



{kind=link}
{kind=link}
水稻重测序核心种质资源的稻瘟病抗性鉴定与评价
[李旭升1, * , 1, *, * , 向小娇2, *, * , 申聪聪2, * , 杨隆维1, *  , 陈凯
, 陈凯2, 3 , 王小文1 , 邱先进1 , 朱小源4 , 邢丹英1 , 徐建龙2, 3, * ]
, 陈凯]
|
|


稻瘟病一直是制约水稻产量的重要因素之一, 稻瘟病抗源筛选是抗性基因挖掘和抗病育种的基础。本试验利用3000份(简称3K)重测序中的1217份核心种质资源, 通过湖北恩施两河和芭蕉2个病圃自然诱发鉴定抗性, 结合不发病条件下农艺性状考察和抗病资源的苗期人工接种抗谱测定, 综合评价和筛选优异的稻瘟病抗源。自然诱发鉴定结果显示材料间的稻瘟病抗感差异显著, 从中共获得144份抗苗瘟、叶瘟和穗瘟的抗病种质。选用稻瘟病综合抗性较好的34份材料以30个不同来源的稻瘟病菌株苗期接种, 鉴定显示有17份材料的抗性频率≥70%, 抗谱较广。农艺性状考察结果显示, 大部分抗病材料植株偏高, 单株产量低, 农艺性状差。结合病圃鉴定、人工接种鉴定和农艺性状考察, 鉴定出7份稻瘟病抗性强、抗谱广且农艺性状较好的优异抗源材料IRGA 411-1-6-1F-A、YJ30、金早47、泉珍10号、YN 1353-3、云粳23和IRAT1047, 可作为抗源亲本用于稻瘟病抗性基因挖掘和品种抗稻瘟性改良。
, CHEN Kai
Rice blast is one of the key factors that restrict rice yield, and screening blast resistance resources is a basis work for mining blast resistance genes and breeding resistant varieties. A set of 1217 accessions selected from 3000 (3K) re-sequenced rice core collection were comprehensively evaluated for blast resistance in two natural disease nurseries at Lianghe and Bajiao in Enshi area and agronomic traits in disease-free field as well as resistance spectrum of resistant accessions at seedling stage. There were significant differences in response of different accessions to rice blast. Among them, a total of 144 accessions with blast resistance at different growing stages were obtained. Thirty-four accessions with relatively high comprehensive blast resistance were inoculated with 30 diverse isolates, showing that 17 accessions with a broad resistance spectrum (≥70%). According to the investigation of agronomic traits, most resistant accessions had high plant height, low grain yield per plant and poor agronomic traits. Finally, seven accessions with high blast resistance, broad resistant spectrum and relatively good agronomic traits were selected, including IRGA411-1-6-1F-A, YJ 30, Jinzao 47, Quanzhen 10, YN 1353-3, Yunjing 23, and IRAT1047, were identified. Those resistant accessions can be used in mining resistance gene and variety improvement for blast resistance.
