{kind=link}
白芝麻籽粒油脂、蛋白质及芝麻素含量QTL定位分析
[吴坤1, 2 , 吴文雄1 , 杨敏敏1 , 刘红艳1 , 郝国存1 , 赵应忠1, *  ]
]
]
|
|


第一作者联系方式: E-mail: adwukun@126.com, Tel: 18672996981
芝麻籽粒油脂、蛋白质和芝麻素含量是芝麻品质育种的3个重要目标。为了解析其遗传机制并检测相关QTL, 利用近红外谷物品质分析仪对一个包含224个株系的F9代重组自交系(RIL)群体在2年3个环境下的籽粒品质性状检测, 结果表明, 群体内株系间差异显著且呈典型正态分布, 而同一环境不同重复间表现差异不显著。相关性分析显示, 籽粒含油量与蛋白质含量显著负相关, 籽粒含油量与芝麻素含量显著正相关, 蛋白质含量与芝麻素含量显著负相关; 利用该RIL群体已构建的高密度遗传图谱, 采用基于混合线性模型的复合区间作图法(MCIM)检测到8个QTL, 表型贡献率为0.41%~14.55%; 采用多重区间作图法(MIM)检测到13个QTL, 可解释5.2%~18.6%的表型变异。其中5个主效QTL被2种方法同时检测到且定位区间相同, 2个主效QTL在2个或3个环境中被重复检测到。控制含油量的 Qoc-5与芝麻素含量的 Qsc-5位于LG5连锁群上的相同区段, 加性效应均为正值; 而控制蛋白质含量的 Qpc-5也位于相邻位置, 但加性效应为负值。LG2和LG1连锁群上也存在相似情况, 反映品质性状相关QTL之间存在一因多效或紧密连锁。因此, 在芝麻品质育种中选择高含油量可以兼顾高芝麻素, 但应对蛋白质含量进行负向选择。
Seed oil content, protein content and sesamin content are three important targets for sesame breeding. The objective of this study was to explore the genetic model and detect quantitative trait loci (QTLs) for three quality traits using a mapping population containing 224 recombinant inbred lines (RILs, F9). The three quality traits of seeds were measured by near infrared reflectance spectroscopy in three environments. Correlation analysis showed oil content was negatively correlated with protein content but positively correlated with sesamin content, whereas, protein content was negatively correlated with sesamin content. Using mixed composite interval mapping (MCIM) method, eight QTLs for the three seed quality traits were detected with heritability of additive effect ranging from 0.41% to 14.55%. Using multiple interval mapping (MIM) method, 13 QTLs were mapped on nine linkage groups (LGs), and a single QTL explained 5.2%-18.6% of the total phenotypic variation. Among them, five loci had at least one marker identical to the QTL identified with MCIM, indicating they might be located in the same region. Two QTLs were detected in multiple environments. QTLs Qoc-5 and Qsc-5 were located in the same region on LG5, both had positive effects, whereas Qpc-5 in the adjacent region had a negative effect. Similar loci exist on LG2 and LG1, probably caused by pleiotropism or close linkage of several QTLs. These results suggest that the oil and sesamin contents in seed can be improved together, while the protein content needs to be selected negatively with the oil or sesamin content.
