splZ97斑点叶性状的显隐性; 其次, 根据F2群体中具有正常叶色的水稻单株数与具有斑点叶表型的单株数的比例, 确定控制斑点叶性状的基因数量。同时, 分单株剪取splZ97/02428的F2定位群体中具有斑点叶性状的单株、splZ97、02428以及F2群体中10株正常植株的叶片, 采用CTAB法提取基因组总DNA。利用均匀分布于水稻12条染色体的500对SSR标记, 其引物序列来自Gramene数据库(http://www.gramene.org/), 以及Shen等[29]所开发的50对InDel标记进行基因定位。PCR总反应体系为20 μ L, 内含0.8 U TaqDNA聚合酶, 1 × PCR buffer (Mg2+Plus), 1 mmol L-1 dNTP Mixture, 50 ng DNA, 上下游引物各0.25 μ mol L-1。PCR反应条件为95℃预变性5 min; 94℃, 30 s; 55℃, 30 s; 72℃, 30 s; 共35个循环; 72℃, 10 min。PCR产物经8%非变性聚丙烯酰胺凝胶电泳, 快速银染后观察[30]。
从splZ97/02428 F2群体中的853个单株中获得207株具有斑点叶性状的单株组成基因定位群体, 利用上述筛选到的基因连锁分子标记进行定位群体的遗传分析, 获得每个标记的重组交换单株数。同时在RAP-DB数据库(http://rapdb.dna.affrc.go.jp/)查找基因连锁分子标记在染色体上的具体物理位置以确定其排列顺序, 结合每个标记的重组交换单株数确定SPLZ97基因与连锁标记的顺序, 构建基因定位图谱。
突变体splZ97叶片在分蘖期(6~7叶期)就表现斑点叶性状(图1-A), 而后除上部1~2片展开叶及心叶外, 其他叶片的褐色斑点始于叶尖及边缘, 并向叶中部及下部迅速蔓延, 严重时叶中上部枯死。而且自分蘖开始至抽穗期, 相对于野生型珍97, 突变体splZ97叶片的叶色明显更黄(图1-B, C)。突变体splZ97生长至抽穗开花前期, 除剑叶外所有叶片均不同程度出现斑点叶症状(图1-D, E, F)。由于叶片出现类病变症状, 导致突变体splZ97的主要农艺性状, 如株高、穗长、每穗粒数和结实率分别比野生型对照珍97下降25.16%、12.78%、47.29%和42.60% (表1)。
 | 图1 突变体及其野生型的表型A: 分蘖期; B: 抽穗期; C: 抽穗期叶片; D: 开花期; E: 开花期叶片; F: 开花期倒二叶; 1~4代表剑叶至倒四叶。Bar=20 cm。Fig. 1 Phenotype of splZ97 and its wild-type (WT) plants at different growth stagesA: tillering stage; B: heading stage; C: leaves at heading stage; D: flowering stage; E: leaves at flowering stage; F: the 2nd leaf from top at the flowering stage; 1-4 mean the 1st leaf to 4th leaf from top. Bar=20 cm. |
| 表1 突变体及其野生型的农艺性状 Table 1 Agronomic traits ofsplZ97 and its wild-type (WT) plants |
