



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
一个棉花纤维伸长期优势表达启动子pGhFLA1的克隆与鉴定
[胡海燕 , 刘迪秋, 李允静, 李阳, 涂礼莉
, 刘迪秋, 李允静, 李阳, 涂礼莉* ]
, 刘迪秋, 李允静, 李阳, 涂礼莉]
|
|


第一作者联系方式: E-mail:yanhai0987@163.com
棉花纤维是研究单细胞生长发育的良好模式细胞, 寻找在棉花纤维发育时期优势表达的启动子, 对于棉花纤维发育的功能基因组研究非常重要。本试验对发掘到的一个在纤维伸长期优势表达基因 GhFLA1的启动子进行克隆, 并将该启动子融合GUS转化棉花和烟草。转基因棉花的GUS蛋白染色表明, 在0~20 DPA (days post anthesis)的纤维中检测到GUS蛋白, 同时在柱头、雄蕊、萼片、胚根、子叶和根中也检测到GUS蛋白, 但在花瓣、叶片和苞叶中没有检测到。同时GUS定量检测结果显示, pGhFLA1驱动GUS蛋白积累的高峰是在20 DPA的纤维中, 其驱动GUS表达的能力是CaMV 35S::GUS的22倍。转基因烟草组织化学染色显示pGhFLA1可以驱动GUS在叶片及叶片的表皮毛中优势表达, 同时在子房、柱头、花药、苞叶、花柄基部、花瓣、种子等生殖器官上也存在GUS蛋白。启动子pGhFLA1在棉花纤维和烟草叶片的表皮毛中优势表达, 可用于棉花纤维功能基因组研究, 在改良纤维品质育种中具有潜在的应用价值。
Cotton ( Gossypium hirsutumL.) fiber is a perfect model cell for studying cell initiation and growth, so it is critical to find new promoters specifically or preferentially expressed in fiber for functional genomics of fiber and improvement of cotton fiber yield and quality. We found a gene GhFLA1 with very high expression level in fiber elongation stage and cloned its promoter (pGhFLA1). pGhFLA1::GUS was transferred into cotton YZ1 ( Gossypium hirsutum) and tobacco ( Nicotiana tabacum) to identify its expression pattern in different plant tissues. GUS activity in transgenic plant was obviously detected in fiber from 0 DPA (days post anthesis) to 20 DPA, also in stamens, pistils, sepal, radicle, cotyledon and root, but not in petal, leaf and bract. Quantitative analysis of GUS activity showed that GUS expression level driven by pGhFLA1 was the highest in 20 DPA fiber and 22-fold higher than that in CaMV 35S. In transgenic tobacco, GUS activity was detected in leaf and trichome and reproductive organs like ovary, stigma, anther, bract, petal and seed. In general, the promoter pGhFLA1 fusion with GUS highly expresses in cotton fiber and tobacco trichome, which could be used in fiber functional genomics research and improvement of cotton fiber quality.
