


{kind=link}
{kind=link}
{kind=link}
四倍体马铃薯熟性连锁SCAR标记的开发与验证
[李兴翠 , 李广存, 徐建飞, 段绍光, 卞春松, 庞万福, 刘杰, 金黎平
, 李广存, 徐建飞, 段绍光, 卞春松, 庞万福, 刘杰, 金黎平* ]
, 李广存, 徐建飞, 段绍光, 卞春松, 庞万福, 刘杰, 金黎平]
|
|


第一作者联系方式: E-mail:lixingcui1215@163.com
熟性是马铃薯的重要数量性状之一。本研究以马铃薯早熟品种中薯3号和晚熟品种中薯19及其熟性分离群体(221份)为材料, 高通量简化基因组测序和集群分离分析(BSA)相结合, 开发获得一个SCAR标记, 命名为SCAR5-8。进一步利用该标记对分离群体的53份早熟和63份晚熟子代及70份四倍体马铃薯品种进行验证, 结果表明, 该标记在分离群体和四倍体品种中的检测结果与表型鉴定结果总体吻合度分别达87.1%与81.4%, Pearson’s双侧相关分析显示, 其相关性均达极显著水平。该SCAR标记可被用于标记辅助选择, 对加速四倍体马铃薯育种进程具有重要意义。
, LI Guang-Cun, XU Jian-Fei, DUAN Shao-Guang, BIAN Chun-Song, PANG Wan-Fu, LIU Jie, JIN Li-Ping
Maturity is one of the most important quantitative traits in potato. In this study, a maturity linked molecular marker SCAR5-8 was developed using Zhongshu 3, Zhongshu 19 and their F1 segregation population consisting of 221 individuals via reduced-representation sequencing and bulked segregant analysis. A further testing of marker SCAR5-8 was performed using 53 early maturity individuals, 63 late maturity individuals and 70 tetraploid varieties, showing the selection accuracy 87.1% and 81.4% between marker test and phenotyping in segregation population and tetraploid varieties, respectively. Moreover, Pearson’s correlation analysis also showed that the marker SCAR5-8 was significantly correlated to maturity trait in potato. The developed SCAR5-8 marker could be used in marker-assisted selection.
熟性是鉴别马铃薯品种特征的重要依据之一, 也是重要的农艺性状和主要育种目标。不同熟性的品种可以满足不同地区和季节种植和消费需求, 选育不同熟性的品种对于促进马铃薯产业全面发展具有重要意义。表型遗传学研究表明马铃薯熟性性状受微效多基因控制, 且为隐性遗传[1, 2, 3]。利用分子标记方法对马铃薯熟性进行相关研究, 定位出的熟性QTL几乎分布于马铃薯的12条染色体上, 然而多数研究都将熟性主效QTL定位在第5染色体上[4, 5, 6, 7]。利用二倍体材料研究, Jliwka等[8]在第5染色体上定位了一个与分子标记BA47f2t7(P1)紧密连锁的生育期相关QTL, Danan等[9]发现第5染色体上控制熟性的主效QTL与晚疫病抗性QTL紧密相邻, 但互相分开并与分子标记GP21紧密连锁, Kloosterman等[10]通过精细定位, 在第5染色体短臂分子标记GP21附近克隆了马铃薯熟性相关基因StCDF1, 该基因属于DOF转录因子家族, 在马铃薯块茎诱导途径中作为中间调控因子介导块茎形成。
