

{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻组织特异型人工合成启动子的设计、构建及功能鉴定
[王睿1, 2  , 朱梦琳
, 朱梦琳1 , 高方远2 , 任鄄胜2 , 陆贤军2 , 任光俊2 , 林拥军1, * ]
, 朱梦琳]
|
|


第一作者联系方式: E-mail:wangray1987@163.com
合成启动子是合成生物学的一个重要研究领域及研究热点。水稻是世界上最重要的粮食作物之一, 亦是禾本科作物功能基因组研究的模式植物。本研究旨在对水稻组织特异表达启动子的合成作有益尝试。根据已发表的文献选取了一些与组织特异表达相关的顺式元件, 将它们以不同的方式组合并连接Mini 35S核心启动子以驱动 GUS报告基因的表达。转基因水稻的GUS组织化学染色和酶活测定结果证实, 通过上述方法在水稻中成功构建出组织特异型合成启动子, 同时也揭示了顺式元件在合成启动子中不同的组合方式对启动子的表达活性和表达模式起着关键作用。本研究为植物合成启动子的设计思路和构建方法提供了有益信息和实践基础。
, ZHU Meng-Lin
As an important part of synthetic biology, synthetic promoter has gradually become a hotspot in current biology. Rice is one of the most important food crops in the world and a model plant for functional genomic research in cereals. The purpose of the present study was to design and construct tissue-specific synthetic promoters in rice. We selected several tissue-specific cis-elements based on previous reports and assembled them in different patterns. The combinations were respectively fused to Mini 35S promoter. GUS assays of the transgenic plants indicated that our work successfully created tissue-specific synthetic promoter with the above approach and also revealed that the assembly patterns of cis-elements are critical for the expression patterns and expression efficiencies of synthetic promoters. Our study provides useful information and a practical basis for the design ideas and construction approaches of synthetic promoters in plants.