棉花纤维是由胚珠表皮细胞突起发育而来。从开花当天开始, 棉花纤维经历启始、伸长、次生壁增厚和脱水成熟相互重叠的4个阶段, 最后发育成一个长达3 cm左右, 次生壁中纤维素含量达90%以上的纤维细胞[1]。因此棉花纤维细胞是研究植物单细胞发育及纤维素沉积的良好材料。目前, 棉花的基因工程中普遍使用的是组成型CaMV 35S启动子, 使用组成型启动子存在诸多缺点, 例如CaMV 35S启动子驱动的目的基因在受体植物各个组织中持续且恒定的表达会过度消耗能量; 在受体不表达的部位表达目的基因会使植株的形态改变, 不能达到预期目的[2]。因此分离鉴定棉花纤维优势表达的启动子, 无论是在棉花纤维发育机制解析还是在分子育种中都具有重要的应用价值。
目前已经克隆了一些棉花纤维优势表达启动子, 其中最早克隆出来的棉花纤维优势启动子E6表达高峰在开花后5~28 DPA (days post anthesis), 而在其他组织中不表达[3]。FbL2A启动子在20 DPA纤维中优势表达, 活性约为35S启动子的1/3[4]。棉花纤维发育相关的脂转移蛋白基因(LTP3、LTP4和LTP6)的启动子可以驱动GUS在烟草表皮毛中优势表达, 但是这几个启动子的启动能力很弱, 其中LTP6基因启动子的启动活性仅为CaMV 35S启动子的1/1000左右[5, 6, 7, 8, 9]。棉花纤维伸长期优势表达启动子pGbEXPA2在棉花纤维伸长期和拟南芥的表皮毛中优势表达[10]。CFSP启动子可以在逆境条件下启动基因在纤维发育时期优势表达[11]。pGhSCFP在烟草外种皮优势表达, 在转化棉花突起的纤维细胞特别是伸长期纤维中优势表达[12]。启动子GbPDF1-1能够驱动GUS在0~5 DPA胚珠和纤维细胞中表达[13]。一些外源启动子也能够驱动目的基因在棉花的种皮和纤维细胞中优势表达。Luo等[14]发现矮牵牛FBP7基因的启动子pFBP7在烟草中具有种皮表达特异性, 并在-1~10 DPA的棉花胚珠和纤维中优势表达。以来源于拟南芥的BAN启动子融合GUS转化棉花, 其0 DPA的胚珠表面纤维细胞或非纤维细胞中都有较强的GUS信号, 5 DPA的胚珠GUS活性最强, 且在30 DPA的纤维中仍能检测到GUS的表达[15, 16]。
Liu等[17]在20 DPA的纤维SSH cDNA文库中发掘到一个在陆地棉纤维伸长期优势表达基因GhFLA1, 并对GhFLA1进行qRT-PCR与Northern杂交表达验证发现, GhFLA1在下胚轴中有微量的表达, 在纤维发育的不同时期优势表达, 且在15 d时纤维中表达量最高, 而在根、叶片、花中没有表达。GhFLA1在海岛棉和陆地棉之间的表达模式比较相似, 但前者表达强于后者[18]。前人已经克隆及验证了一些在棉花纤维伸长期优势表达的启动子, 但是这些启动子驱动GUS的能力较CaMV 35S弱很多。本研究克隆并验证了GhFLA1的启动子pGhFLA1 (promoter-GhFLA1)在棉花和烟草中的表达模式, 旨在探索棉花纤维发育各个时期驱动GUS的能力和优势。
于2010年在华中农业大学试验田进行田间试验, 在华中农业大学作物遗传改良国家重点实验室进行室内试验。受体作物为陆地棉YZ-1 (Gossypium hirsutum)和烟草(Nicotiana tabacumL.)。
通过Genome-walking的方法扩增得到基因GhFLA1的启动子序列并进行PCR。PCR产物经过TA克隆连接到T载体上, 测序后进行顺式元件分析。对已经克隆到的启动子pGhFLA1构建植物表达载体, 并在引物上游和下游分别添加Hind III和BamH I酶切位点及保护碱基, p-FLA-S: 5° -GCAAAG CTTTCTATAGCATGGAGCTAGGTGGT-3° ; p-FLA- A: 5° -GCAGGATCCGTCTTTTGGTTTGTGTTGGAA AGG-3° 。将PCR产物纯化后用Hind III和BamH I酶切, 经T4连接酶连接到本实验室改造的启动子载体PBI121上。将构建好的启动子载体pBI-PGhFLA1转化到农杆菌菌株EHA105中。
以pGhFLA::GUS和CaMV 35S::GUS转化陆地棉YZ-1, 采用农杆菌介导的下胚轴遗传转化法[19]。
采用农杆菌介导的叶盘转化法以PGhFLA:: GUS转化烟草。用75%乙醇或无菌水浸泡烟草种子30 s, 0.1%升汞浸泡10 min, 无菌水冲洗3~5次, 每次1 min, 将无菌的种子放在1/2 MS培养基上, 于25℃下培养直至烟草的叶片生长到适合转化的时期。
取烟草叶片于无菌平皿上, 用无菌剪刀将烟草叶片剪切成约1 cm2的小片, 接种到MGL悬浮的农杆菌菌液中, 侵染5~10 min后将叶片放到铺有一层滤纸的共培养培养基上, 21℃暗培养3 d, 转到筛选培养基上(含50 mg L-1的卡那霉素, 400 mg L-1的头孢霉素)培养并筛选2次, 将烟草苗移植到土壤中。