熟性分子标记开发对马铃薯熟性标记辅助选择至关重要, 然而由于马铃薯普通栽培种(2n=4x=48)高度杂合, 四体遗传复杂, 且自交多代后衰退严重, 难以获得重组自交系等分离群体, 分子标记开发困难。而二倍体染色体数目较少, 遗传背景相对简单, 易于进行遗传分析和操作, 所以研究者大多利用二倍体材料开发标记, 再将标记应用到四倍体材料中。在实际的标记辅助选择应用中, 由于四倍体马铃薯在减数分裂过程中的复杂性, 导致染色体交换重组而出现标记与性状偏分离、筛选准确率低等问题。尽管已经有研究者利用四倍体获得了与熟性连锁的分子标记STM3179、c2_476095和_snp_c2_ 22986[11, 12, 13], 但其准确性和稳定性较差, 筛选效率低, 仍难用于四倍体马铃薯熟性育种。因此, 急需在四倍体水平上开发出与熟性紧密连锁的分子标记, 为辅助育种提供新的标记资源, 加速马铃薯熟性育种进程。本研究以四倍体马铃薯熟性分离群体为材料, 基于高通量简化基因组测序和集群分离分析(BSA)开发四倍体熟性分子标记, 并利用群体早熟和晚熟材料及四倍体马铃薯品种进行验证, 以获得与四倍体马铃薯熟性紧密连锁的分子标记, 为马铃薯标记辅助育种提供条件。
1.1.1 四倍体F1代分离群体 以早熟品种中薯3号为父本, 晚熟品种中薯19为母本, 杂交获得221个F1熟性分离后代, 作为开发熟性分子标记的试验材料。
1.1.2 四倍体品种 随机选择70份不同熟性的四倍体马铃薯品种(表1), 作为验证熟性分子标记的试验材料。
| 表1 四倍体马铃薯品种信息 Table 1 Information of tetraploid potato varieties |
2014— 2015连续两年, 将分离群体分别种植于河北省张北县与山东省聊城大学试验基地, 每份材料3株。同时, 2014年将候选早熟材料种植于人工气候室中, 每份材料1株, 模拟大田环境, 进行早熟材料的再次鉴定。播种后20 d左右, 开始调查群体出苗时间, 每5 d调查1次。在出苗后60 d左右开始调查群体生理成熟时间, 每5 d调查1次, 最后计算生育期。根据该分离群体的熟性性状, 按生育期将熟性分为7类, 即极早熟(小于等于60 d)、早熟(61~70 d)、中早熟(71~80 d)、中熟(81~100 d)、中晚熟(101~110 d)、晚熟(111~120 d)、极晚熟(大于等于121 d)。
根据该分离群体的熟性性状, 选取生育期小于75 d的作为极端早熟子代, 生育期大于110 d的作为极端晚熟子代。四倍体马铃薯品种的熟性性状则根据该品种审定时所描述的生育期来判定, 各品种生育期是在适宜生长的区域内, 通过多年多点的品种区域试验, 按照记载标准调查获得的最终结果。
取马铃薯幼嫩叶片, 用CTAB法提取供试材料基因组DNA, 并用琼脂糖凝胶检测DNA质量。
分别采集极端早熟与极端晚熟子代叶片, 等量混合, 构建极端子代DNA混池, 提取方法及质量检测方法同上。
将质量检测合格的亲本DNA与极端子代DNA混池, 进行高通量简化基因组测序。根据测序结果, 获得极端早熟与极端晚熟池间的差异区段。在差异区段中, 根据差异标签的位置, 从马铃薯基因组序列网站(http://solanaceae.plantbiology.msu.edu/pgsc_ downloadShtml)中下载该差异标签左右各700 bp的序列。利用Mega5.2将标签序列与下载的序列比对, 确定该标签的准确性。利用Premier 5引物设计软件设计引物, 上游引物设置在1~550 bp处, 下游引物设置在850~1400 bp处, 引物扩增片段大小设定在400~900 bp之间, Tm值设定在55~63℃之间。最后将设计好的引物送上海生工生物工程有限公司合成。
