



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻紫鞘染色体片段代换系Z519的鉴定及 PSH1候选基因分析
[周可1, *  , 李燕
, 李燕1, 2, * , 王世明1 , 崔国庆1 , 杨正林1 , 何光华1 , 凌英华1 , 赵芳明1, * ]
, 李燕]
|
|


第一作者联系方式: 周可, E-mail: 960968356@qq.com, Tel: 023-68250486 **同等贡献(Contributed equally to this work)
花青素是广受人们喜爱的植物色素, 在食品加工、杂种纯度鉴定中具有重要的作用。本研究鉴定了一个以日本晴为受体、优良紫鞘恢复系R225为供体亲本的紫鞘水稻染色体片段代换系Z519。Z519共含有16个代换片段, 分布于除第10染色体外的其他11条染色体, 平均长度为6.85 Mb。Z519在芽鞘3 mm时鞘尖呈现紫色, 其后在叶鞘、叶缘、茎维管束和柱头等部位出现紫色线条, 而日本晴各部位均为绿色。Z519叶鞘中花青素含量极显著高于日本晴, 剑叶中没有显著差异。与受体日本晴相比, Z519的株高显著降低, 千粒重、主穗总粒数和实粒数显著增加, 有效穗数、主穗长和结实率无显著差异。进一步以日本晴与Z519杂交产生的F1和F2群体对紫鞘基因进行了遗传分析和分子定位。该紫鞘表型受显性单基因控制, 位于第1染色体InDel标记L03和SSR标记L01之间37.8 kb的区域, 被命名为 PSH1。对该区间进行候选基因预测和测序, Z519在一个编码质体ATP/ADP转运蛋白的 LOC_Os01g45910基因第一外显子的第238~252碱基的GTG重复区又多插入了GTG 3个碱基, 导致增加了一个甘氨酸。qRT-PCR结果进一步表明其表达量在Z519中明显降低, 初步确定 LOC_Os01g45910是 PSH1的候选基因。该研究为 PSH1调控花青素的分子机制奠定了良好基础。
, LI Yan
Anthocyanins as plant pigments are widely liked by people and play a very important role in food processing and hybrid purity identification. Here, a rice chromosome segment substitution line (CSSL) Z519 with purple sheath was identified deriving from recipient Nipponbare and donor R225. Z519 contained 16 substitution segments with 6.85 Mb of average length, which were distributed on 11 chromosomes of rice except the 10th chromosome. The bud sheath of Z519 began to appear the purple color stripes when it was about 3 mm long. Then the purple stripes displayed on sheaths, leaf margins, vascular bundles of stem and stigmas. While all parts of Nipponbare were green. Anthocyanin content in leaf sheath of Z519 was significantly higher than that of Nipponbare, whereas no significant difference was in flag leaf. Compared with Nipponbare, plant height of Z519 was significantly decreased, spikelets number and grain number of main panicle, and 1000-grain weight of Z519 were significantly increased. There was no significant difference between Z519 and Nipponbare in the other traits such as panicle number, main panicle length and seed-setting rate. Then, F2 population from the cross of Nipponbare and Z519 were used for genetic analysis and gene mapping of the purple sheath. The purple sheath in Z519 was controlled by a single dominant gene, named as PSH1, which was mapped on the chromosome 1 between InDel marker L03 and SSR marker L01 with the physical distance of 37.8 kb. By sequencing and gene-predicting in the region, Z519 had three bases (GTG) insertion in the GTG repeat area of the 238th-252th base in the first exon compared with Nipponbare, which resulted in increasing a Gly amino acid. Furthermore, the expression of LOC_Os01g45910was obviously decreased in Z519 by qRT-PCR analysis. Thus, LOC_Os01g45910was preliminary identified as the candidate gene of PSH1. The results lay a good foundation for studying molecular mechanisms of regulating anthocyanin by PSH1.