正常条件下, 突变体splZ97与野生型珍97之间的株高和根长均无明显差异; 而在盐胁迫条件下, 两者的均受到抑制, 但与野生型相比, 突变体splZ97的根长和株高抑制更明显(图2-A)。与突变体splZ97对照相比, 经100 mmol L-1 NaCl处理后的突变体株高和根长分别降低19.70%和21.76%; 而经200 mmol L-1 NaCl处理的则分别下降26.71%和56.68% (图2-B, C)。
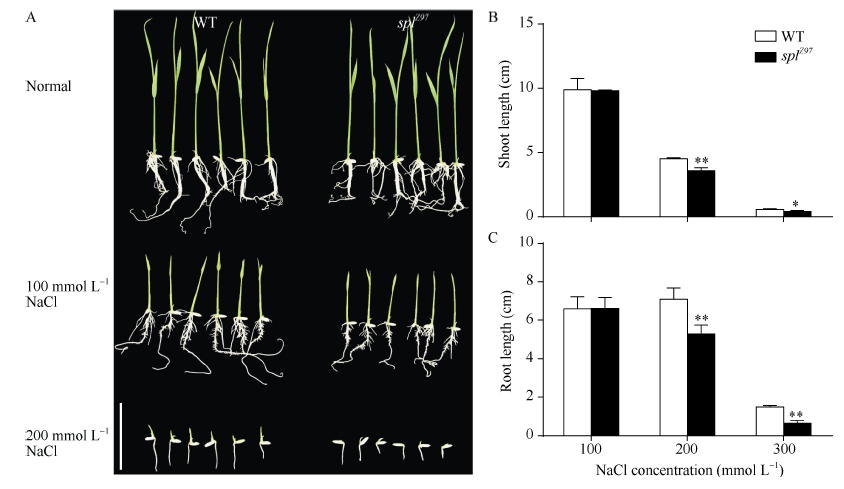 | 图2 盐胁迫对突变体splZ97及其野生型的生长影响A: 突变体splZ97及其野生型的表型, Bar=5 cm; B: 芽长; C: 根长。* 在0.05水平上差异显著; * * 在0.01水平上差异显著。Fig. 2 Response ofsplZ97 mutant and its wild-type (WT) to NaCl stressA: phenotype of splZ97 and its WT plants, Bar=5 cm; B: Shoot lengths; C: primary root lengths of seedlings. * Significantly different at P< 0.05; * * Significantly different at P< 0.01 (t-test). |
2.3.1 叶绿素含量 孕穗期突变体splZ97的剑叶、倒二叶和倒三叶的叶绿素a (图3-A)、叶绿素b (图3-B)、叶绿素总含量(图3-C)及叶绿素a/b比值(图3-D)依次下降且显著低于野生型, 其中突变体splZ97的倒二叶和倒三叶的总叶绿素含量分别比其剑叶下降7.47%和39.46% (图3-C)。与野生型相比, 突变体splZ97的剑叶、倒二叶和倒三叶的总叶绿素含量分别下降43.18%、47.62%和58.59% (图3-C)。
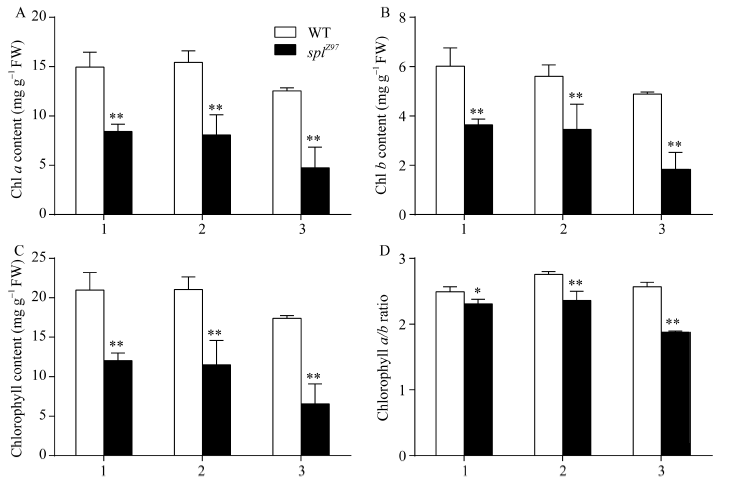 | 图3 孕穗期突变体及其野生型叶片的叶绿素含量1: 剑叶; 2: 倒二叶; 3: 倒三叶。* 在0.05水平上差异显著; * * 在0.01水平上差异显著。Fig. 3 Chlorophyll content of leaves in splZ97 and its wild-type (WT) plants at the booting stage1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top. * Significantly different at P< 0.05; * * significantly different at P< 0.01 (t-test). |
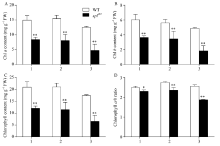
2.3.2 POD、CAT和SOD的活性 图4-A显示, 相对于野生型对照珍97, 突变体splZ97的剑叶、倒二叶与倒三叶的POD活性均极显著增加, 分别为113.49%、982.69%和929.22%。图4-B显示, 野生型对照珍97的剑叶与倒二叶间的CAT活性无显著性差异, 但倒三叶则分别比剑叶与倒二叶下降11.72%和8.58%; 而突变体splZ97的剑叶、倒二叶与倒三叶间的CAT活性则依次极显著下降, 且其倒二叶与倒三叶的CAT活性极显著低于野生型珍97, 分别下降21.16%和37.00%。图4-C显示, 突变体splZ97的剑叶、倒二叶与倒三叶间的SOD活性无显著性差异, 而野生型珍97则依次下降。与野生型珍97相比, 突变体splZ97的倒二叶与倒三叶的SOD活性分别增加14.75%和23.13%。