启动子是调控基因转录的一段DNA序列, 它是基因表达调控中最关键的因子, 能够精确调控目的基因的表达模式及表达丰度[1]。随着研究的深入, 人们在基因组中发现了不同类型的天然启动子, 如组成型启动子、时空特异型启动子、诱导型启动子等[2, 3]。在基因工程研究中, 天然启动子的使用和开发占较大部分。但由于人们对天然启动子中的许多序列认识不足, 增加了其在应用中的不确定性。与天然启动子相比, 人工合成启动子可根据不同目的自由构建, 提高基因表达时期、部位和条件的精确性。随着合成生物学的发展, 人工合成启动子的报道逐渐增多。其中大部分是微生物合成启动子的研究。这些研究采用的主要策略是将大批量不同顺式调控元件或者随机序列与核心启动子融合并转化不同细菌或者真菌, 通过对报告基因表达情况的检测来筛选表达模式和表达丰度与试验目的吻合的启动子[4, 5, 6]。然而, 这种方法并不适合转化周期长、鉴定工作量大的受体, 比如植物。与此同时, 在动物中也报道了人工构建的合成启动子。这些合成启动子的构建方法主要通过不同表达调控序列的同向组装, 这类启动子同时具备期望的表达效率和理想的表达模式[7, 8]。
植物中合成启动子的研究相对较少且大多集中于诱导型合成启动子, 主要利用顺式调控元件融合核心启动子的方法构建。Rushton等[9]曾利用W1-box、W2-box、GCC元件、JERE元件等组合成多种四聚体人工合成启动子。不同病原菌处理下的转基因植株GUS分析结果显示, 不同启动子的诱导因子、本底表达水平和诱导程度均存在明显差别。通过四拷贝小麦来源的脱落酸应答元件(4× ABRE)或者两拷贝脱落酸应答元件连接两拷贝大麦HVA22结合元件(2× ABRC)分别与Mini 35S融合发现, 合成启动子能够诱导GUS报告基因在转基因烟草中对高盐、脱水和脱落酸应答表达, 证明它们能够作为一种有效的胁迫诱导启动子[10]。Koschmann等[11]根据PathoPlant数据库中的拟南芥芯片数据, 挑选出受病原菌诱导上调表达的基因, 利用BEST软件寻找这些基因启动子区的保守序列, 然后与AthaMap、PLACE和AGRIS数据库中的顺式调控元件比对, 挑选与已知顺式调控元件的相似度低的保守序列, 通过合成启动子的方法验证。Liu等[12]将一些与病原菌和植物防御信号分子诱导相关的顺式调控元件与核心启动子融合并稳定转化烟草和拟南芥, 对转基因植株进行病原菌、水杨酸、乙烯和茉莉酸甲酯处理, 结果证实合成的诱导型启动子在转基因烟草和拟南芥中能够发挥预期功能。次年, 该团队根据大豆基因芯片数据, 挑选出受大豆胞囊线虫诱导的基因, 综合利用7种生物信息学工具寻找这些基因启动子中可能与大豆胞囊线虫诱导相关的顺式调控元件, 并通过合成启动子的方法在转基因大豆中对候选的顺式元件验证[13]。植物中合成启动子的研究主要集中于诱导型合成启动子而很少涉及组织特异型合成启动子。一个重要原因就是通过单一顺式元件连接核心启动子的方法无法兼顾促进目标组织表达同时抑制非目标组织表达这两个需求, 而多种顺式元件在合成启动子设计中的组合方式和拷贝数等研究报道非常少。因此, 在植物中构建组织特异型合成启动子的难度相对较大。
水稻是世界上最重要的粮食作物之一, 亦是禾本科作物功能基因组学研究的模式植物。完备的基因组信息[14, 15, 16]及较为清楚的基因表达信息[17]为组织特异型启动子的研究提供了极大的便利。前人已经克隆了一些与水稻组织特异表达相关的顺式调控元件[1, 18, 19], 这些研究为水稻组织特异型合成启动子的研究奠定了坚实的基础。本研究将一些与组织特异表达相关的顺式元件以不同的方式组合用于构建合成启动子并通过转基因水稻对这些合成启动子进行功能鉴定。该结果揭示了顺式元件在合成启动子中不同的组合方式对启动子的表达活性和表达模式起着关键作用。本研究为探索多种顺式元件在合成启动子设计中的组合方式和拷贝数等问题提供了有价值的信息。