花青素属类黄酮化合物, 是一种天然的食用色素, 广泛存在于植物中, 具有抗氧化、预防心脑血管疾病、抵御低温、抑制肿瘤细胞等生物功能[1]。花青素的生物合成主要通过苯丙氨酸途径[2], 即苯丙氨酸在一系列酶的催化作用下生成过渡产物4-香豆酸CoA和二氢黄酮醇, 最后生成各种花青素类似物[3]。随着分子生物学的不断发展, 已发现大量调控其合成途径所需酶的结构基因, 如CHS、CHI、F3H、F3’ H、F3’ 5’ H、DFR[4, 5, 6, 7]等; 以及编码转录因子的调节基因, 如R2R3-MYB蛋白、MYB家族的BHLH蛋白、WD40蛋白基因家族[8, 9]等。WD40-bHLH- MYB形成的复合体主要影响其下游基因的表达, 直接调控花青素合成早期基因[10]。此外, 花青素的转运与积累在一定程度上也影响着植株颜色的表现, 但其转运过程尚未完全清楚[11, 12]。目前比较公认的有4种蛋白(GST、MRP、MATE和BTL-homologue)可能参与花青苷类物质向液泡的转运, 即花青素在细胞质中可在GST的作用下糖基化, 在植物液泡膜的谷胱甘肽泵作用下运到液泡中。目前已在许多植物中克隆了GST基因, 如玉米中的Bronze2基因和拟南芥中的TT19基因等[13, 14]。花青素在叶鞘、叶片、种皮等部位累积, 可产生有色稻, 据其沉积的浓淡及分布, 在食品改良、分类学和遗传学研究中具有重要的应用价值, 尤其在杂种纯度鉴定, 目的基因瞬时表达, 以及转基因的指示中具有非常重要的作用[15]。
我们以粳稻日本晴为受体、优良紫鞘恢复系R225为供体通过高代回交和分子标记辅助选择, 鉴定了一个紫鞘水稻染色体片段代换系Z519。目前, 在水稻中, 分子定位的紫鞘基因还较少, 如位于第1染色体的PSH1(t)[16]、第6染色体的PSH(t)[17]和OsC1[15]及第11染色体的OsA1[15]。图位克隆的仅有OsC1和OsA1[15]。此外, 2个水稻紫叶的候选基因OsB1和OsB2是以玉米的B基因序列作探针被同源克隆的[18]。这些研究为揭示花青素分子调控途径奠定了良好基础, 但还远远不够, 鉴定新的水稻花青素相关基因, 对深入理解花青素调控网络有重要意义。
本文进一步对该紫鞘染色体片段代换系Z519进行分子鉴定、形态分析、生理生化和重要农艺性状评价; 对该紫鞘性状进行遗传分析、基因定位和候选基因分析。研究结果对该紫鞘基因克隆和花青素调控途径的分子机制研究有重要意义。
水稻染色体代换系片段Z519是以日本晴为受体亲本、西南大学水稻研究所自主选育的优良紫鞘恢复系R225为供体亲本, 经连续回交和自交并通过表型和分子标记选择, 在BC3F7获得的遗传稳定的紫鞘染色体片段代换系。
用于遗传分析和基因定位的材料是以日本晴和Z519杂交产生的F1和F2隐性群体。
2014年3月8日, 将日本晴、Z519和F2种植于重庆西南大学实验基地育苗。4月15日移栽, 各栽亲本30株及F2全部。每行栽10株, 行、株距分别为26.4 cm和16.5 cm, 常规管理。
1.2.1 Z519代换片段鉴定 以绿鞘日本晴和紫鞘R225有多态性的253个多态性标记[19]从BC2F1开始进行分子标记和表型双重选择。2013年在BC3F4的Z120中发现绿鞘株(8株)与紫鞘株(21株)的比例接近1∶ 3, 收获5株紫鞘株, 连续自交, 然后从每个株系取20株进行分子标记辅助选择, 在BC3F7选出了纯合紫鞘染色体代换系片段Z519。在分子标记筛选的过程中, 当某一标记的带型与受体亲本日本晴带型一致时, 则认为该DNA片段来自受体亲本日本晴的基因组; 当某一标记的带型与供体亲本R225带型一致时, 则认为该DNA片段来自供体亲本R225的基因组; 当连续几个标记的带型与供体亲本一致时, 则认为该段为一个代换片段。参照Paterson等[20]方法计算代换片段的长度。
1.2.2 农艺性状分析 成熟后, 分别取日本晴和Z519中间行的10株考查其株高、有效穗数、主穗长、主穗实粒数、主穗总粒数、结实率和千粒重等农艺性状, 然后进行t测验统计分析。