以晚熟亲本中薯19与早熟亲本中薯3号的基因组DNA为模板进行PCR扩增, 用浓度为1.2%的琼脂糖凝胶电泳检测扩增产物, 查看引物的质量及特异性。PCR体系含5.6 µ L ddH2O, 1.0 µ L缓冲液(10× PCR), 0.8 µ L dNTPs (10 mmol L-1), 0.2 µ L引物F (10 µ mol L-1), 0.2 µ L引物R (10 µ mol L-1), 0.2 µ L Taq酶(2.5 U µ L-1), 2 µ L DNA (25 ng µ L-1)。PCR扩增条件94℃ 3 min, 94℃ 30 s, 59℃ 30 s, 72℃ 50 s, 35个循环; 72℃ 10 min。选择只在某一亲本中扩增出条带, 而另一亲本未扩增出条带的引物, 或者在两亲本中扩增出不同条带的引物开发SCAR标记。最后, 利用极端早熟与极端晚熟子代材料验证, PCR扩增与产物检测方法同上。
分别以马铃薯F1代分离群体的53份早熟、63份晚熟子代及70份四倍体品种的基因组DNA为模
板, 以分子标记SCAR5-8为引物进行PCR扩增, 分别得到扩增产物, 并利用浓度为1.2%的琼脂糖凝胶进行检测产物。
根据分子标记检测结果, 将与早熟亲本和晚熟亲本带型一致的基因型分别记为“ 1” 和“ 0” , 利用统计分析软件SPSS17.0的Pearson’ s双侧检验, 分析马铃薯F1代分离群体中的早熟和晚熟子代材料及四倍体品种的标记检测结果与表型鉴定结果的相关性。
鉴定获得生育期小于80 d的早熟材料53份、生育期大于100 d的晚熟材料63份以及生育期介于80~100 d之间的中熟材料105份; 生育期小于75 d的极端早熟材料35份、生育期大于110 d的极端晚熟材料33份(表2)。
| 表2 熟性表型鉴定结果 Table 2 Results of investigation for maturity |
根据测序结果, 在第5染色体上3.68~4.15 Mb区间内获得极端子代混池间的差异区段, 且该区段与早熟性相关, 区段大小为470 kb, 在该区段中, 共设计引物15对(表3)。以早熟亲本中薯3号和晚熟亲本中薯19基因组DNA为模板对引物的质量与特异性进行检测(图1)。其中引物5-5和5-8只可在早熟亲本中薯3号中扩增出条带, 而在晚熟亲本中薯19中无扩增带。
| 表3 所用引物信息 Table 3 Primer information in this study |
 | 图1 部分引物在亲本中薯3号和中薯19中的扩增产物电泳图Fig. 1 PCR products of some primers in parents Zhongshu 3 and Zhongshu 19 引物编号: 5-1、5-2、5-4、5-5、5-8; 每对引物第1泳道和第2泳道分别为亲本中薯3号和中薯19。 The first lane and the second lane of each primer combination are Zhongshu 3 and Zhongshu 19. |
将在亲本中只扩出某一亲本条带, 而另一亲本无扩增条带的引物, 利用极端性状子代进行验证, 结果发现, 在亲本PCR扩增条带存在差异的2对引物5-5、5-8, 在极端早熟与极端晚熟子代材料中的扩增结果一致。这2对引物在极端早熟子代中的扩增结果与早熟亲本中薯3号一致, 均扩增出条带; 而在极端晚熟子代中的扩增结果与晚熟亲本中薯19一致, 均无扩增条带。因此, 本研究中仅选取了引物5-8用于下一步研究, 并将引物5-8命名为SCAR5-8。
