

{kind=link}
{kind=link}
{kind=link}
不同栽培模式对油菜产量和倒伏相关性状的影响
[蒯婕1  , 左青松
, 左青松2 , 陈爱武3 , 程雨贵4 , 梅少华5 , 吴江生1 , 周广生1, * ]
, 左青松]
|
|


第一作者联系方式: E-mail:kuaijie@mail.hzau.edu.cn
为建立与油菜相匹配的高产高效栽培管理方式, 设置3种栽培模式即常规栽培(FP)、超高产栽培(SP)和高产高效栽培(HH), 于2013—2014年在湖北枝江单季稻区用中熟油菜品种华双5号, 武穴双季稻区用早熟品种华早291选择不同肥力田块(高肥力、低肥力)进行试验, 测定3种栽培模式下油菜生物量和生育期间的光能资源利用率、产量、田间倒伏等指标。结果表明, 与FP相比, 在高、低肥力下, SH和HH均提高了各时期的光能截获率和光能利用效率, HH模式薹肥施用比例高, 在后期光能截获率下降速率最低, 仍保持较高的光合面积, 有利于干物质的积累。SH模式和HH模式下, 收获指数和产量均显著高于FP模式, 且以SH模式最高。株高、根冠比和抗折力均表现为SH>HH>FP; 倒伏指数与倒伏角度的变化趋势较为一致, 在不同地力条件下均表现为SH > FP > HH。综上, HH模式的籽粒产量虽略低于SH模式, 但不显著, 而后期倒伏显著降低。与SH模式相比, HH模式通过增加种植密度, 减少氮肥投入和施肥次数, 起到了“以密抗倒、以密省肥”的效果, 机械收获效率显著提高, 可实现高产高效栽培。
, ZUO Qing-Song
To establish a reasonable cultivation and management mode matching canola production, three cultivation modes including farmers practice cultivation (FP), super high cultivation (SH) and high yield and high efficiency cultivation (HH) were adopted with middle-mature winter canola Huashuang 5 in single cropping rice region (Zhijiang) and early-mature winter canola Huazao 291 in double-rice cropping region (Wuxue) of Hubei province at high and low soil fertility levels. Biomass accumulation, radiation use efficiency (RUE), yield and lodging related indices were investigated and analyzed. The light inception rate (LIR) and RUE under SH and HH were higher than those under FP during whole growth period. The deceleration rate of LIR at the end of growth stage was least under HH as the rate of bolting nitrogen increased, which was benefit for biomass accumulation. The harvest index and yield of SH and HH were significantly higher than those of FP. Although HH had higher yield than SH, the difference was not significant. The lodging index of HH was lower than that of SH which indicated that HH had higher lodging resistance. All these revealed that HH could achieve high yield and high lodging resistance with lower nitrogen cost compared with SH. It is concluded that the HH cultivation model can promote the mechanical harvesting efficiency by increasing lodging resistance and reducing nitrogen cost due to higher plant density. Therefore, HH should be encouraged to extend in the Yangtze River Valley in the future for increasing mechanical canola productivity.