芝麻(Sesamum indicum L.)属胡麻科胡麻属, 是世界上古老的油料作物之一。芝麻中富含脂肪及蛋白质, 并含有芝麻木酚素(lignan)等多种生物活性物质。芝麻油是一种品质好、营养价值高的优良食用油; 芝麻蛋白质含量对食品用原料及榨油后饲料用芝麻饼的品质影响十分关键; 芝麻素(sesamin)是芝麻木酚素的重要组成成分, 具有重要营养和药用价值, 已成功用于抗氧化功能食品开发。因此, 提高芝麻种子的含油量、蛋白质含量和芝麻素含量, 对进一步提高芝麻生产效益有着非常重要的意义。
与产量等复杂农艺性状相似, 含油量、蛋白质含量、芝麻素含量等品质性状也是典型的受多基因控制的数量性状。利用分子标记技术开展数量性状位点(quantitative trait locus, QTL)遗传定位及分子标记辅助育种被证明是解决产量品质等复杂性状遗传改良的有效手段[1, 2]。在油料作物种子含油量QTL定位研究方面, Yan等[3]在3个环境中检测到与油菜含油量相关的11个QTL。金梦阳等[4]利用黄籽油菜与黑籽油菜杂交后代重组自交系(recombinant inbred lines, RIL)群体定位获得了7个与含油量相关QTL, 单个位点的遗传贡献率在3.73%~10.46%之间。Zhang等[5]利用184个株系的大豆重组自交系群体定位到一个位于Dl连锁群的控制含油量的主效QTL。郑永战等[6]利用114个株系的BC1F1群体, 定位了9个大豆脂肪及脂肪酸组分相关QTL, 但主效QTL数量不多。廖伯寿等[7]利用远杂9102与中花5号构建花生重组自交系群体, 获得了3份含油量高达60%以上的超亲本重组自交系后代, 也检测到多个QTL, 但大多效应值较小。相对于其他油料作物, 芝麻含油量高, 营养价值丰富, 种子含油量为45%~54%, 蛋白质含量为22%~25%[8]。研究还发现, 芝麻种子油脂、蛋白质和芝麻素含量之间存在一定的相关性。梅鸿献等[9]对我国209份芝麻种质资源的芝麻素、脂肪和蛋白质含量分析, 表明芝麻素与脂肪含量呈显著正相关, 与蛋白质含量相关不显著, 芝麻素含量比蛋白质、脂肪含量变异范围更大, 遗传多样性更丰富。张秀荣等[10]和Wang等[11]分别对我国芝麻育成品种及核心种质资源的品质成分分析发现, 芝麻素与芝麻酚林含量之间呈显著正相关。金青哲等[12]发现不同粒色芝麻的芝麻素含量存在显著差异, 芝麻素平均含量以白芝麻最高, 棕黄色芝麻稍逊, 黑芝麻次之。危文亮等[13]通过核心种质群体的关联分析获得了与芝麻种子含油量性状极显著相关的8个
分子标记。王蕾等[14]通过该种质资源核心群体关联分析获得了1个与芝麻素含量显著相关的SSR标记。目前, 芝麻中还没有利用高密度遗传连锁图谱开展种子油脂、芝麻素和蛋白质含量等品质性状相关QTL定位的研究报道。课题组前期利用品质等农艺性状差异显著的白芝麻育成品种中芝14和白芝麻地方品种“ 庙前芝麻” 构建了一个包含224个株系的白芝麻重组自交系群体, 通过群体的简化基因组测序构建了一张包含1230个标记的高密度遗传图谱, 并且获得了13个单株蒴果数、每蒴籽粒数、千粒重等芝麻籽粒产量性状相关QTL[15]。本研究拟在上述高密度遗传连锁图谱基础上, 通过该RIL群体多年多点的籽粒油脂、蛋白质和芝麻素含量测定, 解析并验证三者之间的相互关系, 同时开展上述品质性状的QTL定位研究, 获得芝麻品质性状相关分子标记, 为芝麻高油、高蛋白质及高芝麻素种质资源的发掘及品质育种提供重要理论依据和技术支持。
以优质白芝麻育成品种中芝14 [含油量(54.29± 0.55)%, 蛋白质含量(22.06± 0.65)%, 芝麻素含量(0.307± 0.037)%]和白芝麻地方品种庙前芝麻[含油量(50.27± 0.32)%, 蛋白质含量(24.95± 0.35)%, 芝麻素含量(0.233± 0.020)%]杂交获得F1, 自交后获得F2群体, 再通过单粒传法连续自交8代, 获得高世代重组自交群体, 该群体包含224个稳定纯合株系, 被用于高密度遗传图谱构建及品质性状相关QTL定位研究。
2012年6月在中国农业科学院油料作物研究所武昌徐东(2012XD)试验基地, 2013年6月继续在徐东(2013XD)试验基地及武汉阳逻(2013YL)试验基地设置随机区组试验, 每个小区2行, 行长2 m, 行距0.4 m, 每行13株, 重复3次。按照长江流域夏播芝麻正常栽培管理方式进行田间管理, 每小区分单株收获中间6株健康成熟植株, 晒干、脱粒并整净后, 将籽粒充分晒干、混匀、称取10 g用于检测品质性状。