载体构建和遗传转化所涉及的菌株包括大肠杆菌TOP10和农杆菌EHA105, 它们均由本实验室长期保存。遗传转化所涉及的水稻品种为粳稻品种中花11 (Oryza sativa L. ssp. japonica), 由本实验室保存。
将一些已鉴定与组织特异表达相关的顺式调控元件以不同方式组合并连接Mini 35S核心启动子(详见结果与分析2.1), 并将设计好的合成启动子送交金斯瑞公司合成。获得上述合成启动子后, 分别将它们通过Hind III和BamH I内切酶位点连入启动子功能分析载体pDX2181[18]。将测序验证正确的重组质粒通过电转化法导入农杆菌菌株EHA105, 用于后续农杆菌介导的水稻转化试验。
水稻愈伤组织的培养和农杆菌介导的遗传转化参照Hiei等的方法[20]。将成熟的水稻种子去壳, 以75%酒精表面消毒2 min, 0.15%升汞深度消毒15~ 20 min, 灭菌蒸馏水洗5~6次; 置装有诱导培养基的小三角瓶中, 暗培养室诱导40 d左右, 长出胚性愈伤组织后接入继代培养基, 暗培养2代, 每次15 d左右; 挑选状态良好的愈伤组织进行农杆菌浸染; 19~21℃共培养2 d; 用灭菌的蒸馏水洗涤愈伤组织直至液体澄清; 将其转至含有50 mg L-1潮霉素的筛选培养基上, 筛选2代, 每次15 d左右; 挑取抗性愈伤组织转入分化培养基, 光照培养30~40 d得到分化苗; 炼苗后, 将绿苗移栽至大田。
取转基因植株幼苗叶片, 利用小量抽提DNA法抽提其基因组DNA, 以其作为PCR模板用于转化株的阳性检测, 设计引物GUS-F: 5° -GGGCGAACAGT TCCTGATTA-3° 和GUS-R: 5° -AACGTATCCACGCC GTATTC-3° , 扩增GUS报告基因内部一段699 bp的片段。PCR体系(20 µ L)含: 模板(50 ng µ L-1) 1 µ L、10 × PCR buffer 2 µ L、dNTP (2 mmol L-1) 2 µ L、GUS-F (10 µ mol L-1) 0.2 µ L、GUS-R (10 µ mol L-1) 0.2 µ L、r-Taq 0.2 µ L、ddH2O 14.4 µ L。PCR程序为: 94℃ 5 min; 94℃ 50 s, 60℃ 50 s, 72℃ 50 s, 30个循环; 72℃ 7 min。
转基因植株的GUS组织化学染色参照Jefferson等的方法[21]。将阳性转化植株的各个组织(叶片、叶鞘、茎秆、根、穗和种子)浸入GUS染液(50 mmol L-1 pH 7.0的磷酸钾缓冲液、10 mmol L-1 Na2-EDTA、0.1% Triton X-100、1 mg mL-1 X-Gluc、100 µ g mL-1氯霉素、1 mmol L-1的铁氰化钾、1 mmol L-1的亚铁氰化钾以及20%的甲醇)中, 37℃温育2~10 h后用70%乙醇脱色, 然后在解剖显微镜(Leica MZFLIII)下照相。
选取4个与组织特异表达相关的顺式调控元件, 即GEATFLK[19]、LPSE1、LSE1和LPRSE1[1](表1)用于合成启动子的构建。为构建种子不表达的组织特异型合成启动子, 使用抑制启动子在种子中表达的顺式元件LPRSE1。将以上这些顺式调控元件以5种不同方式组合并连接Mini 35S核心启动子, 得到5个新的合成启动子(图1)并分别构建到启动子功能分析载体pDX2181以驱动GUS报告基因的表达。
| 表1 本研究中用到的顺式元件的信息 Table 1 Information of cis-elements used in this study |