油菜既可为人类提供食用植物油, 也可作为畜牧业的蛋白饲料来源[1], 发展油菜生产意义重大。油菜生产中, 机械化收获能显著降低油菜成本、提高生产效率。但目前我国的油菜机械化作业面积仍处于较低水平[2], 其主要原因是, 机械收获作业效率较低且收获损失率居高不下。成熟期倒伏则是产生该问题的重要因素[3]。合理的栽培管理措施和适宜的温光水肥条件是油菜高产与抗倒相互协调的基础[2, 3]。当前全球气候多变的背景下, 温光等资源的分配有更大的不确定性[4, 5]; 同时油菜生产中肥料的不合理施用不仅激化了高产与抗倒的矛盾, 而且也造成了严重的环境污染和资源浪费。因此, 通过合理的栽培措施来解决上述难题是实现油菜高产抗倒相互协调的重要途径[6]。氮肥和密度是作物高产栽培的重要途径, 同时也影响着作物倒伏的程度[7]。光照是作物生长的重要环境因子, 不同氮肥用量和种植密度下, 作物群体结构不同, 导致光照拦截以及光能利用率等的不同, 最终影响群体产量和抗倒伏能力。通过适宜的施氮量和密度配置, 构建合理冠层结构, 可提高作物光拦截量和光能利用率, 达到增产的效果[8]。一定范围内, 增施氮肥, 可提高作物光能利用率[9], 但氮肥利用效率下降, 且过量施用氮肥易导致植株旺长, 茎秆充实度下降, 增加倒伏风险[10], 倒伏后, 作物光能利用率下降[11], 产量降低, 且不利于机械收获。一定范围内, 增加种植密度可增加群体光合面积和光能利用率, 充分发挥作物群体效应而提高产量[6]。但我国油菜生产中, 种植密度一般为15~30万株 hm-2, 与欧洲国家相比, 密度偏低, 导致产量较低[12]。关于密度对作物倒伏的影响, 多数认为, 密度增加, 倒伏加重[13, 14]。如水稻密度从40兜 m-2增加到160兜 m-2, 株高虽降低, 但是倒伏指数增加, 抗倒性减弱[15]。而油菜相关研究则表明, 一定范围内, 密度增加, 抗倒性增强[16]。因此, 优化种植方式, 协同提高产量与抗倒性, 是油菜生产的关键。油菜产量和抗倒性受肥料、种植密度、播期和灌溉等诸多因素的影响。前人的研究多集中在肥料、种植密度、播期等单一或两因素的互作效应, 如何将上述因素整合成为优化栽培模式及其对产量和倒伏的调控效应尚不清楚。本试验通过研究高、低肥力水平下不同栽培模式对油菜冠层光能截获和利用效率、根系生长发育及其对产量和抗倒性的影响, 从光能吸收利用和干物质生产的角度探讨提高油菜产量和抗倒性的可行性及进一步的优化措施, 为油菜高产高效栽培技术的制定提供科学依据。
试验于2013— 2014年度在湖北武穴和枝江2个肥力条件(高肥力和低肥力)下进行, 0~20 cm土壤养分见表1。武穴试点油菜品种为早熟品种华早291, 播期为2013年9月25日, 移栽期为2013年11月9日至10日, 2014年5月6日80%角果变黄时收获。枝江试点油菜品种为中熟品种华双5号, 2013年10月7日直播, 2014年5月12日80%角果变黄时收获。
依据当地油菜生产现状, 结合前期试验研究基础, 选择不同地力田块, 设常规栽培管理方式(FP)、超高产栽培管理方式(SH)和高产高效栽培管理方式(HH), FP处理中, 设零氮对照区(CK)。完全随机区组设计, 4次重复, 小区面积20 m2。N、P2O5、K2O肥料分别为尿素(含N 46%)、过磷酸钙(含P2O5 12%)和氯化钾(含K2O 60%)。磷肥和钾肥用量均为150 kg hm-2; 硼砂用量为7.5 kg hm-2。磷、钾、硼肥均作基肥施用。氮肥运筹和不同栽培模式参数配置见表2, 其他田间管理同常规。
| 表1 不同试点耕层0~20 cm养分状况 Table 1 Chemical properties of tested soil |