熟性表型鉴定结果表明, 在四倍体马铃薯F1代分离群体的221份子代中, 有53份早熟材料、63份晚熟材料和105份为中熟材料, 其中的53份早熟材料和63份晚熟材料用于SCAR5-8的标记检测(图2和表4)。在标记为阳性(有PCR扩增产物)的54个材料中有46个表型为早熟, 分子标记检测与表型鉴定结果吻合度达85.2%; 在标记为阴性(无PCR扩增产物)的62个材料中有55个表型为晚熟, 分子标记检测与表型鉴定结果吻合度达88.7%。116个材料的标记检测结果与表型鉴定结果的总体吻合度达87.1%, 将表型鉴定结果与分子标记检测结果进行Pearson’ s双侧相关分析, 相关系数r为0.740, 且达极显著水平。上述结果表明, 分子标记SCAR5-8与四倍体马铃薯熟性性状紧密连锁, 能较好地区分四倍体马铃薯分离群体后代的熟性, 可通过标记辅助选择用于四倍体马铃薯熟性育种。
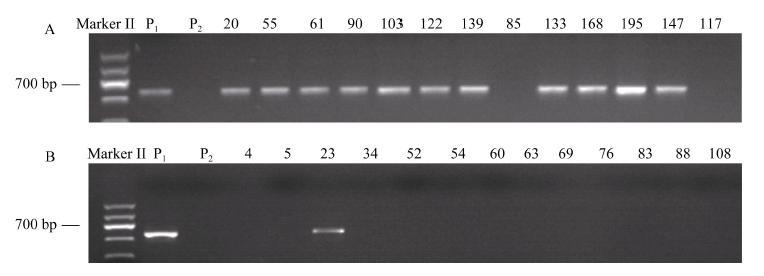 | 图2 SCAR5-8标记在部分早熟(A)与晚熟(B)子代中的扩增结果Fig. 2 Amplification results of SCAR5-8 in early (A) and late (B) maturity progenies P1: 早熟亲本中薯3号; P2: 晚熟亲本中薯19; 数字: 依次为早熟与晚熟子代材料编号。 P1: early parent Zhongshu 3; P2: late parent Zhongshu 19; Number: code of early and late maturity progenies. |
| 表4 表型鉴定和标记检测统计结果 Table 4 Statistics results of phenotyping and molecular marker test |
熟性统计结果表明, 在70份四倍体马铃薯品种中, 生育期小于75 d的早熟品种有27份, 占总材料数的38.6%; 大于110 d的晚熟品种有43份, 占总材料数的61.4% (表1)。利用分子标记SCAR5-8对70份四倍体马铃薯品种检测结果表明, 在标记为阳性的24个品种中有19个表型登记为早熟, 分子标记检测与表型登记结果吻合度达79.2%; 在标记为阴性的46个品种中有38个表型登记为晚熟, 分子标记检测与表型登记结果吻合度达82.6% (图3和表5)。70个品种的标记检测结果与表型登记结果的总体吻合度达81.4%。将分子标记检测结果与表型登记结果进行Pearson’ s双侧相关分析, 相关系数r为0.602, 达极显著水平。上述结果表明, 分子标记SCAR5-8能较好地区分四倍体马铃薯品种的熟性, 该标记可以用于四倍体马铃薯熟性的标记辅助选择。
 | 图3 SCAR5-8标记在部分早熟(A)与晚熟(B)品种中的扩增Fig. 3 Amplification of SCAR5-8 in early (A) and late (B) maturity varieties 1~15: 中薯2号、中薯4号、中薯6号、中薯8号、郑薯5号、郑薯6号、早大白、克新5号、克新9号、黄麻子、春秋7号、春薯2号、双丰4号、川芋56、超白; 16~30: 青薯4号、青薯8号、青薯9号、宁薯5号、宁薯7号、晋薯2号、晋薯4号、高原2号、高原6号、燕子、中薯21、云薯401、天薯5号、天薯9号、陇薯5号。 1-15: Zhongshu 3, Zhongshu 4, Zhongshu 6, Zhongshu 8, Zhengshu 5, Zhengshu 6, Zaodabai, Kexin 5, Kexin 9, Huangmazi, Chunqiu 7, Chunshu 2, Shuangfeng 4, Chuanyu 56, Chaobai; 16-30: Qingshu 4, Qingshu 8, Qingshu 9, Ningshu 5, Ningshu 7, Jinshu 2, Jinshu 4, Gaoyuan 2, Gaoyuan 6, Yanzi, Zhongshu 21, Yunshu 401, Tianshu 5, Tianshu 9, Longshu 5. |
| 表5 表型登记和标记检测统计结果 Table 5 Statistics results of phenotype registration and molecular marker test |