由子囊菌Magnaporthe oryzae引起的稻瘟病是最具毁灭性的一种水稻病害, 也是影响水稻稳产、高产的主要障碍因素[1, 2]。一般年份引起水稻减产10%~20%, 严重的减产40%~50%, 甚至绝收[3]。实践证明, 培育和推广抗稻瘟病品种是控制稻瘟病最经济、有效和环保的措施[4, 5]。但由于稻瘟病菌生理小种数量众多而且极易变异, 以致一个抗病品种在生产上使用3~5年就丧失抗性[6, 7, 8]。因此不断挖掘新的优异抗源和抗病基因, 培育和推广抗病品种, 是提高品种抗性水平的有效手段[9]。
植物种质资源蕴藏着植物新品种选育赖以生存的各种有利变异[10]。截至2015年3月, 已从水稻种质资源中定位了至少84个稻瘟病抗性主效基因, 分布在69个位点上, 已有24个基因被成功克隆[11], 其中Pigm、Pi1和Pi2属广谱抗性基因, 被广泛利用于抗病育种[12]。水稻品种的抗瘟性鉴定和评价是抗源挖掘和利用的重要基础工作[13], 其中自然诱发鉴定成本低、操作简单, 是一种较经济实用和符合生产实际的抗瘟性鉴定技术[14]。湖北省恩施两河病圃是恩施州农业科学院20世纪80年代初创建的, 经30余年的长期培育, 其稻瘟病菌的菌源丰富, 生理小种分布广泛, 小种组成复杂, 致毒力强的小种多[15, 16], 已成为全国24个二级病圃中发病最重的病圃之一。恩施芭蕉病圃发病条件虽不如两河病圃好, 但海拔较低, 有利部分熟期偏迟材料的抗性鉴定, 有较好的辅助作用。
目前, 3000份(简称3K)全球水稻核心种质重测序项目已经完成[17]。该项目产生的序列数据、SNP数据和InDel数据为水稻重要性状的遗传研究提供了基础[18]。3K水稻来自全球89个国家, 类型多样, 遗传多样性丰富, 是筛选稻瘟病抗源和挖掘优异抗瘟性基因的理想群体。本研究在湖北省恩施州两河和芭蕉病圃的自然诱发条件下对部分3K水稻核心种质资源进行苗瘟、叶瘟和穗瘟的自然诱发抗性鉴定, 结合苗期人工接种抗谱测定和农艺性状考察, 旨在筛选主要农艺性状较好的稻瘟病优异抗源, 为抗稻瘟病基因发掘和抗稻瘟病育种提供基础材料。
根据3K种质资源在海南种植的生育期和株高表现, 选取抽穗期在125 d以内且株高较适中的1217份材料用于稻瘟病抗性鉴定与分析。田间自然诱发地点在稻瘟病自然重病区恩施土家族苗族自治州, 设计的两个病圃分别在恩施市的两河(30° 08′ N, 109° 13′ E, 海拔1005 m)和芭蕉(30° 09′ N, 109° 25′ E, 海拔440 m)。于湖北荆州长江大学基地种植农艺性状考察材料, 由广东省农业科学院植物保护研究所完成人工接种抗性评价。