| 表2 不同田间管理模式设计 Table 2 Schematic representation of experimental design and treatments |
1.3.1 关键生育时期群体透光率 用冠层分析仪(SUNSCAN Canopy Analysis System)测定群体透光率指标, 计算各小区光拦截量, 测定日期见表3。
| 表3 不同试验点生育进程差异 Table 3 Growth stages in two sites |
1.3.2 成熟期农艺性状和产量 用冠层最高点至子叶节连线与垂直方向夹角表示田间倒伏角度[17]。人工收获面积为16 m2的油菜, 晾晒5~7 d后脱粒、扬净、晒干、称重, 得出各小区籽粒产量。从各小区连续取样15株, 将植株带土挖出, 用流水冲洗根系表面的土壤, 最大程度保持根系完整。考察株高、根长、根鲜重、根颈粗。株高以子叶节至植株顶端的高度表示; 根颈粗为游标卡尺测定的子叶节下1 cm粗度; 地上部鲜重为植株子叶节以上部鲜重; 根鲜重为子叶节以下部鲜重; 将根系及地上部置105℃杀青30 min、80℃烘干至恒重后, 测定干物质量并计算根冠比。以YYD-1型茎秆强度测量仪(浙江托普仪器有限公司生产)测定茎秆基部10 cm处抗折力; 倒伏指数=(株高× 地上部分鲜重)/抗折力[18]。按文献[19]: 营养器官花前贮藏物质转运量=开花期营养器官干重-成熟期营养器官干重; 营养器官花前贮藏物质转运率 = (开花期营养器官干重-成熟期营养器官干重)/开花期营养器官干重× 100%。
采用SPSS 11.0统计软件进行数据统计和方差分析, 以最小显著差法(least significant difference, LSD)检验显著性, 显著性水平均为0.05, 用Origin 8.0作图。
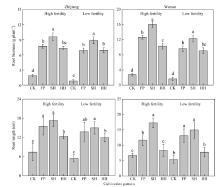
枝江试点油菜各生育期均晚于武穴试点(表3)。按表3所示日期测定各期光照拦截率, 如图1所示。各模式下, 随着生育时期的推进, 光照拦截率均呈先下降后升高再降低的趋势, 在CK模式下尤为明显。不同栽培模式下, 油菜各生育期光照拦截率差异显著, 2个试验点均表现为SH> HH> FP> CK, 且对终花期拦截率的影响最大。以武穴试验点为例, 高产田和低产田终花期光照拦截率均在SH模式下最高, 较FP模式分别高出12.53%和10.74%。光照拦截率在不同地力水平间亦存在差异, 2个试验点均表现为高产田> 低产田。由图 1 还可看出, 在结实中期至成熟期, HH栽培模式下的油菜光照拦截率降低速率最低, 2个试验点在不同土壤肥力条件下的表现一致。
 | 图1 不同栽培模式下油菜关键生育时期光照拦截率差异Fig. 1 Light interception rate of canola under different cultivation modes CK、FP、SH和HH分别表示零氮对照区、农民习惯栽培模式、超高产栽培模式和高产高效栽培模式; EAF、ENF、MOM和MAT分别表示初花期、终花期、结实中期和成熟期; * * 表示在0.01水平差异显著。 CK, FP, SH, and HH represent no nitrogen, farmer pattern, super high yield pattern and high yield and high efficiency pattern; EAF, ENF, MOM, and MAT represent early flowering, end flowering, middle of pod filling and maturity, respectively; * * represents significant difference at the 0.01 probability level. |