利用近红外谷物品质分析仪(FOSS Infratec 1241)测定RIL群体各株系种子含油量(oil content, OC)、蛋白质(protein content, PC)和芝麻素(sesamin content, SC)含量, 每份样品重复测定3次取平均值。芝麻种子品质分析模型由福斯华(北京)科贸有限公司根据我们提供的112份白芝麻种质资源品质标准化学测定数据(测定脂肪含量采用国家标准GB/T 14772-2008; 测定蛋白质含量采用国家标准GB/T 5009.5-2010; 测定芝麻素采用高效液相色谱法)及近红外品质分析光谱数据构建(未发表)。
3个试验中亲本及RIL群体各株系3个重复的平均值用于计算频次分布、标准误差、Pearson’ s相关系数和ANOVA方差分析(SAS statistics package[16])。利用公式H2=σ g2/(σ g2 + σ e2/r)计算广义遗传率(H2), 其中σ g2代表遗传方差, σ e2代表残差, r代表各基因型的重复数。利用软件Windows QTL Cartographer 2.5中的多重区间作图法(multiple interval mapping, MIM)[17], 以及软件QTL Network 2.1中的基于混合线性模型的复合区间作图法(mixed composite interval mapping, MCIM)[18], 分别对含油量、蛋白质含量及芝麻素含量进行不同环境的QTL扫描。MCIM模型通过对群体各性状不同环境及重复同时进行扫描, 通过F阈值(α = 0.01)进行1000次的排列在全基因组范围内QTL检测, 并通过Monte Carlo Markov Chain (MCMC)算法以Gibbs样本大小为20 000估计QTL效应, 同时计算各QTL的加性效应遗传率ha2。MIM模型通过对群体在3个环境中各性状3个重复的平均值分别进行扫描, 利用排列测验法(permutation test)来估算不同环境中各个性状的LOD显著性阈值, 选取参数1000次回归, 在显著水平0.01来判断是否存在QTL, 同时给出相关的加性效应、显性效应及基因位点对性状表型方差的贡献率(R2)。
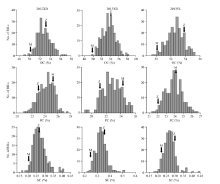
RIL群体在3个环境下的油脂、蛋白质及芝麻素含量均表现正态分布或偏正态分布(图1), 群体平均值都比较稳定, 变幅范围也都表现双向超亲分离, 偏度和峰度大多小于1 (表1和图1), 表明油脂、蛋白质及芝麻素含量是典型数量性状, 受多基因控制。并且同一环境不同重复间表现差异不显著, 该群体3个环境检测的品质性状数据符合QTL定位要求。而3个环境下芝麻素含量峰度都大于1, 说明该性状受环境影响较大。含油量、蛋白质含量和芝麻素含量的广义遗传率(H2)分别为40.38%、22.76%和14.68% (表3)。
| 表1 RIL群体3个品质性状在3个环境下的表型分布特征 Table 1 Phenotypic variation of the three quality traits of RIL population in three trials |
含油量、蛋白质含量及芝麻素含量的相关性均达到极显著水平(表2)。其中3个环境下含油量与蛋白质含量之间均存在显著负相关, 含油量和芝麻素含量之间均存在显著正相关, 蛋白质含量与芝麻素含量之间均存在显著负相关。
| 表2 3个环境下3个品质性状的相关系数 Table 2 Correlation coefficients of the three quality traits in three environment trials |
2.3.1 含油量相关QTL定位 利用QTL Network 2.1中MCIM法检测到1个含油量相关QTL (表3), Qoc-5位于LG5连锁群上, 该QTL加性效应遗传率为3.47%, 其加性效应在P≤ 0.001水平表现极显著, 且为正值, 表明来自父本中芝14的等位变异可以显著提高籽粒含油量。利用WinQTLcart 2.5中MIM法检测到5个含油量相关QTL, 分布在LG1、LG2、LG5、LG9和LG16连锁群上, 且均只在1个环境中被检测到(表4), 并且仅有LG5连锁群上的Qoc-5同时用MCIM和MIM方法被检测到。其中LG1连锁群上的Qoc-1可以解释10.1%的表型变异; LG5连锁群上的Qoc-5可以解释7.7%的表型变异; 其余3个QTL均可以解释6.5%以上的表型变异。上述QTL中, Qoc-2和Qoc-5的加性效应为正值, 而Qoc-1、Qoc-9和Qoc-16的加性效应为负值, 表明该RIL群体的亲本中芝14和庙前芝麻中均存在能够提高籽粒含油量的等位变异。