2015年4月15日将3K种质资源中筛选出的1217份材料在恩施市芭蕉乡播种育苗, 分2个区组。以壮苗标准管理苗床, 且只防虫不防病。播种30 d后, 将材料秧苗移栽到芭蕉和两河病圃中。每份材料设置2次重复, 每重复种2行, 每行10棵, 组成一个小区。每隔19个小区插入1个感病对照小区, 2个垄小区之间种植混合诱发苗, 合并为1厢, 厢的四周种植1株混合诱发苗, 田四周种植3~4行混合诱发苗, 以创造均等的发病条件。由湖北省宜昌市农业科学院水稻研究所提供稻瘟病诱发和指示材料, 包括生育期不同的品种丰两优4号、文胜糯、CO39、广陆矮4号和丰两优香1号, 混合种植诱发稻瘟病。早稻以文胜糯和CO39为感病指示对照; 早熟中稻以广陆矮4号和丰两优香1号为指示对照; 迟熟中稻以丰两优4号为指示对照。按丰产标准管理稻田, 并尽量创造稻瘟病发病条件。
在秧苗移栽前3 d左右调查苗瘟, 在水稻分蘖盛期至拔节期间的病害流行高峰期调查叶瘟, 在80%稻穗尖端5~10粒稻谷进入黄熟期时调查穗瘟, 均以感病对照品种发病达到稳定为标准确定调查日期。穗瘟调查中每份材料每个重复随机调查100穗。参照《农作物品种区域试验抗病性鉴定操作规程》(DB42/T208-2001)[19]的标准评定各阶段病情。苗、叶瘟病级分0、1、2、3、4、5、6、7、8、9共10级, 0~3级为抗, 4~9级为感。穗瘟病级和综合病级分别根据穗瘟发病率和综合指数划分为0、l、3、5、7、9共6级, 0~1级为抗, 3级穗瘟为中抗, 综合病级为感, 5~9级为感。穗瘟发病率(%)=发病总穗数/调查总穗数× 100, 综合指数=叶瘟病级× 0.25+穗瘟发病率计算值× 0.25+穗瘟损失率计算值× 0.5。
将通过田间诱发鉴定得到的34份稻瘟病抗性材料, 于30 cm × 20 cm × 5 cm的搪瓷盆育苗, 将待鉴定种子和对照品种Tetep (抗病对照)及丽江新团黑谷(感病对照)的种子消毒并催芽后挑选饱满露白的分区均匀播于盆中, 每个材料10粒种子, 2次重复。水稻苗长至3.5~4叶龄时进行人工喷雾接种。稻瘟病菌分生孢子悬浮液浓度大约为1× 105个 mL-1, 每盆20 mL的喷雾量。将接种后的水稻苗在24℃的接菌室中保湿培养48 h后移至温室, 25~28℃下保湿培植, 7~10 d当感病对照发病情况达到稳定后, 调查各个材料的发病情况。参照国际水稻研究所的稻瘟病抗性分级标准[24], 对每株材料的病情分级, 0~3级为抗(R), 4~9级为感(S)。记载每个材料对每个菌株的抗性反应时, 去掉病情重的那个重复, 另一个重复中感病植株< 3株, 则定为抗病, 否则定为感病。
抗性频率(%)=(接种后表现抗的菌株数/接种总菌株数)× 100。抗性频率(%)≥ 90为高抗(HR), 80≤ 抗性频率(%)< 90为抗(R), 70≤ 抗性频率(%)< 80为中抗(MR), 60≤ 抗性频率(%)< 70为中感(MS), 50≤ 抗性频率(%)< 60为感(S), 抗性频率(%)< 50为高感(HS)。