从表4看出, 不同生育时期, 油菜光能利用率不同, 表现为初花期至终花期最低, 终花期至结实中期最高, 而结实中期至成熟期次之。不同栽培模式下, 油菜光能利用率差异显著, 2个试验点高、低
产田均表现为SH> HH> FP> CK; 干物质累积量在不同栽培模式下的变化趋势与光能利用率变化趋势基本一致(表5)。方差分析表明, 不同栽培模式对终花期至结实中期的RUE和干物质累积量影响最大, 且地力水平和栽培模式存在极显著的互作效应。以终花期至结实中期为例, 与FP模式相比, SH栽培模式下, 光能利用率在武穴试验点低产田和高产田分别高出6.38%和2.00%, 在枝江试验点则分别高出7.19%和8.02%; 干物质累积量在武穴试验点低产田和高产田分别高出19.39%和24.82%, 在枝江试验
点则分别高出13.67%和11.65%; 与FP模式相比, HH栽培模式下, 光能利用率在武穴试验点低产田和高产田分别高出2.13%和8.67%, 在枝江试验点低产田变化不大, 高产田高出6.17%; 干物质累积量在武穴试验点低产田和高产田分别高出15.66%和21.58%, 在枝江试验点低产田和高产田分别高出7.04%和6.02%。
| 表4 不同栽培模式下各关键生育时期油菜光能利用率 Table 4 Effect of cultivation mode on radiation use efficiency of canola |
| 表5 不同栽培模式下各关键生育时期油菜干物质累积 Table 5 Effect of cultivation mode on biomass accumulation of canola |
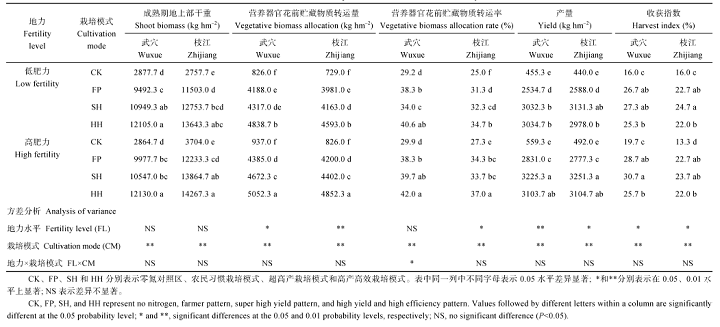
不同肥力地块干物质生产能力和产量差异显著, 高肥力地块成熟期地上部干重、营养器官花前贮藏物质转运量和产量均高于低肥力地块。上述各指标同时受到不同栽培模式影响。不施肥田块干物质生产和产量显著低于其他3种栽培模式。成熟期地上部干重、营养器官花前贮藏物质转运量和转运率均表现为HH> SH> FP> CK。而产量和收获指数在SH表现最高, 与SH相比, HH产量略低, 但差异不显著, 而收获指数在低肥力田块则显著降低(表6)。
| 表6 不同栽培模式对油菜花前干物质转运量和产量相关指标的影响 Table 6 Effects of cultivation modes on pre-anthesis biomass allocation and yield of canola |
由图2可知, 不施氮肥模式下, 根系生长受到严重抑制, 干物质累积量最低; 其他模式下则表现为SH> FP> HH。与FP相比, SH模式下, 枝江试验点单株根系干重在高肥力田和低肥力田分别增加29.35%、29.35%, 在武穴试验点分别增加26.96%和28.13%。与FP相比, HH模式下, 2个试点不同肥力条件下整体表现为降低的趋势, 降幅为4.62%~ 18.51% (图2-A)。不同栽培模式下油菜根长变化趋势与根干重基本一致(图2-B)。