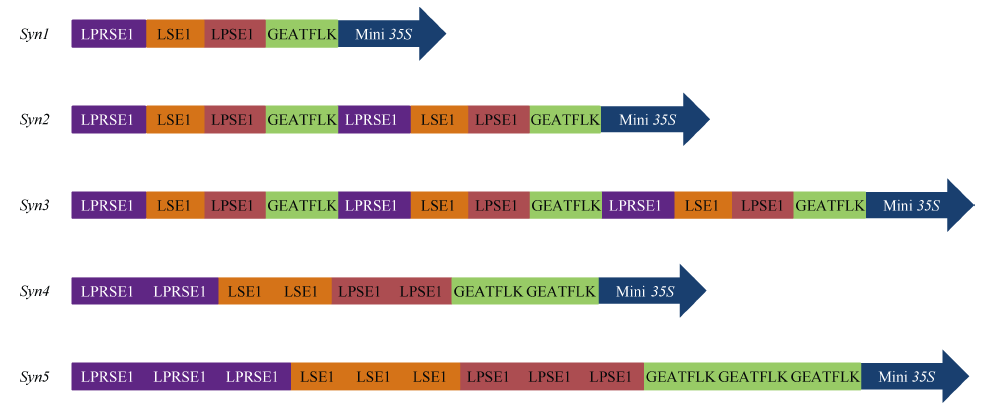 | 图1 不同顺式调控元件组装而成的合成启动子结构示意图Fig. 1 Scheme of synthetic promoters assembled with different cis-regulatory elements |
利用农杆菌介导的遗传转化方法获得不同合成启动子驱动GUS报告基因的转基因植株, 并通过GUS组织化学染色分析不同合成启动子在水稻中的表达模式(图2)。染色的结果显示, 启动子Syn2、Syn3和Syn4与对照Mini35S相比, 叶片中GUS表达出现明显升高; 与之相似, 启动子Syn2、Syn3和Syn4在叶鞘和穗中GUS表达同样得到了提升, 但启动子Syn5驱动GUS在叶鞘和穗中的表达却低于Mini35S; 茎秆染色结果显示, 除Syn3与对照相比表达活性有明显升高外, 其余合成启动子的转基因植株在茎秆中几乎无法观察到蓝色。转基因植株种子的染色结果表明, 除Syn3之外, 其余合成启动子与对照相比种子中的表达活性都得到了不同程度的抑制, Syn5在种子中的表达活性完全丧失。
 | 图2 组织化学染色分析不同合成启动子在水稻中的表达模式Fig. 2 Histochemical analysis of transgenic rice plants containing different synthetic promoters/GUS fusions |
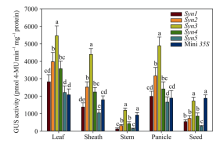
对不同合成启动子驱动GUS基因的转基因植株进行GUS酶活测定。各组织GUS活性定量检测结果(图3)与组织化学染色结果一致, 并更精确地反映出不同合成启动子在不同组织中的表达活性, 也为研究顺式元件的不同组合方式对启动子表达活性和表达模式的影响提供更加可靠的证据。启动子Syn2与Mini35S相比, 叶片、叶鞘和穗中的GUS活性显著升高, 而种子和茎秆中的GUS活性显著降低, 该结果与顺式元件GEATFLK、LPSE1、LSE1和LPRSE1的预期功能一致; 启动子Syn3在5个组织中的表达活性均为所有合成启动子中最高, 在叶片、叶鞘和穗这3个绿色组织中的表达活性与对照相比显著升高, 但其在种子中的表达活性与对照相比不存在显著差异, 该结果表明顺式元件LPRSE1在启动子Syn3中并没有发挥预期功能; 启动子Syn5虽然在5个组织中的表达活性均为最低, 但其在种子中的表达活性完全丧失, 该结果证明本研究通过多种顺式元件组合、连接核心启动子的方式成功构建出种子不表达的组织特异型合成启动子。上述结果表明, 顺式调控元件组合方式的变化对启动子的表达活性具有显著的影响, 进而改变启动子的表达模式。
 | 图3 不同合成启动子驱动GUS报告基因的转基因水稻各个组织的GUS活性定量检测Fig. 3 Quantitative analysis of GUS activity in various tissues of the transgenic plants containing different synthetic promoters/GUS fusions 柱上标以不同字母表示不同转基因植株之间以及转基因植株与对照之间存在显著差异(P< 0.05)。 Bars denoted by different letters are significantly different between different transgenic plants and also between the transgenic plants and the control at P< 0.05. |