同年夏季于湖北荆州种植1217份种质资源, 5月5日播种, 6月3日移栽。每份材料种植5行, 每行10株, 株行距20.0 cm × 20.0 cm, 2次重复。从每重复随机取中间5株调查田间生育期、株高、剑叶长宽和单株产量, 从每株取2个主穗, 考察穗长、一次枝梗数、每穗实粒数、每穗颖花数、结实率和千粒重。
利用Microsoft Excel 2007整理数据, 并计算平均值、标准差、变异系数和变幅。利用SPSS 21.0软件进行病圃、材料和重复的方差分析和两病圃各抗病指标的相关性分析。对于方差分析使用一般线性模型和类型1的平方和模型; 相关性分析前先对数据进行z标准化, 标准化后群体的平均值为0, 标准差为1。
根据重测序种质资源的SNP聚类, 3K种质资源被划分成12个组, 包括5个籼稻组、4个粳稻组和Aus、Aro和Adm 3个组(图1)。参试的1217份资源中, 完成测序的材料有1159份, 另58份材料没有测序结果。1159份(黄色圆点表示)材料在3K种质资源12个组中都有分布, 表明测试材料的遗传多样性丰富。
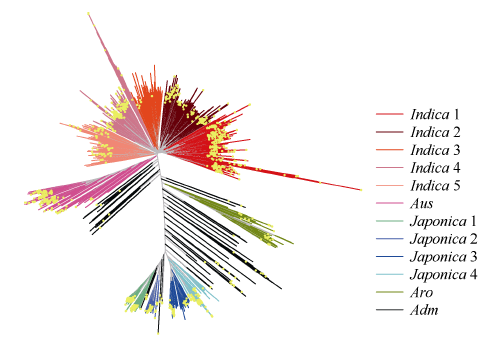 | 图1 参试材料(黄色圆点表示)在3K重测序种质资源分类图中的分布Fig. 1 Distribution of the tested materials with yellow dots in the catalog of 3K re-sequenced germplasm |
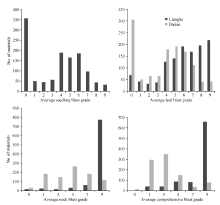
1217份种质资源种植在恩施两个病圃中, 914份生育期较一致, 146份迟熟, 37份特迟熟, 另外120份不抽穗。对于苗叶瘟采用所有材料的病情数据, 而穗瘟及综合病情评定选用914份生育期较一致材料的病情数据。两个病圃中不同时期各材料的病情分布如图2和表1所示, 各材料的苗叶瘟病级和穗瘟病级呈离散分布, 表明这些材料对稻瘟病抗性的多样性丰富, 适用于稻瘟病抗源的筛选。
不同病圃、不同时期各材料稻瘟病病级差异极显著(P < 0.0001), 变异系数在23.71%~80.26%之间, 表明各材料对稻瘟病的抗感差异显著。2个病圃间各时期病情差异极显著(P < 0.0001), 两河病圃中材料不同时期病级的平均值(5.83~8.28)均大于芭蕉病圃中材料的病级平均值(3.18~4.46), 表明两河病圃的病情比芭蕉病圃更为严重。芭蕉病圃中材料各时期病情的变异系数(60.09%~80.26%), 均大于两河病圃中材料的病情变异系数(23.71%~42.90%), 表明各材料芭蕉病圃的抗感差异比两河病圃的更为显著。同一病圃中不同时期病情在重复间差异不显著(P > 0.05), 说明除了品种以外的其他条件一致性较好, 鉴定结果比较可靠(表1和表2)。