取转基因植株的幼嫩叶片0.2~0.3 g, 并用植物基因组提取试剂盒DP305 (北京天根)提取基因组DNA。取10 μ g转基因植物的基因组DNA, 用Hind III酶切48 h后, 凝胶电泳检测DNA是否被彻底酶切。将彻底酶切后的DNA混合适量溴酚蓝, 并用0.8%的0.5× TBE琼脂糖凝胶, 于0.5× TBE电泳缓冲液中, 250 V高压电泳10 min, 接着电压调至40 V继续电泳12~14 h。将电泳后的胶条上的溴酚蓝和点样孔之间的凝胶小心整齐地切下; 随后酸变性10 min, 碱变性20 min, 碱转移液处理15 min。然后用碱转移液作为转移缓冲液转膜约36 h, 用2× SSC漂洗膜2次(各5 min), 80℃真空烘膜2 h以上固定DNA。
先将尼龙膜用2× SSC漂洗后装入杂交管, 加入65℃预热的预杂交液10 mL, 赶除气泡后置于65℃空气恒温杂交炉上预杂交6 h。将标记好的Npt II探针沸水变性10 min, 冰浴10 min后一起加入杂交管, 65℃空气恒温杂交炉上杂交18 h以上。用低严谨的洗膜液I先热洗10 min, 再洗5~10 min之后, 将膜平摊于滤纸上吸干表面水分, 用保鲜膜包裹, 平铺在磷屏上(Fuji Photo Film Co., Ltd., Japan)过夜, 用FLA-5000 Plate/ Fluorescent Image Analyzer (Fuji Photo Film, Tokyo, Japan)扫描磷屏成像。
将转基因植株的不同器官、组织放到GUS染色液中, 37℃保温2 h至过夜, 经70%酒精脱色后FAA固定液中保存, 肉眼或在体式显微镜下观察GUS基因表达情况, 然后在体式显微镜(Leika MZFLIII)下照相。GUS染色液成分为X-gluc 90 mg、氯霉素10.0 mg、0.1 mol L-1磷酸钠缓冲液(pH 7.0) 50 mL、甲醇20% (v/v), 补足无菌水至100 mL。
采用Jefferson等[20]报道的方法定量检测GUS。在1 mL离心管中加入400 µ L GUS蛋白提取缓冲液、100 µ g GUS蛋白、10 µ L 40 mmol L-1 4-MUG, 37℃反应1 h后加入1.6 mL反应终止液终止反应。以50 nmol L-1的4-MU为标准, 在激发光365 nm发射光455 nm条件下测定标样吸光值。以pmol 4-MU μ g-1 min-1表示GUS活性。
采用Genome-walking的方法从陆地棉中克隆到了长961 bp的pGhFLA1启动子序列, 利用启动子结构预测网站(http://bioinformatics.psb.ugent.be/webtools/ plantcare/html/)进行结构预测发现该启动子有4个CAAT BOX和4个TATA BOX。同时检测到多个光响应和激素响应位点, 如脱落酸、茉莉酸等(图1)。
 | 图1 pGhFLA1核酸序列及作用元件Fig. 1 Sequence of pGhFLA1 and predicted cis-acting elements 方框中为CAAT BOX; 椭圆中为TATA BOX; 红色下画线表示茉莉酸响应位点; 绿色下画线表示光响应位点; 蓝色下画线表示脱落酸响应位点。 Square box indicates CAAT BOX, oval indicates TATA BOX, red underline indicates JA responsive elements, green underline indicates light responsive elements, blue underline indicates ABA responsive element. |
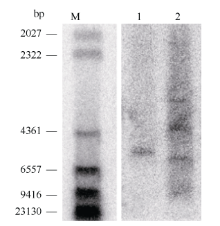
GhFLA1是一个在纤维伸长期优势表达的基因。通过Southern验证2个独立的pGhFLA1::GUS转化株系的拷贝数发现, 其中一个是单拷贝, 一个多拷贝(图2)。
 | 图2 转基因植株的拷贝数Fig. 2 Southern blotting analysis of transgenic cotton lines M: marker; 1, 2: 不同的转化事件。 M: marker; 1, 2: two different transgenic cotton lines. |