在植物合成启动子的研究中, 组织特异型合成启动子一直以来鲜有报道, 而利用顺式调控元件连接核心启动子的方式构建组织特异型合成启动子从未有相关报道。主要原因有二: 一是通过单一顺式元件连接核心启动子的方法无法兼顾促进目标组织表达同时抑制非目标组织表达这两个需求, 而多种顺式元件在合成启动子设计中的组合方式和拷贝数等研究报道非常少, 造成组织特异型合成启动子研究的理论基础薄弱; 二是目前已鉴定且功能清晰的组织特异表达相关顺式元件数量相对较少, 导致组织特异型合成启动子研究的候选资源稀缺。以上两点一直是组织特异型合成启动子构建中的障碍和瓶颈。本研究成功构建出种子不表达的组织特异型合成启动子, 证实了利用顺式调控元件连接核心启动子的方式构建组织特异型合成启动子的可行性, 同时也为探索多种顺式元件在合成启动子设计中的组合方式和拷贝数等问题提供了有价值的信息。
本研究鉴定的5个合成启动子在水稻不同组织中的表达程度不尽相同, 因此可利用它们驱动不同靶基因的表达以实现不同的目标。Syn3在叶片、叶鞘、穗和茎秆这4个绿色组织中的表达强度较高, 可用于光合作用[24]、叶片衰老[25]、株高[26]等相关基因的功能研究。Syn5为种子不表达的组织特异型合成启动子, 利用它驱动抗病虫害基因[27, 28, 29]的表达可在特定的绿色组织中赋予水稻对病虫害的抗性, 同时避免外源蛋白在种子中的产生。
5个合成启动子中, Syn1是4个单拷贝顺式调控元件串联并连接Mini 35S核心启动子而成, Syn2/Syn3是4个单拷贝顺式调控元件串联后以组合的形式重复二/三次并连接Mini 35S核心启动子而成, Syn4/Syn5是4个顺式调控元件以二/三拷贝的形式组合并连接Mini 35S核心启动子而成。分析发现, Syn3在5个组织中的表达活性均为所有合成启动子中最高, 而Syn5则在5个组织中的表达活性均为最低, 且在种子中的表达活性完全丧失。据此推测, 单拷贝顺式元件串联后以组合的形式重复多次的设计方式有利于上调启动子的表达活性, 即有利于正调控顺式元件发挥功能; 而顺式元件以多拷贝形式组合的设计方式有利于下调启动子的表达活性, 即有利于负调控顺式元件发挥功能(图4)。这一结果也证明了顺式元件发挥功能不仅来自自身特性, 而且取决于顺式元件之间的位置关系和相互作用。本研究成功构建出种子不表达的组织特异型合成启动子, 也为植物合成启动子的设计思路和构建方法提供了有益信息和实践基础。
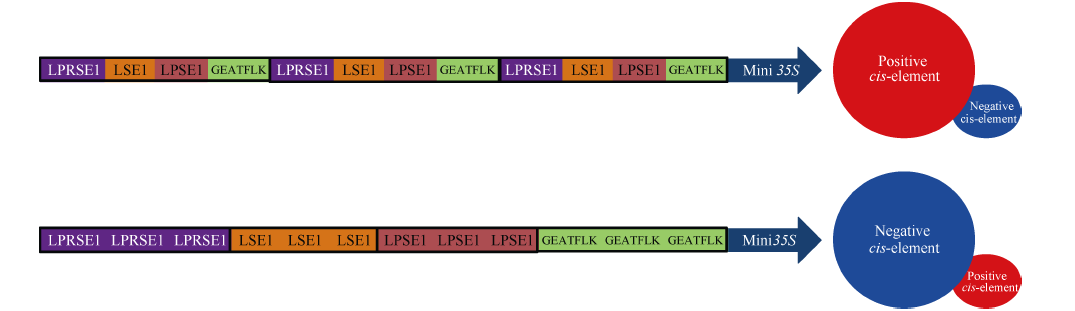 | 图4 多种顺式元件的不同组合方式对合成启动子表达活性影响的推测Fig. 4 Speculation of the influence of different assembly patterns of multiple cis-elements on the expression efficiencies of synthetic promoters |
利用顺式调控元件连接核心启动子的方式构建出种子不表达的组织特异型合成启动子, 并推测出合成启动子中顺式调控元件的组合规律: 单拷贝顺式元件串联后以组合的形式重复多次的设计方式有利于正调控顺式元件发挥功能; 而顺式元件以多拷贝形式组合的设计方式则有利于负调控顺式元件发挥功能。该结果表明, 控制启动子的表达活性和表达模式的因素不仅来自于顺式调控元件自身的功能, 更重要的是顺式元件之间的位置关系和相互作用。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|