1.2.3 叶绿素及花青素含量测定 在抽穗期分别取日本晴和Z519各10株的剑叶和叶鞘, 按邹琦方法测定叶绿素含量[21]; 按于晓南等方法测定花青素含量[22]。
1.2.4 分子定位 以日本晴/Z519的F2分离群体的276株隐性(绿鞘)株作为定位群体。选用16个代换片段中效果较好的标记对F2群体的30个绿鞘株进行基因连锁分析, 当发现某一标记与紫鞘基因可能连锁时, 用F2群体中的10株绿鞘单株和10株紫鞘单株进一步验证。当证明确实存在连锁时, 扩大群体, 用这一代换片段上的所有标记进行定位。
采用碱煮法[23]提取用于基因定位的F2单株DNA, 采用CTAB法[24]提取亲本DNA。PCR反应体系和10%非变性聚丙烯酰胺凝胶电泳及快速银染方法与向佳等方法相同[23]。将具有日本晴带型的单株记为A, 具有Z519带型的单株记为B, 具有双亲带型的单株记为H。用MapMaker 3.0进行数据分析和作图, 用Kosambi函数将重组率转化为遗传距离。
1.2.5 候选基因预测与测序 在水稻自动释义数据库(RAD)(http://ricegaas.dna.affrc.go.jp/rgadb/)和水稻释义计划数据库(RAP-DB)(http://rapdb.dna.affrc. go.jp/)中, 查询精细定位区间内所有基因信息。与Gramene (http://www.gramene.org/rice_mutant/)中查询的基因信息比对, 结合基因预测功能, 筛选候选基因。
以日本晴和Z519的DNA序列设计引物, 分段扩增候选基因。PCR产物经琼脂糖凝胶电泳分离后切胶回收并连接到pMD19-T载体, 连接产物转化大肠杆菌感受态DH5α , PCR检测筛选阳性克隆子送上海英潍捷基贸易有限公司测序, 然后分析测序结果。
1.2.6 候选基因的qRT-PCR表达分析 按TaKaRa生物工程有限公司试剂盒说明书 提取及纯化日本晴和Z519的叶鞘RNA, 按照逆转录试剂盒说明书制作cDNA。以Actin为内参基因, 910d1引物检测候选基因的表达量。在Bio-Rad CFX96荧光定量PCR仪上完成定量PCR, 反应总体系为25 μ L, 即2 μ L的cDNA模板, 2 μ L 0.4 μ mol L-1的引物, 12.5 μ L 1X的SYBR Green荧光染料和8.5 μ L RNase free ddH2O。用CFX-Manager软件处理数据。
在前期研究的基础上, 用Z519代换片段上的82个SSR标记及代换片段外的36个SSR标记对10株Z519进行进一步代换片段鉴定和遗传背景检测, 发现10个单株的代换片段一致, 且没有检出背景片段, 说明Z519已纯合。Z519共含有来自R225的16个代换片段, 分别位于除第10染色体外的其他11条染色体。第1染色体的代换区间为RM3475- RM6960, 估计长度为9.85 Mb。第2染色体的代换区间为RM2483-RM6843, 估计长度约为17.67 Mb。第3染色体的代换区间为RM132-RM3417和RM6266-RM3346, 估计长度分别为6.30 Mb和11.60 Mb。第4染色体的代换区间RM5900-RM1155, 估计长度为6.85 Mb。第5染色体上代换区间为RM2010-RM405和RM5441-RM4837, 估计长度分别为2.40 Mb和1.85 Mb。第6染色体上的代换片段为RM3330-RM3183-RM7193, 估计长度为4.5 Mb。第7染色体上的代换片段为RM8006-RM3583- RM1180和RM234-RM8261-RM3552, 估计长度分别为0.65 Mb和0.45 Mb。第8染色体的代换区间为RM1376-RM8264, 估计长度为17.05 Mb。第9染色体的代换区间为RM5657-RM1189, 估计长度为2.75 Mb。第11染色体的代换区间为RM3717- RM3747和RM3701-RM2191, 估计长度分别为1.45 Mb和17.75 Mb。第12染色体的代换区间分别为RM1261-RM1986和RM6869-RM5990, 估计长度分别为5.4 Mb和3.1 Mb。其中, 最短代换长度为0.45 Mb, 最长代换长度为17.67 Mb。代换总长度合计109.62 Mb, 平均代换长度为6.85 Mb (图1)。