获得与目的基因紧密连锁的分子标记是构建遗传图谱及标记辅助育种的基础, RAPD、AFLP、SSR和SRAP等方法极大地推动了分子标记辅助育种技术的研究, 如Bradshaw等[14]利用RFLP标记结合BSA分析方法筛选到2个与抗晚疫病连锁的标记, 雷剑等[15]利用BSA方法筛选到与马铃薯青枯病抗性基因连锁的SRAP标记。但由于其复杂程度和通量的限制, 以及四倍体马铃薯遗传的复杂性, 难以满足大规模分子标记辅助育种的需要。本研究利用高通量简化基因组2b-RAD测序技术结合集群分离分析(BSA)法直接对四倍体马铃薯进行了遗传区段的挖掘与标记SCAR5-8的开发, 并在四倍体分离群体中进行了标记的验证。2b-RAD技术避免了其他RAD技术中片段大小选择的过程, 使标签在基因组中的分布更加均匀, 对于复杂基因组的标记开发和染色体区段的挖掘更为适用。因此, 该方法解决了马铃薯四倍体材料标记开发困难的问题, 且在标记的实际应用中, 不易出现应用二倍体标记筛选四倍体材料时出现的标记与性状偏分离、筛选准确率低等问题。因此, 在四倍体马铃薯熟性选择中, 实用性更强, 筛选效率及准确性更高, 对于马铃薯遗传研究和品种选育更有意义。
分子标记辅助育种技术极大提高了育种中性状选择的准确性和效率。到目前为止, 马铃薯上已有抗晚疫病、抗病毒病、抗青枯病、薯肉颜色、炸片颜色、薯形等几十个性状被标记[16, 17, 18, 19, 20, 21], 但关于马铃薯熟性紧密连锁的标记开发的报道极少, 尽管已获得一些与熟性相关的标记, 但这些标记筛选准确性差、效率低, 难以用于四倍体马铃薯分子辅助育种。本研究所开发的标记SCAR5-8, 在116个分离群体材料与70个育成品种的标记检测结果与表型鉴定结果的总体吻合度分别达87.1%与81.4%, 且均达极显著水平, 可直接用于不同熟性材料的筛选, 而且准确性高。此外, SCAR标记是显性遗传, 在实际应用中, 方便、快捷、稳定性强, 可以快速检测大量个体。
分子标记对目标性状辅助选择的准确性是一个核心问题。尽管本研究开发的标记SCAR5-8在四倍体品种应用检测中具有较高的符合率, 但仍有18.6%的四倍体品种的分子标记检测结果与表型描述结果不一致, 可能有以下几方面的原因。首先, 熟性是数量性状, 除了受主效基因控制外, 可能还受其他微效基因的影响, 如控制马铃薯熟性主效QTL位于第5染色体上, 但是在第3和其他染色体上还存在一些微效QTL [14, 22], 这些微效QTL都会影响标记关联度。其次, 块茎是马铃薯的经济器官, 在马铃薯品种审定和实际应用中, 生育期(即熟性)是按植株生理成熟期和(或)块茎成熟(块茎膨大早、块茎膨大快)等分类, 这两种不同类型的熟性可能受不同基因控制, 而本研究基于植株生理成熟类型开发了标记SCAR5-8, 分子标记检测结果与表型鉴定结果不一致的材料有可能属于不同熟性类型。第三, 国内育成品种遗传背景狭窄, 血缘关系相近, 对试验中采用的早熟品种系谱分析表明, 除少数品种外, 分成了2个类群, 一个是米拉的后代, 大多数都有SCAR5-8扩增条带, 表现为阳性; 另一个是白头翁无带)的后代, 大多无带, 表现为阴性。也就是说, 不同系谱里的品种, 其熟性性状的遗传基础可能存在差异, 而同一系谱里的品种可能有相同的遗传基础。但由于所用品种较少, 不足以将所有试验用品种联系起来进行全面完整的系谱分析, 若能增加足够的品种, 将可获得更为理想的结果。除此之外, 环境因素及人工鉴定误差也会影响结果的准确性。因此, 在进行马铃薯熟性分子标记开发时, 最好结合多年的田间熟性鉴定结果; 在进行标记辅助选择时, 选用多个熟性连锁标记或功能基因标记同时进行检测, 结果将更加准确可靠。
本研究开发的分子标记SCAR5-8用于鉴定筛选四倍体熟性材料时, 早熟材料可扩增出条带, 晚熟材料则扩不出条带。该标记可应用于四倍体马铃薯品种的标记辅助选择, 筛选效率较高。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|