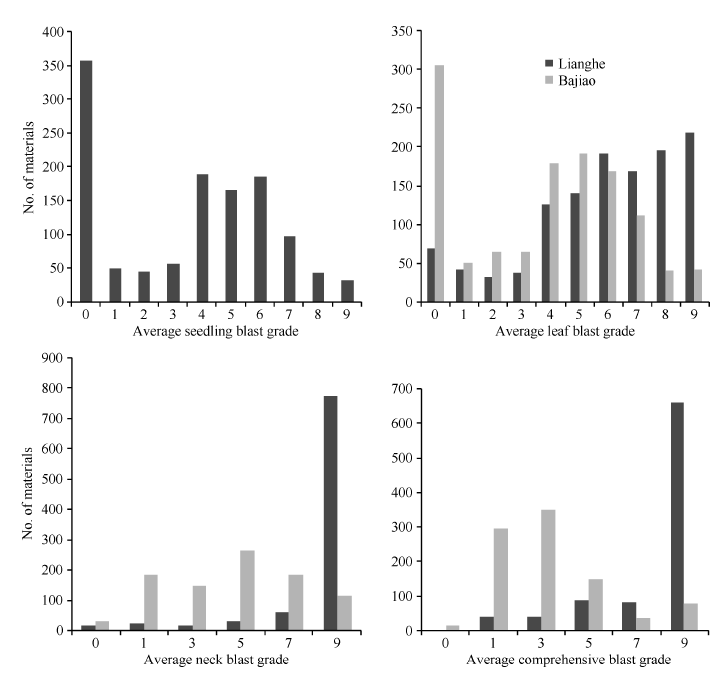 | 图2 参试材料在两河和芭蕉病圃中的稻瘟病病情分布Fig. 2 Distribution of blast incidence in the nurseries of Lianghe and Bajiao |
| 表1 参试种质资源在两个病圃中的稻瘟病病情表现 Table 1 Performance of blast incidence of the tested germplasm in the nurseries |
| 表2 稻瘟病病情的方差分析 Table 2 Variance analysis of blast incidence |
瘟同一病圃不同时期病情数据和不同病圃同一时期病情数据的相关性分析表明, 芭蕉病圃苗瘟与叶瘟的相关系数为0.952, 呈极显著正相关, 苗瘟与穗瘟的相关系数为0.410, 叶瘟与穗瘟的相关系数为0.414, 均呈极显著中度正相关, 表明苗瘟抗性与叶抗性相关性较高, 而苗瘟、叶瘟与穗瘟抗性相关性较低。两河病圃和芭蕉病圃材料之间的叶瘟、穗瘟和综合病级间相关系数分别为0.569、0.362和0.435, 均呈极显著中度正相关, 表明2个病圃中材料的发病情况具有较大程度的一致性。
综合两个病圃中材料的病情数据, 根据不同时期的最高病级筛选不同时期的稻瘟病抗性材料。1217份种质资源中, 抗苗瘟和叶瘟的材料有129份, 占10.59%。914份正常抽穗种质中, 最高穗瘟率病级小于1级的材料有29份, 占3.17%, 苗叶瘟和穗瘟3个时期抗性均强的材料有19份, 占正常抽穗材料的2.08% (表3)。
| 表3 不同时期稻瘟病抗性材料统计 Table 3 Statistics of blast resistant materials at different stages |
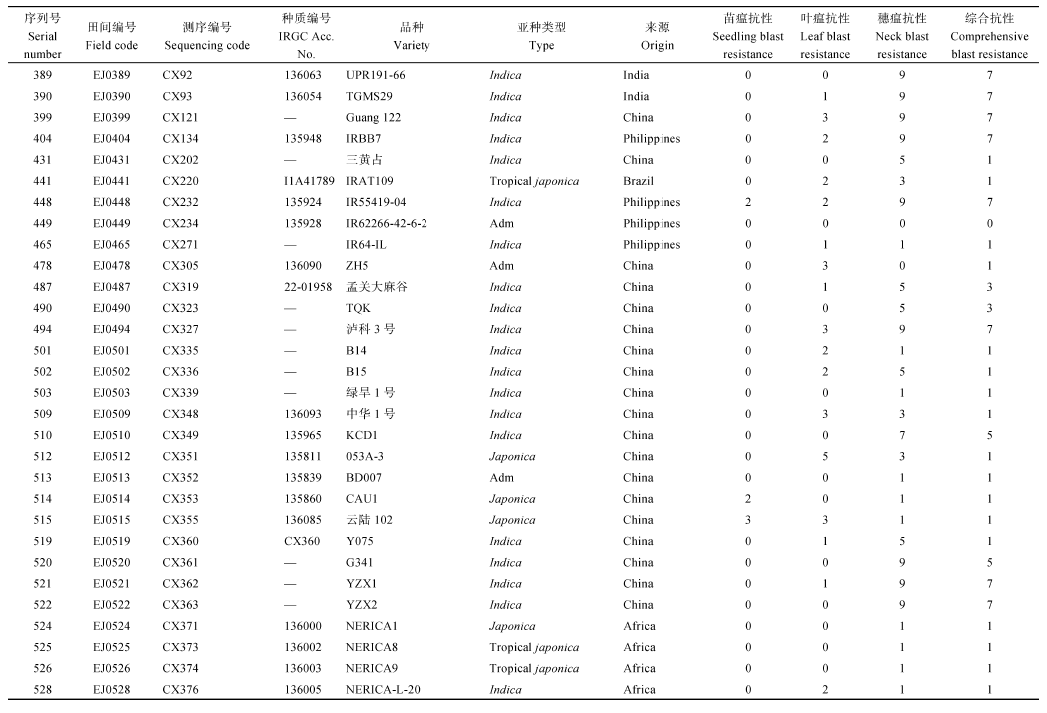
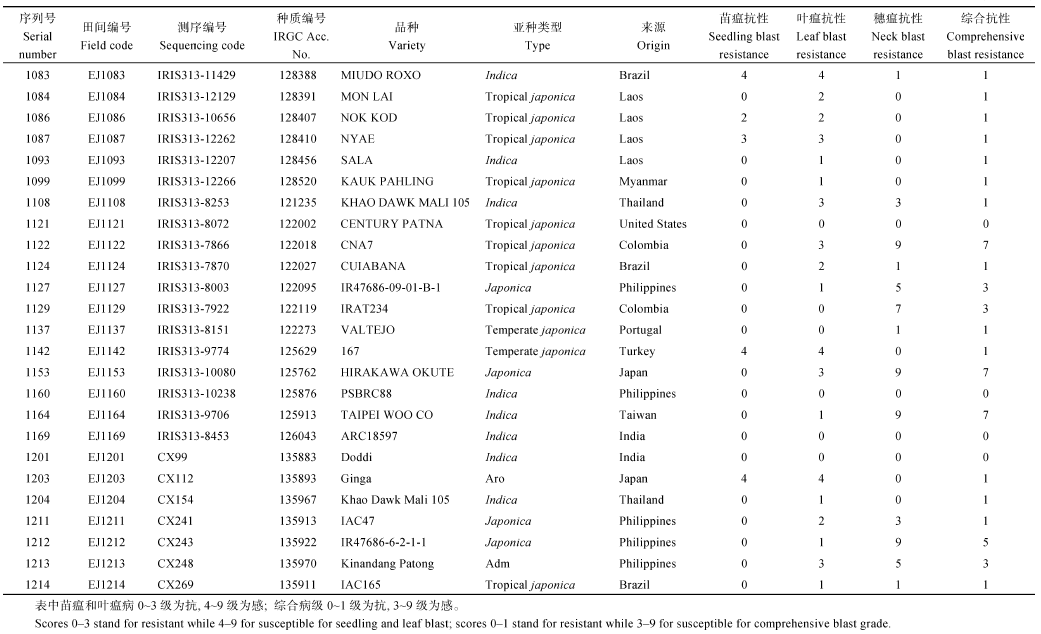
总共获得的144份抗不同时期稻瘟病的抗病材料中(见附表1), 籼稻82份, 粳稻15份, 温带粳稻11份, 热带粳稻23份, Adm 6份, Aus/boro 5份, Aro和中间类型各1份。其中一半以上的抗性材料属于籼稻, 占56.9%, 其次是热带粳稻, 占15.9%。这些材料中, 37份来自中国, 20份来自菲律宾, 12份来自印度, 7份来自老挝, 来自哥伦比亚、印度尼西亚、巴西、越南的各5份, 来自美国、阿根廷、非洲的各4份, 来自孟加拉国、马来西亚、意大利的各3份, 剩余的27份材料分别来自缅甸、柬埔寨、古巴、罗马尼亚、巴基斯坦、法国、日本等20个国家。