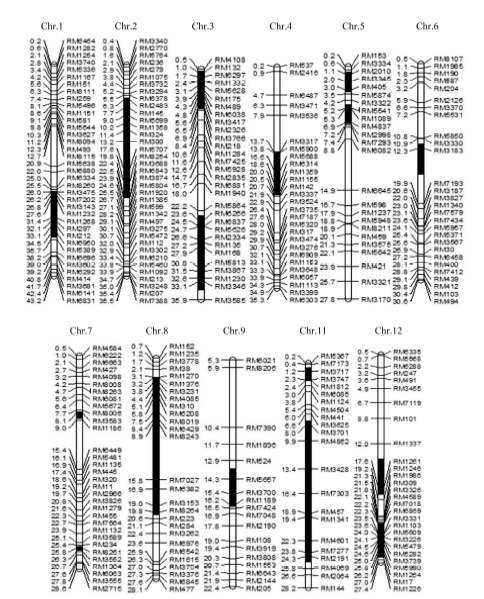 | 图1 Z519的染色体代换片段(图中黑色区段为代换片段所在的位置, 染色体左端数字为物理图距(Mb), 右侧为分子标记。)Fig. 1 Chromosome substitution segments of Z519(Black intervals indicate substitution segments. The digits on the left of chromosomes represent physical distance (Mb) while the markers are on the right.) |
Z519的芽鞘长到近3 mm时, 其尖端开始出现紫红色线条, 随着芽鞘的不断伸长和叶片的长出, 叶鞘颜色逐渐加深。随着生育进程的推进, 紫色条纹也在柱头、茎维管束、叶边缘等部位呈现, 而受体日本晴的各部位均呈绿色(图2-A~E)。与日本晴相比, Z519的株高显著降低(5.9 cm), 主穗总粒数、实粒数和千粒重显著增加, 主穗长、有效穗数和结实率无显著差异(表1)。暗示Z519的代换片段上可能含有矮化、多粒和粒重等有利QTL, 因而是一个有重要应用价值的材料。
 | 图2 日本晴和Z519的表型(A: 苗期日本晴和Z519的表型; B: 分蘖期日本晴、Z519和供体R225的鞘; C: 出芽期日本晴和Z519的鞘; D: 日本晴和Z519的雌蕊柱头; E: 抽穗期日本晴和Z519的表型。)Fig. 2 Phenotype of Nipponbare and Z519(A: plant morphology of Nipponbare and Z519 at the seeding stage; B: sheath of Nipponbare, Z519 and donor R225 at the tillering stage; C: sheath of Nipponbare and Z519 at the sprouting stage; D: pistil stigma of Nipponbare and Z519; E: plant morphology of Nipponbare and Z519 at the heading stage.) |
| 表1 日本晴和Z519的农艺性状 Table 1 Agronomic traits of Nipponbare and Z519 |
在抽穗期, Z519剑叶中的叶绿素a、叶绿素b和总叶绿素含量极显著高于日本晴(图3-A); Z519叶鞘中的叶绿素a、叶绿素b和总叶绿素含量与日本晴无显著差异(图3-B); 花青素含量在Z519叶鞘中极显著高于日本晴, 在剑叶中与日本晴无显著差异(图3-C)。
 | 图3 日本晴和Z519的叶绿素及花青素含量(A: 抽穗期剑叶的叶绿素含量; B: 抽穗期鞘的叶绿素含量; C: 抽穗期剑叶和鞘中的花青素含量。)Fig. 3 Contents of chlorophyll and anthocyanidin of Nipponbare and Z519(A: chlorophyll content of the flag leaves at the heading stage; B: chlorophyll content of the sheath at the heading stage; C: anthocyanin content in the sheath and flag leaves of Nipponbare and Z519 at the heading stage.) |