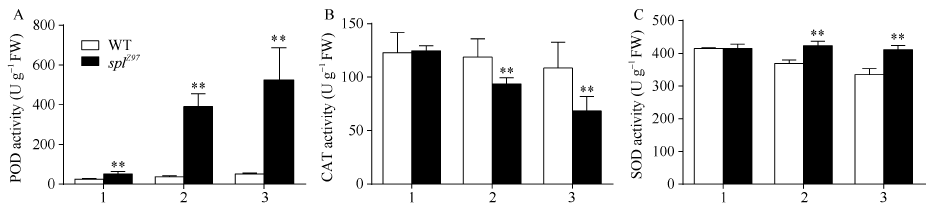 | 图4 孕穗期突变体splZ97及其野生型叶片的POD、CAT和SOD活性1: 剑叶; 2: 倒二叶; 3: 倒三叶。* * 在0.01水平上差异显著。Fig. 4 POD, CATs, and SOD activities of splZ97and its wild-type (WT) plants at booting stage1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top. * * Significantly different at P< 0.01 (t-test). |
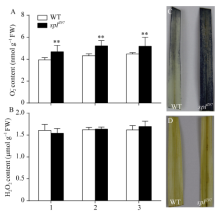
2.3.3 超氧阴离子(O2܋)和过氧化氢(H2O2)的累积
图5-A显示, 野生型对照珍97和突变体splZ97倒二叶和倒三叶间的O2܋含量差异不显著, 但显著高于剑叶。与野生型相比, 突变体splZ97剑叶、倒二叶和倒三叶的O2܋含量分别增加15.71%、17.31%和13.66%。图5-B结果显示, 野生型及突变体的剑叶、倒二叶和倒三叶间的H2O2含量无显著差异。同时, 抽穗期突变体splZ97剑叶的部分组织细胞明显被NBT染成更多更深的蓝色(图5-C), 进一步证实其体内累积O2܋; 而突变体splZ97及其野生型对照叶片组织均没有被DAB染成明显的棕红色(图5-D), 证实两者之间的H2O2含量无明显差异。
 | 图5 孕穗期突变体splZ97及其野生型叶片的O2܋和H2O2累积1: 剑叶; 2: 倒二叶; 3: 倒三叶。* * 在0.01水平上差异显著。C: NBT染色。D: DAB染色。Fig. 5 Accumulation of O2܋and H2O2 in splZ97 and its wild-type (WT) plants at booting stage1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top. * * Significantly different at P< 0.01 (t-test). C: NBT staining.D: DAB staining. |
2.3.4 MDA和可溶性蛋白含量 图6-A显示, 与野生型相比, 突变体剑叶、倒二叶和倒三叶的MDA含量分别增加29.63%、34.48%和56.25%, 达极显著水平; 且突变体splZ97倒二叶和倒三叶的MDA含量分别比剑叶增加11.42%和42.86%。图6-B显示, 野生型和突变体剑叶、倒二叶和倒三叶间的可溶性蛋白含量依次降低, 尤其是两者倒三叶的含量极显著低于其剑叶和倒二叶(图6-B), 与野生型相比, 除剑叶外, 突变体倒二叶和倒三叶的可溶性蛋白含量分别下降12.26%和16.74%。
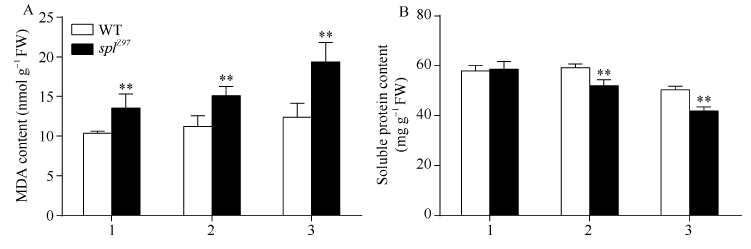 | 图6 孕穗期突变体splZ97及其野生型叶片的MDA和可溶性蛋白含量1: 剑叶; 2: 倒二叶; 3: 倒三叶。* * 在0.01水平上差异显著。Fig. 6 MDA and soluble protein contents of splZ97 and its wild-type (WT) plants at booting stage1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top. * * Significantly different at P< 0.01 (t-test). |
splZ97/珍97、splZ97/02428和splZ97/秀水110三个杂种F1单株的叶片均表现正常, 说明突变体splZ97的斑点叶性状由隐性位点控制。而这3个杂种F1的自交F2后代的田间结果表明, 具正常叶片表型的单株与具斑点叶表型的单株数之比均符合孟德尔3:1的分离比(表2), 证实突变体splZ97的斑点叶性状受一对隐性核基因控制。