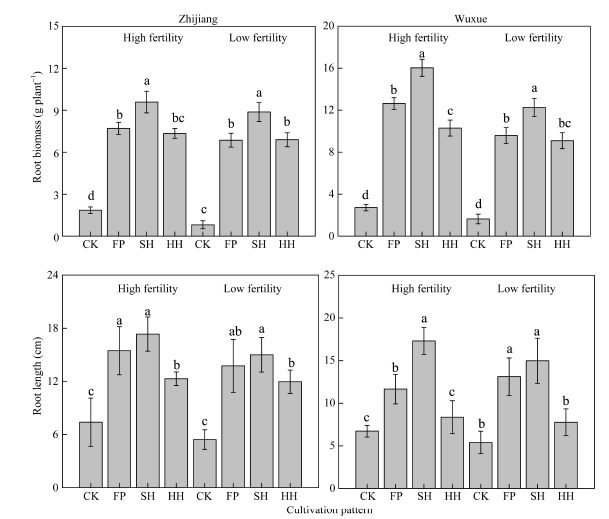 | 图2 不同栽培模式对油菜成熟期根系干重(A)和主根长(B)的影响Fig. 2 Root biomass of canola under different cultivation modes CK、FP、SH和HH分别表示零氮对照区、农民习惯栽培模式、超高产栽培模式和高产高效栽培模式。 CK, FP, SH, and HH represent no nitrogen, farmer pattern, super high yield pattern, and high yield and high efficiency pattern. |
油菜田间倒伏受栽培模式影响显著。如图3所示, 油菜不施氮肥条件下根颈最细; 与FP相比, SH模式下, 油菜根颈粗增加, 枝江高肥力和低肥力田分别增加了12.78%和8.81%, 武穴高肥力和低肥力田分别增加了22.53%和21.09%; 与FP相比, HH模式下, 油菜根颈粗整体上有降低的趋势。如表7所示, 油菜株高和抗折力在不施氮条件下最低。不同栽培模式下, 株高、根冠比和抗折力均表现为SH> HH> FP。武穴试验点未发生倒伏, 因此以枝江试验点为例。各栽培模式中以不施氮处理倒伏最严重, 其次为SH模式, HH模式下倒伏最轻。与FP相比, SH模式下, 油菜在高肥力田和低肥力田倒伏角度分别增加34.29%和16.20%; HH模式下, 油菜在高肥力田和低肥力田倒伏角度分别减小1.36%和24.19%。倒伏指数与倒伏角度的变化趋势较为一致, 在不同地力条件下均表现为CK> SH> FP> HH。
 | 图3 不同栽培模式对油菜根颈粗的影响Fig. 3 Root crown diameter of canola under different cultivation modes CK、FP、SH和HH分别表示零氮对照区、农民习惯栽培模式、超高产栽培模式和高产高效栽培模式。 CK, FP, SH, and HH represent no nitrogen, farmer pattern, super high yield pattern, and high yield and high efficiency pattern. |
| 表7 不同栽培模式对油菜倒伏相关性状的影响 Table 7 Lodging related indices of canola under different cultivation modes |
施氮量和密度是调控产量和抗倒性的重要栽培措施, 可调节群体结构和光合作用, 影响植株茎秆与生殖器官间碳水化合物的积累与运转及茎秆结构形态的建成, 进一步影响茎秆机械强度等抗倒伏关键指标及最终的籽粒产量[20]。产量和抗倒性是两个相互制约的因子。在一定范围内, 增施氮肥或增加
种植密度, 最终产量虽增加, 但是, 由于冠层重量增加, 增加了后期倒伏的风险。可见, 为提高油菜机械生产效率, 同时提高产量和抗倒性, 需要将上述栽培因素合理整合, 充分发挥其正向效应, 减弱其负向效应, 以协同提高光能的截获利用、光合产物生产及其转运分配效率, 协调高产和倒伏间的矛盾。
油菜产量和抗倒性均以干物质累积为基础。干物质是光合作用的产物, 与作物产量密切相关[21]。增施氮肥, 影响了群体光合特性[22], 光合器官(叶片与角果)发育较好, RUE提高, 产量增加; 增加种植密度后, 株距变小, 油菜个体之间对光照养分等的竞争激烈, 个体发育受限制, 但单位面积株数增多, 群体数量的优势弥补了单株生产力的不足: 密植条件下, 油菜单株产量降低, 但群体增产[23]; 此外, 种植密度可通过改变作物株型及冠层结构, 影响光能拦截与利用, 最终RUE也增加[24]。因此, 随密度增加, 产量显著增加。群体光能截获及RUE是干物质形成的关键[25], 可见, 增施氮肥或增加种植密度, 均可通过增加群体光能截获和RUE的提高, 促进光合产物积累, 最终达到增产的效果[26]。