从田间的阳性株系上取不同时期的胚珠、纤维、叶片、苞叶及花组织, 及萌发1 d的胚和萌发7 d的下胚轴和根进行GUS组织化学染色表明, 纤维整个发育时期(0~25 DPA)及刚萌发的子叶、幼嫩的根尖、雌雄蕊和根中都检测到GUS蛋白, 但在花瓣、叶片、苞叶中没有检测到GUS蛋白(图3)。为了验证该启动子在纤维发育不同时期驱动GUS能力的强弱, 分别定量测定5 DPA (纤维/胚珠), 10、15、20和25 DPA的纤维组织的GUS活性。结果表明启动子pGhFLA1在5~25 DPA的纤维中都有表达, 但是在10 DPA的表达量最低, 在20 DPA的表达量达到一个峰值, 达到34 938.04 pmol4-MU μ g-1 min-1(图4)。所以可以确定pGhFLA1是一个伸长期及次生壁增厚转换时期优势表达的启动子。
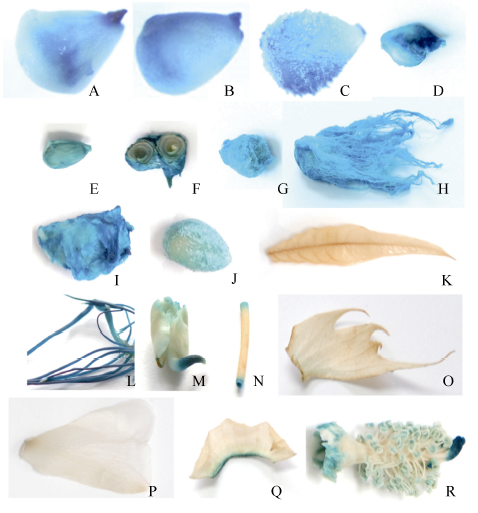 | 图3 组织化学染色分析启动子pGhFLA1:GUS的表达模式Fig. 3 Histochemical analyses of transgenic cotton plants containing pGhFLA1::GUS A: 0 DPA; B: 1 DPA; C: 3 DPA; D: 10 DPA; E: 10 DPA胚珠; F: 15 DPA胚珠; G: 20 DPA胚珠; H: 20 DPA; I: 25 DPA; J: 25 DPA胚珠; K: 叶片; L: 根; M: 胚; N: 下胚轴; O: 苞叶; P: 花瓣; Q: 萼片; R: 雌雄蕊。A: 0 DPA; B: 1 DPA; C: 3 DPA; D: 10 DPA; E: 10 DPA ovules; F: 15 DPA ovules; G: 20 DPA ovules; H: 20 DPA; I: 25 DPA; J: 25 DPA ovules; K: leaf; L: root; M: embryo; N: hypocotyl; O: bract; P: petal; Q: sepal; R: pistil and stamen. |
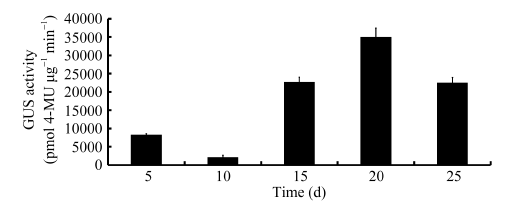 | 图4 启动子pGhFLA1在棉花纤维发育不同时期的GUS活性Fig. 4 GUS activity of transgenic plants containing pGhFLA1::GUS at different fiber development stages |
为了与最常用的CaMV 35S启动子进行对比, 我们从CaMV 35S::GUS转化棉花的转基因纯系植株上同时取5 DPA (纤维/胚珠), 10、15、20和25 DPA的纤维进行GUS活性定量检测, 发现在20 DPA pGhFLA1::GUS的活性是CaMV 35S::GUS的22倍(图5)。
 | 图5 CaMV 35S启动子在纤维发育不同时期纤维组织的GUS活性定量测定Fig. 5 GUS activity of transgenic plants containing CaMV 35S promoter fragment/GUS fusions at different fiber development stages |
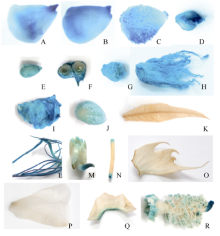
以pGhFLA1::GUS转化烟草, 取阳性植株的不同组织进行GUS染色表明GUS蛋白表达主要集中在子房、柱头、雌蕊、花药、苞叶、花柄基部、花瓣、种子等生殖器官。值得注意的是在叶片及叶片的表皮毛上染色尤其明显, 表明pGhFLA1可能在这些部位驱动GUS优势表达(图6)。
 | 图6 组织化学染色分析启动子pGhFLA1::GUS在烟草不同组织中的表达模式Fig. 6 Histochemical analyses of GUS activity in different tissues of transgenic plants containing pGhFLA1 promoter fragment/GUS fusions A: 子房; B: 柱头; C: 雄蕊; D和F: 花柄; E: 叶柄; G和I: 苞叶; H: 花瓣; J: 种子; K和L: 叶片; M和N: 表皮毛。 |
A: ovary; B: stigma; C: stamen; D and F: peduncle; E: petiole; G and I: bract; H: petal; J: seed; K and L: leaf; M and N: epidermal.