从这些材料中筛选了34份稻瘟病综合抗性较好(最高苗瘟和叶瘟均≤ 5级, 最高穗瘟发病率≤ 10%, 最高综合指数≤ 2.5, 最高综合抗级≤ 3)且病圃田间目测综合表现较好的材料(表4)。其中, 籼稻13份(4份来自中国), 粳稻、温带粳稻和热带粳稻共计18份, Adm 2份, Aro 1份。不同种子来源(EJ0885和EJ0967)的IRAT 104在病圃的抗性表现略有细微差异。
| 表4 病圃综合抗性强和田间表现较好材料的稻瘟病病情 Table 4 Blast incidence of materials with good blast resistance and field performance in the nursery |
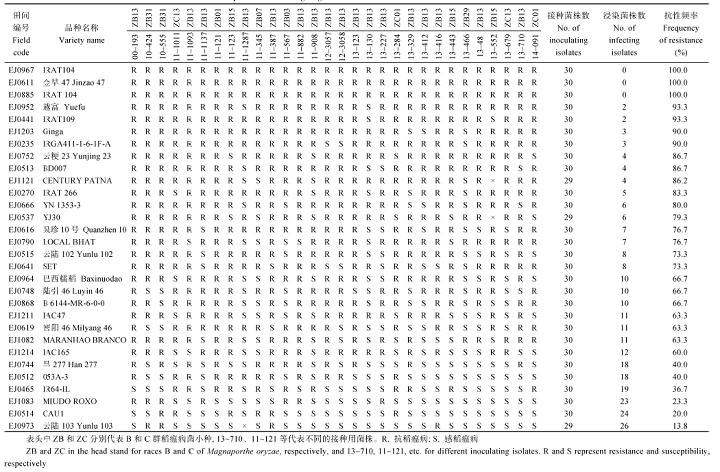
34份材料中, EJ0546和EJ1142因缺种未接种鉴定, EJ0876和EJ1137因发芽率低未能测定其抗谱。剩余30份材料接种30个籼稻小种后的抗性反应如表5所示, 抗性频率≥ 70%的材料共17份, 占接种材料的56.67%, 达到中抗以上水平, 抗谱较广。其 中, 7份材料EJ0967、EJ0611、EJ0885、EJ0952、EJ0441、EJ1203和EJ0235的抗性频率≥ 90%, 达到高抗(HR)水平, 抗谱广; 5份材料EJ0752、EJ0513、EJ1121、EJ0270和EJ0666的抗性频率在80%~90%之间, 达到抗(R)水平; 5份材料EJ0537、EJ0616、EJ0790、EJ0515和EJ0641的抗性频率在70%~80%之间, 达到中抗(MR)水平。
| 表5 从2个病圃筛选获得的30份抗性材料的苗期抗谱测定 Table 5 Test of resistance spectrum at seedling stage for 30 accessions with llast resistance screened from the two nurseries |
除未正常抽穗的18份材料和EJ1203未获得数据外, 144份抗不同时期稻瘟病的材料的抽穗期、穗长和结实率变幅较小, 其余性状变幅均较大, 变异系数> 15% (表6)。这些材料的单株产量均值为27.1 g, 变幅为8.1~65.4 g, 株高均值为115.6 cm, 变幅为64.9~172.0 cm, 表明多数材料产量较低、植株较高, 农艺性状较差。
综合病圃自然诱发鉴定、苗期抗谱测定和农艺性状表现, 以综合病级≤ 1、抗谱≥ 70%、单株产量≥ 抗病资源单株产量均值27.1 g、株高≤ 抗病资源株高均值115.6 cm为标准, 最后获得7份优异的稻瘟病抗源材料, 其主要农艺性状、最高综合病级和抗谱如表7所示。其中, 金早47和泉珍10号是来自中国的籼稻, IRGA 411-1-6-1F-A是来自哥伦比亚的籼稻, YJ30和云粳23是来自中国的温带粳稻, YN1353-3和IRAT 104分别是来自缅甸的粳稻和法国的热带粳稻。