与传统农民习惯(FP)模式相比, 超高产(SH)模式增加了种植密度和氮肥投入, 同时基肥比例减少, 薹肥比例增加。SH模式下油菜全生育期光能截获量和光能利用效率显著提高, 有利于干物质累积。作物不同生育时期的RUE不同[27], 对氮肥、密度的响应也存在差异。初花至终花期, 是营养生长与生殖生长竞争激烈的时期, 此时期增加RUE, 有利于缓解营养生长和生殖生长间的矛盾, 为油菜高产抗倒打下基础。本试验中, 不同栽培措施对终花期至结实中期的光能利用率影响最大, 此时正是油菜产量形成关键时期, 可显著影响产量。根系形态决定植株获得水分和养分的能力, 因此根系是影响产量的重要器官, 且受氮肥、密度影响显著[28]。反映根系状况的指标包括根长、根表面积、平均根直径等[29]。一定范围内增施氮肥使根长增加[30], 随种植密度增加, 玉米根长增加[31], 大豆根长降低[28]。本试验条件下, SH模式促进了根系生长, 根系长度、根颈粗和干物质累积量均增加, 有利于植株吸收土壤水分和矿质营养[32], 最终整株干物质积累量显著增加, 收获指数增加, 籽粒产量显著提高。但是, SH栽培模式下, 由于用肥量增加过多, 株高增加, 茎秆密度实则降低, 抗折力因此降低, 再加上冠层重量增加, 导致在3种栽培模式下倒伏指数最高, 倒伏最严重, 降低了油菜机械收获效益。
与肥料投入量偏高的SH模式相比, HH模式适当减少了氮肥用量和施肥次数, 并进一步提高种植密度, 因此个体间竞争加剧, 单株根系生长受阻, 表现为根长、根系干物质累积量降低, 结合SH和FP之间根系形态的差异, 说明油菜根系生长对密度的响应存在一个阈值, 在该值之前, 根系长度和根系干物质累积随密度的增加而增加, 即在SH模式下, 根系生长较好, 超过该值之后, 根系长度和干物质累积随密度的增加而降低, 这与王树丽等[31]研究结果一致。但是从群体角度来讲, 高密度种植增加了根层群体结构, 从而对土壤养分的吸收利用能力更强[33], 一定程度上缓解了个体间的养分竞争, 增加了群体产量。HH模式下, 花前光能截获量和利用效率虽然低于SH模式, 导致花前干物质积累较少, 收获指数低, 但由于薹肥施用量增加, 花后叶片衰老延缓, 成熟后期光能截获量降低速率低于SH模式, 从而促进了花后干物质积累, 群体干物质量增加, 在一定程度上缩小了两种模式间的产量差异, 使HH模式的产量仅略低于 SH模式, 但不显著。此外, HH模式下, 油菜株高低于SH模式, 茎秆抗折力最大, 且由于高密条件下单株产量降低, 冠层重量减轻, 倒伏角度、倒伏指数最小, 倒伏最轻。与SH模式相比, HH模式适当减少了氮肥投入和施肥次数, 但能获得相同水平的产量, 同时减轻了倒伏风险, 提高了机械收获效率。
与当地农民习惯栽培模式相比, 超高产栽培模式和高产高效栽培模式增加了种植密度和施氮量, 提高了各时期的光能截获量和光能利用效率, 促进了干物质的积累, 为增产和抗倒奠定了基础。超高产栽培模式和高产高效栽培模式的籽粒产量均显著高于当地农民习惯栽培模式和不施氮肥模式。高产高效栽培模式增加了种植密度, 减少了施氮量和施肥次数, 其籽粒产量虽然较超高产栽培模式略有降低, 但不显著, 因氮肥投入量少, 施肥次数减少, 节约了生产成本和劳动力, 后期倒伏也显著降低, 起到了"以密抗倒、以密省肥"的效果, 机械收获效益显著提高, 实现了高产高效栽培。综合考虑籽粒产量和抗倒性, 以"减肥、增密"为技术特点的高产高效模式为适应油菜机械收获的推荐栽培模式。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|