| 表2 突变体splZ97的遗传分析 Table 2 Genetic analysis of the mutant splZ97 |
利用splZ97/02428的F2群体中的207株具斑点叶性状的单株作为定位群体。选取均匀分布于水稻12条染色体上的500对SSR标记及50对InDel标记逐条对02428和突变体splZ97进行亲本多态性分析, 利用筛选到的多态性标记分析正常基因池和突变基因池。发现在突变体基因池和正常基因池水稻第12染色体上的SSR标记RM511、RM1246和RM1300 (引物序列见附表1)存在明显偏分离, 利用207个突变体单株验证以上标记, 初步把SPLZ97定位在RM511和RM1300之间(图7)。为进一步定位SPLZ97基因, 利用RM511和RM1300之间的5对在两亲本之间具有多态性的SSR标记(包括RM28204、RM1246、RM28466、RM28485和RM28502)(引物序列见附表1)将该基因定位在RM28466与RM28485之间, 物理距离约189 kb, 横跨AL935072、AL928753、BX072546和AL713908等4个BAC (图7), 其间有EST支持的22个ORF (http://rapdb.dna. affrc.go.jp/viewer/gbrowse/irgsp1/)(表3)。
| 附表1 用于SPLZ97基因定位的SSR标记 Supplementary table 1 SSR markers used for SPLZ97 gene mapping |
| 图7 水稻斑点叶基因SPLZ97在第12染色体上的分子定位Fig. 7 Molecular mapping of SPLZ97 gene on the long arm of chromosome 12 |
| 表3 定位区间内的基因及功能注释 Table 3 Gene names and their annotations in the target interval |
斑点叶突变体最明显的外在表现是叶片上出现坏死斑点或斑块, 而内在表现则为程序性细胞死亡(programmed cell death, PCD), 并由此带来叶绿体降解、蛋白质降解和ROS (reactive oxygen species, ROS)累积等众多生理生化变化[7, 8]。研究表明, 叶绿体降解不仅引起ROS累积, 也介导PCD形成[7], 因此叶绿素含量及叶绿素a/b的比值是衡量细胞死亡的重要生理指标。利用EMS诱变获得斑点叶突变体splZ97, 其斑点叶症状始于分蘖期(图1-A), 孕穗期后除剑叶外所有叶片均不同程度出现褐色斑点(图1-B, C, D, E, F)。叶绿素含量测定表明, 突变体剑叶、倒二叶和倒三叶的叶绿素a/b比值均低于其野生型珍97且依次显著下降, 说明突变体叶肉细胞已发生PCD, 而且其叶绿素a比叶绿素b下降更快, 究其原因可能是倒二叶和倒三叶中的ROS累积, 特别是O2܋的累积显著高于剑叶(图5-A), 导致叶绿素a对活性氧离子的敏感性高于叶绿素b[31]。至于孕穗期突变体光合色素含量明显低于野生型的原因, 可能是突变体基因的突变影响叶绿素的合成[32], 从而造成突变体叶色偏黄(图1-B, C)。