| 表3 利用MCIM模型同时检测3个环境下RIL群体品质性状相关QTL Table 3 QTL for quality traits detected by MCIM using the RILs in multi-trials |
2.3.2 蛋白质含量相关QTL定位 用MCIM法检测到2个蛋白质含量相关QTL, Qpc-2和Qpc-5分别位于LG2和LG5连锁群上(表3), 两者的加性效应遗传率分别为4.26%和3.31%。2个QTL的加性效应均为负值且达到极显著水平, 表明来自庙前芝麻的等位变异可以显著提高籽粒蛋白质含量。用MIM法检测到5个QTL, 分布位于LG1、LG2、LG3、LG5和LG6连锁群上(表4), 且均只在1个环境中被检测到。同时用MCIM和MIM方法检测到LG2和LG5连锁群上的Qpc-2和Qpc-5, 其中Qpc-2可以解释6.2%的表型变异, Qpc-5可以解释10.9%的表型变异; 其余3个QTL均可以解释5.7%以上的表型变异。并且仅有位于LG1连锁群上Qpc-1的加性效应为正值, 其余4个QTL的加性效应均为负值, 表明中芝14和庙前芝麻中均存在能够提高籽粒蛋白质含量的等位变异。
 | 图1 RIL群体3个品质性状在3个环境下的频率分布(OC、PC和SC分别代表油分、蛋白质和芝麻素含量, 箭头所指为各环境中亲本测定值, Z和M分别代表亲本中芝14和庙前芝麻。)Fig. 1 Frequency distribution of three quality traits of RIL population in three environments(OC: oil content; PC: protein content; SC: sesamin content. Arrows indicate determined values of two parents under environments. Z and M stand for Zhongzhi 14 and Miaoqianzhima, respectively.) |
2.3.3 芝麻素含量相关QTL定位 用MCIM法检测到5个芝麻素含量相关QTL (Qsc-5、Qsc-6、Qsc-8、Qsc-11和Qsc-16), 分别位于LG5、LG6、LG8、LG11和LG16连锁群上(表3), 它们的加性效应遗传率分别为14.55%、2.01%、5.05%、0.41%和3.34%。5个QTL的加性效应均达到极显著水平, 其中仅有Qsc-16的加性效应为负值, 其余均为正值, 表明中芝14和庙前芝麻中均存在能够提高籽粒芝麻素含量的等位变异。Qsc-6和Qsc-11间存在加性上位效应, 该效应在P≤ 0.05水平表现显著且为正值, 该加性上位效应遗传率为0.79%。用MIM法检测到3个QTL (Qsc-4、Qsc-5和Qsc-8), 分别位于LG4、LG5和LG8连锁群上, 且加性效应均为正值(表4)。LG4连锁群上的Qsc-4可以解释5.4%的表型变异, 仅在2013年阳逻环境中被检测到; LG5连锁群上Qsc-5在2012年徐东、2013年徐东和2013年阳逻3个环境中均被检测到, 且位置非常相近, 分别可以解释11.1%、18.6%和14.1%的表型变异; LG8连锁群上Qsc-8在2013年徐东和2013年阳逻两个环境中被检测到, 分别可以解释8.9%和5.2%的表型变异。Qsc-5和Qsc-8用MCIM和MIM扫描均被成功检测到, 且对表型贡献率均较大, 为控制芝麻素含量主效QTL。
| 表4 利用MIM模型分别检测3个环境下RIL群体品质性状相关QTL Table 4 QTL for quality traits detected by MIM using the RILs in three environment traits |