| 表6 144份抗病资源的农艺性状 Table 6 Agronomic traits of 144 blast resistant germplasm |
| 表7 7份优异抗源的农艺性状和稻瘟病抗性 Table 7 Agronomic traits and blast resistance of seven excellent resistant resources |
本试验1217份种质材料分为籼稻、粳稻、Aus、Aro和Adm五大类共12个组, 遗传多样性丰富, 生育期和株高差异很大。为了准确评价抗瘟性并尽可能鉴定出不同时期抗病的抗源材料, 我们选择了恩施市的两河和芭蕉2个自然诱发点。两河点海拔1005 m, 在水稻生长期间阴雨、寡照天气居多、气温适宜, 非常利于发病, 常年鉴定结果偏重, 是国家二级稻瘟病鉴定圃。芭蕉点海拔440 m, 发病条件不及两河点, 但由于相对充足的光温条件, 能保证有更多的材料正常抽穗成熟。从实际鉴定结果来看, 1217份材料在2个病圃中, 914份生育期较一致, 能正常成熟; 303份迟熟、特迟熟和不抽穗的材料大多发生在两河点。由于水稻抽穗期对苗、叶瘟没有影响而对穗瘟影响大, 因此试验中调查了全部供试材料的苗、叶瘟病情, 仅调查了914份正常抽穗材料的穗瘟病情。2个点不同时期的稻瘟病病级分布存在差异, 尤其是穗瘟级别和综合病级两河点明显重于芭蕉点, 呈现发病偏重的态势。芭蕉病圃中材料各时期病情的变异系数均大于两河病圃, 表明芭蕉病圃材料间的抗性差异更明显, 其数据更有利于抗瘟性基因的定位和挖掘。两河病圃中材料各时期病级平均值均高于芭蕉病圃, 表明两河病圃材料病情更严重, 更有利于筛选强抗稻瘟病的抗源, 上述结论与以往研究结果一致[16]。
尽管供试材料在2个病圃间病情差异显著, 但叶瘟、穗瘟及综合病级在2个病圃间都呈极显著中度正相关, 表明材料在两个病圃中的病情趋势具较大程度的一致性。两个病圃间的病情差异可能是由两个病圃的病菌生理小种种类差异或气候差异引起的。因此, 对于遗传差异大、类型丰富的种质资源材料的稻瘟病初步鉴定, 利用不同的自然诱发病圃开展抗瘟性鉴定的做法是可行的。结合两个病圃的数据分析, 确保鉴定结果更可靠。
病圃田间的诱发致病的生理小种是未知的混合小种, 而且不同位置的侵染菌种也可能不尽相同, 加上小气候差异等因素, 所以田间诱发鉴定比人工接种鉴定的可靠性差些。鉴于此, 本试验在材料四周种植混合诱发品种, 尽可能创造同等诱发条件, 而且病圃方差分析和不同时期、不同地点的病情相关性分析都证实了病圃鉴定的可靠性。从病圃中筛选出144份抗不同时期稻瘟病的材料(见附表1), 其中29份苗叶瘟和穗瘟抗性都强的材料可作为稻瘟病抗性基因挖掘的基础材料。