传统的育种方法对棉花产量潜力的挖掘和纤维品质的提高在近年来进展不大, 生物技术的快速发展及其在农业上的应用为作物育种和生产提供了新途径。基因功能验证和转基因育种除了对目标基因有要求, 启动子也是一个不可忽视的方面。在研究棉花纤维发育机制过程中, 传统的CaMV 35S启动子所驱动的基因表达系统, 不能排除基因对植株整体发育的影响从而影响纤维生长发育。虽然已经有大量的内源及外源的启动子在纤维不同发育时期优势表达, 但是真正在基因工程中验证能被使用的却为数不多, 因此研究和开发在纤维发育时期优势表达的启动子势在必行。
棉花转化有一定的难度, 许多研究者认为棉花纤维的发育可能与烟草和拟南芥的表皮毛发育存在相似性[21], 因此很多研究用烟草和拟南芥来验证棉花启动子的表达模式。在已有的报道中, 将棉花纤维中优势表达的基因的启动子在烟草和拟南芥中异源表达后, 能够驱动GUS在表皮毛或根中优势表达。如以棉花脂转移蛋白基因Ltp6的启动子转化烟草后只在烟草表皮毛中表达[8], 棉花可逆糖基化多肽基因GhRGP1启动子转化烟草, 在生长4周的植株的根中表达量最高[22]。棉花葡糖醛酸基因GhGlcAT1启动子转化烟草后, 在烟草的根冠、种皮、花粉粒和表皮毛中优势表达[23]。纤维素合酶基因GhCesA4启动子转化烟草, 在烟草的表皮毛和叶脉中优势表达, 同时也在下胚轴和茎的维管组织中表达[24]。RING-type泛素连接酶(E3)启动子转化拟南芥, 在叶、根、表皮毛、花药、柱头等部位表达[25]。GaMYB2启动子转化棉花, 其在纤维中优势表达, 同时也在表皮毛、叶片、苞叶、茎等组织表达, 转化烟草和拟南芥表明其在表皮毛中特异表达[26]。本研究同时将pGhFLA1::GUS转化了棉花和烟草, 棉花染色结果表明该启动子不仅在纤维发育各个时期表达, 同时在幼嫩的根尖、雌雄蕊及根中都有表达。烟草的染色结果表明除了在表皮毛中优势表达, 在花的各个组织中都检测到GUS蛋白的存在。说明棉花与烟草的染色结果存在一致性, 所以用烟草或拟南芥验证棉花启动子的表达模式有一定的可行性。
基因启动子是重要的基因表达调控元件, 决定了基因的转录方向、转录效率以及转录的时空特性[27]。运用基因工程手段对农作物进行遗传改良, 赋予农作物一些新性状, 需要外源基因在受体内以高度受控方式表达, 因此对启动子的表达特性提出了很多新要求。例如要求启动子能接受外部信号刺激, 引导基因在特定器官组织和细胞类型的特定发育阶段表达, 这对于外源基因能否在特定的组织优势高效表达至关重要[29, 30, 31]。启动子结构预测表明, pGhFLA1上存在一些光信号及激素响应位点, Huang等[32]研究发现GhFLA1可能受到激素的调控从而影响纤维的伸长, 因此启动子预测对于基因功能的研究具一定指导作用。
已经报道的启动子中有很大一部分启动能力很弱, 其中有一个棉花泛素相关启动子uceA1.7在花组织里面的表达量是CaMV 35S的7倍之多[33]。pGhFLA1::GUS的组织化学染色结果显示, pGhFLA1可以驱动目的基因在纤维中优势表达, 且表达高峰在15~25 DPA, 在20DPA为CaMV 35S的22倍之多。因此, pGhFLA1是纤维发育的高效优势表达的启动子, 在纤维伸长与次生壁增厚转换时期其启动能力强于CaMV 35S, 并且可以用于提高基因在纤维伸长与次生壁增厚转换时期的表达。
棉花启动子pGhFLA1是一个纤维优势表达的启动子, 在棉花纤维发育的伸长及次生壁增厚的转换时期及烟草表皮毛中优势表达, 并且在20DPA驱动GUS表达的能力是CaMV 35S的22倍, 可作为棉花纤维功能基因组研究的优良启动子和运用于未来的棉花分子育种。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|