2.3.4 不同籽粒品质相关QTL之间关系 采用QTL Network 2.1中MCIM法检测到分布在6个连锁群上的8个籽粒含油量、蛋白质含量及芝麻素含量相关QTL, 其加性效应遗传率为0.41%~14.55% (表3)。其中Qoc-5与Qsc-5位于连锁群LG5上相同区间且位置非常相近, 且加性效应均为正值, 表明此位点能够同时对芝麻籽粒含油量和芝麻素含量产生相同作用。与之相反, 同样位于LG5上的控制籽粒蛋白质含量的Qpc-5与上述2个QTL的遗传位置比较近, 但其加性效应为负值, 也即增效等位变异来源于另一个亲本庙前芝麻。
采用WinQTLcart 2.5中MIM法共检测到分布在9个连锁群上13个籽粒含油量、蛋白质含量和芝麻素含量相关QTL, 可以解释5.2%~18.6%的表型变异(表4)。其中LG2连锁群上的Qoc-2与Qpc-2所在遗传区间完全相同, 两者对含油量和蛋白质含量的表型贡献率也较大, 但产生的加性效应却完全相反, 前者增效等位变异来自中芝14, 后者增效等位变异来自庙前芝麻。与此相似的还有Qoc-5、Qsc-5和Qpc-5, 三者遗传位置更加近, 含油量和芝麻素含量增效等位变异来源于亲本中芝14, 而蛋白质含量增效等位变异来源于庙前芝麻。更有意思的是, 另外1个含油量相关Qoc-1与1个蛋白质含量Qpc-1位置比较近, 分别对含油量及蛋白质含量的表型贡献率也很大, 两者产生的加性效应也完全相反, 前者增效等位变异来源于庙前芝麻, 后者增效等位变异来源于中芝14, 与LG2及LG5连锁群上的现象完全相反。
利用MCIM与MIM方法检测得到16个籽粒品质相关QTL, 其中被两种方法同时检测到且定位遗传区间相同的有5个(Qoc-5、Qpc-2、Qpc-5、Qsc-5和Qsc-8), 它们的加性效应值具有正负一致性。在多环境中同时被检测到的QTL有Qsc-5和Qsc-8, 分别在3个和2个环境中被检测到。被两种方法检测到定位在重叠遗传区间内但控制目标性状不同的QTL有2组(Qpc-6与Qsc-6; Qoc-16与Qsc-16), 其中Qpc-6与Qsc-6加性效应值一负一正, 表明该QTL来源于中芝14的等位变异, 对蛋白质含量表现增效, 对芝麻素含量表现减效; 而Qoc-16与Qsc-16的加性效应值均为负值, 表明该QTL增效等位变异均来源于另一个亲本庙前芝麻。另外7个没有被不同方法或者在不同环境中重复检测到的QTL可能有待于进一步验证。采用MCIM模型未检测到上述QTL的加性效应与环境的互作效应达到显著水平。
关于QTL检测假阳性的问题, 一般可通过界定适当的LOD阈值来克服[19]; 而且大多数植物数量性状由若干主效基因和微效基因共同控制, 进行QTL扫描定位时, 最好通过多个环境以及两种以上的检测方法比较分析, 以确定主效QTL和可能存在的微效QTL。本文采用Windows QTL Cartographer 2.5软件中多重区间作图法MIM法时通过1000次排列测验, 确定LOD阈值为3.0, 按照2年3个环境逐一扫描, 检测到13个QTL, 获得各QTL的加性效应及表型变异贡献率R2; 采用QTL Network 2.1软件中基于混合线性模型的复合区间作图法MCIM对2年3个环境品质数据同时扫描, 检测到8个QTL, 获得各QTL加性效应及遗传率ha2。2种方法计算方式不同, 但都被国内外学者广泛接受和认同, 本研究中有5个QTL被两种方法同时检测并定位到相同区间。通过2年3个环境下的随机区组重复和两种模型方法重复检测, 比较不同环境以及不同检测方法的分析结果, 可以增加检测QTL定位结果的可靠性。
Kamal等[20]研究印度芝麻, 发现芝麻油中芝麻素含量为0.13%~0.59%; 金青哲等[21]分析国内重点产区15个芝麻品种的芝麻素含量, 表明芝麻油中芝麻素的含量为0.12%~0.78%, 均与本研究中RIL群体芝麻素含量变异范围0.17%~0.51%相近。梅鸿献等[9]发现芝麻素与脂肪含量呈显著正相关, 芝麻素与蛋白质含量相关不显著。金青哲等[21]认为芝麻素含量与芝麻含油率呈正相关(相关系数0.556), 芝麻素平均含量以白芝麻最高, 黑芝麻次之。本研究表明, 3个环境下含油量和芝麻素含量显著正相关, 含油量与蛋白质含量显著负相关, 蛋白质与芝麻素含量显著负相关, 表明芝麻含油量可与芝麻素含量同步提高, 降低蛋白质含量有利于含油量和芝麻素含量的提高。