引起叶片斑点叶形成的原因极其复杂, 它是由植物自身的生理生化原因及外界环境因素共同作用决定的。其中, 作为植物体内重要信号分子ROS, 在抵御各种逆境胁迫、调控植物生长发育中起重要作用, 其重要来源是叶绿体, 伴随叶片叶绿体的降解或解体, 叶绿体中的电子传递链受到抑制, 致使1O2、H2O2和O2܋大量形成[33]。研究显示, O2܋是引起拟南芥lsd1突变体类病斑形成的重要因素, 外源SOD能有效抑制该突变体叶片出现类病斑症状[34]。本研究中野生型珍97的剑叶、倒二叶和倒三叶间的ROS(主要包括H2O2和O2܋)无显著性差异, 但突变体的O2܋含量则极显著高于野生型且依次增加(图5-A), 叶片NBT组织化学染色进一步证实突变体的O2܋明显累积(图5-C)。因此推测O2܋是引起splZ97斑点叶形成的重要因素。同时, 由于叶片中大量累积ROS, 导致细胞膜脂质过氧化而产生大量MDA (图5-A), 加剧膜损伤并致使细胞坏死[35]。当然, 在正常植株体内, 存在重要的抗氧化酶系统, 如SOD、POD和CAT, 调控ROS的产生和降解, 维持ROS含量处于一个动态平衡而使细胞免于氧化胁迫伤害[36]。其中, CAT专一作用于H2O2, 将其分解成无毒的H2O和O2。本研究中突变体倒二叶和倒三叶的CAT活极显著低于剑叶, 致使倒三叶细胞中存在H2O2累积(图4-B)。至于突变体叶片, 尤其是倒二叶和倒三叶的POD活性(图4-A)和SOD活性(图4-C)极显著高于野生型且依次显著升高的原因, 可能是斑点叶形成初期细胞自我保护, 诱导SOD和POD合成相关基因的表达而使SOD和POD活性增加, 该结果进一步证实了汪媛[37]和赵晨晨等[38]的研究。
此外, 任何与叶片生理生化代谢相关基因的突变, 均有可能破坏细胞内部的生理生化平衡, 导致ROS累积及内源激素的失衡, 引起细胞程序性死亡并最终造成斑点叶的形成。研究表明, NAC、WRKY和MYB是植物中最重要的三大转录因子, 调控各种分子生理生化过程, 进而影响植物生长发育[39, 40, 41, 42]。过量表达AtMYB30基因, 将导致转基因拟南芥出现类病斑症状, 究其原因, 可能是该基因的过量表达促进了SA合成途径中ICS基因的上调表达, 进而导致SA的累积而造成斑点叶的形成[43, 44]。而编码水稻热激转录因子的SPL7基因的突变将导致突变体spl7从水稻分蘖期到抽穗期, 整个叶片表面出现红棕色小斑点[9]。此外, 在高粱中表达水稻蛋白激酶基因OsCDPK-7, 将导致转基因后代表现类病斑症状且对盐敏感[45]。迄今为止, 在第12染色体长臂上已定位或克隆SPL11[15]和SPL21[46]等斑点叶相关基因, 而这些基因均不在本研究的定位区间, 因此SPLZ97是一个新的斑点叶相关基因。与报道相关的导致植物类病斑形成的基因则仅有蛋白激酶基因(LOC_ Os12g37570)和MYB转录因子(LOC_Os12g37690)。基于突变体的MDA含量、CAT活性、O2-含量及H2O2含量等结果初步认为splZ97基因与ROS密切相关, 而依据突变体对盐敏感的结果则进一步将候选基因指向蛋白激酶基因(LOC_Os12g37570)。当然, SPLZ97基因的最终确定要依赖于候选基因的测序分析及遗传互补验证。
splZ97是一个新的斑点叶且对盐敏感的突变体, 其斑点叶症状始于分蘖期, 此后由叶边缘向叶片内部扩散, 直至遍布整个叶片。与野生型对照相比, 突变体剑叶、倒二叶和倒三叶的叶绿素含量极显著降低, ROS明显累积, MDA含量、SOD和POD活性极显著升高; 而CAT活性显著下降, 清除H2O2的能力也显著下降。splZ97斑点叶性状受1对隐性核基因控制, 位于第12染色体长臂的RM28466和RM28485两个标记的189 kb区间, 本研究结果为进一步克隆SPLZ97基因并揭示斑点叶形成的分子生理机制奠定了基础。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|