| 附表1 抗不同时期稻瘟病的144份种质信息 Supplementary table 1 Supplementary table 1 Information of 144 accessions with blast resistamce at different stages |
苗瘟与叶瘟间呈极显著强正相关, 而苗瘟与穗瘟、叶瘟与穗瘟之间呈极显著中度正相关性, 表明苗、叶瘟抗性与穗瘟抗性虽然没有必然性, 但具有一定的参考价值。目前还没有确切的研究证实苗叶瘟与穗瘟存在必然的相关性[20, 21, 22, 23]。不同研究人员得出苗、叶瘟与穗瘟间的相关性不一致甚至相反的结果, 可能是由稻瘟病生理小种的高度变异性和不同时期不同区域气候条件的差异引起的。尽管苗、叶瘟抗性不一定会增强穗瘟抗性, 但抗苗叶瘟的材料能保证水稻前期正常生长, 能有效降低后期水稻群体的稻瘟病菌源基数。因此最理想的稻瘟病抗源材料是苗、叶瘟和穗瘟全抗的材料。
农艺性状考察结果显示, 病圃筛选的抗性材料农艺性状参差不齐, 单株产量均值低, 株高偏高, 综合农艺性状较差。抗谱测定结果显示, 30份测定材料中, 有17份抗性频率≥ 70%, 达到中抗以上抗病水平, 抗谱较广。综合病圃鉴定、接种鉴定和农艺性状考察结果, 获得7份优异抗源, 在抗性基因挖掘与品种抗瘟性改良方面具有利用价值。
随着分子技术的快速发展, 国内外学者从水稻抗病种质中鉴定和定位的稻瘟病抗性基因至少有101个[24, 25], 其中至少24个抗性基因已被成功克隆[11]。目前国际上对稻瘟病抗性基因的鉴定, 一般是采用稻瘟病菌株与单基因系或近等基因系联合分析的方法[26, 27]。另外, 近年兴起的高分辨率熔解曲线(HRM)技术在理论上能够区分基因内的所有突变, 且该方法比直接克隆测序法简单、经济、分辨率高[28, 29, 30]。现今, HRM技术已经被用于控制水稻重要性状基因的定位、突变检测和分子标记辅助选择育种[31, 32, 33]。对于本试验获得的抗病材料, 尤其是7份优异抗源, 可结合已知抗瘟基因的功能标记和特异无毒菌株或HRM技术鉴定其中的已知基因和挖掘新基因, 再通过杂交、回交和测序等方式定位克隆其中的新基因, 并通过标记辅助选择聚合不同抗性基因以培育持久抗稻瘟病的优良品种。
对重测序的1217份水稻核心种质资源在恩施两河和芭蕉两个病圃进行稻瘟病抗性自然诱发鉴定, 两病圃的鉴定结果基本一致, 从中获得144份抗苗瘟、叶瘟和穗瘟的抗病种质。选稻瘟病综合抗性较好的34份材料进行30个不同来源的稻瘟病菌株苗期接种鉴定, 筛选出17份抗性频率≥ 70%的广谱抗性材料。鉴定出7份稻瘟病抗性强、抗谱广且农艺性状较好的优异抗源材料IRGA 411-1-6-1F-A、YJ30、金早47、泉珍10号、YN 1353-3、云粳23和IRAT1047, 可作为抗源亲本用于稻瘟病抗性基因挖掘和品种抗稻瘟性改良。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|