本研究利用MIM方法检测到5个与含油量相关QTL, 累计可以解释40.5%的表型变异; 5个蛋白质含量相关QTL累计可以解释36.8%的表型变异; 3个芝麻素含量相关QTL, 有2个在不同环境中可被同时检测到, 其中Qsc-5可以解释11.1%~18.6%的表型变异。危文亮等[13]通过216份芝麻核心品种含油量关联分析, 也仅在2个年份环境中重复检测到8个分子标记(其中SSR标记4个), 总的变异解释率为28.46%和38.01%, 变异解释率变幅为2.95%~6.03%; 另外15个分子标记只在一个年份环境中检测到, 变异解释率变幅为3.06%~5.18%。王蕾等[14]同样利用该芝麻核心品种关联分析群体, 在一个环境中利用MLM和GLM两个模型一致检测到4个芝麻素含量显著相关分子标记, 其中一个极显著相关SSR标记SSI182-3在两个模型中可以解释相对较高的芝麻素表型变异(分别达到6.84%和6.36%)。但由于上述研究中涉及SSR标记大都在亲本中芝14和庙前芝麻之间没有多态, 上述显著相关SSR标记均未能被成功定位到本研究涉及的遗传图谱上[15], 因此无法比较它们与本研究QTL之间的关系。这些研究也都表明, 芝麻素含量至少受1个主效基因(Qsc-5)控制, 并在一定程度上受多个微效基因修饰; 而含油量和蛋白质含量受多个微效基因控制。
综合比较QTL定位结果与性状相关性可以发现: (1)控制含油量的Qoc-5与芝麻素含量的Qsc-5位于LG5连锁群上的相同区段, 加性效应均为正值, 而控制蛋白质含量的Qpc-5也位于该连锁群的相邻位置, 但加性效应为负值; 同样LG2连锁群上的Qoc-2与Qpc-2、LG1连锁群上的Qoc-1与Qpc-1以及LG6连锁群上的Qpc-6与Qsc-6也存在遗传位置相近或交叠而加性效应一正一负的情况, 这些从分子遗传机制上解释了上述品质性状相关性分析的结果, 也反映3个品质性状相关QTL存在一因多效或紧密连锁的关系。(2)多数含油量相关QTL (Qoc-2和Qoc-5)增效等位变异来自中芝14, 多数蛋白质含量相关QTL (Qpc-2、Qpc-3、Qpc-5和Qpc-6)增效等位变异来源于庙前芝麻, 多数芝麻素含量相关QTL (Qsc-4、Qsc-5、Qsc-8和Qsc-11)增效等位变异来源于中芝14。(3)与大多数情况相反的是, 3个控制含油量的Qoc-1、Qoc-9和Qoc-16增效等位变异来源于庙前芝麻, 而1个控制蛋白质含量的Qpc-1增效等位变异来源于中芝14, 1个控制芝麻素含量的Qsc-16增效等位变异来源于庙前芝麻。这些结果表明, 相对高油亲本中芝14的籽粒含油量仍有较大提升空间, 相对高蛋白亲本庙前芝麻的籽粒蛋白质含量也仍然有提升空间, 该杂交组合后代也一定能够创制筛选出含油量或蛋白质含量超高值亲本的优异株系, 该RIL群体部分株系籽粒品质性状也的确出现明显超亲优势。这些现象或推论都将对芝麻育种实践提供重要参考。
众所周知, 植物种子含油量是由多基因控制的复杂数量性状[22, 23]。马珍珍等[24]通过连续自交8代构建甘蓝型油菜重组自交系群体, 检测表明4个环境下含油量与蛋白质、纤维素和半纤维素之间都存在显著负相关。戴维等[25]研究认为, 油菜种子含油量以加性基因效应为主, 同时受非加性效应的控制。其种子含油量与蛋白质含量显著负相关[26]。禹山林等[27]利用高油花生和低油花生及其杂交组合的F1、F2群体为材料, 对种子含油量的遗传分析表明, 花生种子含油量由两对加性-显性-上位性主基因+加性-显性多基因共同控制, 主基因遗传率为72.55%。而本研究中得到控制含油量、蛋白质及芝麻素含量相关QTL的数量不多, 效应值不大, 还可能存在更多微效基因。在育种实践中, 目前可能只有芝麻素组分能够通过分子辅助转育单个主效基因实现高效品质性状改良, 而对含油量和蛋白质含量改良可能更多需要依靠多基因聚合等手段。
3个环境下含油量和芝麻素含量间存在显著正相关, 含油量与蛋白质含量间存在显著负相关, 蛋白质与芝麻素含量之间存在显著负相关。检测得到16个籽粒品质相关QTL, 其中5个被两种方法同时检测到且定位遗传区间相同, 2个在多环境中同时被检测到, 包括1个控制芝麻素含量的主效QTL (Qsc-5)。在LG2和LG5上含油量相关QTL与芝麻素含量相关QTL紧密连锁, 且加性效应值均为正值, 而相同或相近位置上的蛋白质含量相关QTL加性效应值为负值, 说明油脂合成与芝麻素合成具有同向效应, 而蛋白质合成与油脂及芝麻素合成具有反向效应。因此, 在芝麻品质育种实践中选择高含油量可以兼顾高芝麻素, 但同时应对蛋白质含量进行负向选择。在利用主效QTL的同时, 也应充分考虑利用微效QTL相关分子标记开展不同等位变异的叠加利用。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|