日本晴与紫鞘代换系Z519杂交产生的F1植株均表现为紫鞘, F2代植株呈现紫鞘与绿鞘分离, 经卡平方测验, 紫鞘(724)∶ 绿鞘(276)符合3∶ 1的分离比(χ 2= 1.67≤ χ 2(1, 0.05) = 3.84), 表明Z519紫鞘表型受显性单基因调控。
以16个代换片段中效果较好的标记对其中的30个隐性绿鞘单株进行基因连锁分析, 发现位于第1染色体的SSR标记RM3475可能与PSH1连锁。然后用10株紫鞘单株和10株绿鞘单株进一步验证表明, 该标记确实与PSH1连锁。最后以该代换片段上的所有标记对F2群体中的276株隐性绿鞘单株将其初定位于第1染色体SSR标记RM3475和RM7202之间, 其遗传距离分别为4.53 cM和4.89 cM。在该区间内设计了3个SSR标记和2个InDel标记, 其中4个具有多态性(表2)。以4个多态性的标记对276株隐性绿鞘单株进行精细定位, 最终将其定位于InDel标记L03和SSR标记L01之间, 物理距离为37.8 kb (图4)。
| 表2 定位区间的多态引物 Table 2 Polymorphism primers for gene mapping |
经相关软件基因预测, 在定位区间内共有6个基因。其中有2个表达蛋白、1个逆转座子、1个假定蛋白、1个编码OsFBX20-F-box结构域蛋白和1个编码质体ATP/ADP转运蛋白基因(表3)。
| 表3 定位区间内的基因预测 Table 3 Gene prediction in the mapped region |
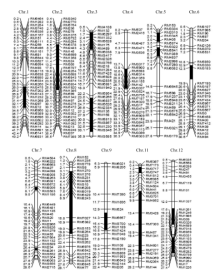
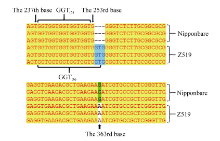
通过分析这些基因, 对其中2个可能的基因(F-box结构域蛋白和质体ATP/ADP转运蛋白)进行了DNA测序, 并分别用BioEdit和Vector NTI软件进行Blast分析。结果表明, 编码F-box结构域蛋白的基因(LOC_Os01g4590)在Z519和日本晴之间无差异; 而编码质体ATP/ADP转运蛋白的LOC_ Os01g45910存在两处差异。第1处差异是, 在日本晴中LOC_Os01g45910的基因编码区从起始密码子ATG下游的238 bp到252 bp处是5个重复GGT, 共编码5个甘氨酸残基, 而在Z519中, 该编码区域多出了一组GGT, 因此编码的蛋白产物中多出了一个甘氨酸残基。第2处差异是第一外显子的第363碱基由G变成A (图5), 使密码子AAG变成AAA, 但这两个密码子均编码赖氨酸, 故并未引起氨基酸的改变。因此, LOC_Os01g45910被初步确定为紫鞘的PSH1的候选基因, 该基因全长5270 bp, 有5个外显子和4个内含子(图4)。通过qRT-PCR分析, LOC_ Os01g45910基因在Z519的表达量比在日本晴中明显降低(图6), 进一步辅证了LOC_Os01g45910是PSH1候选基因。测序时所用的DNA引物及qRT-PCR所用引物见表4。
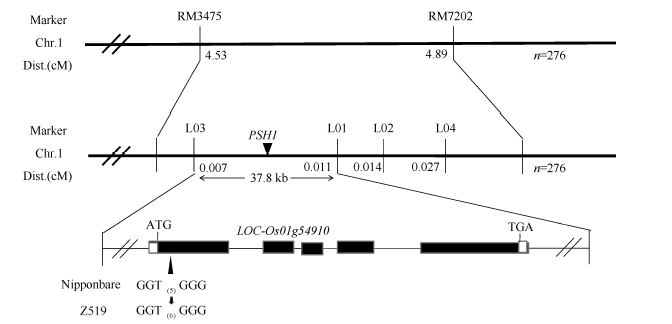 | 图4 PSH1基因分子定位(Chr.1: 第1染色体; Dist.: 遗传距离; GGT(5)为5个GGT重复; GGT(6)为6个GGT重复。)Fig. 4 Molecular mapping of PSH1(Chr.1: chromosome 1; Dist.: genetic distance; GGT(5): 5 GGT repeat; GGT(6): 6 GGT repeat.) |
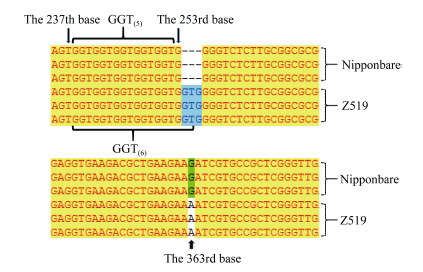 | 图5 候选基因LOC_Os01g45910的比对分析(5 GGT: 5个GGT重复; 6 GGT: 6个GGT重复。)Fig. 5 Blast analysis of the candidate gene LOC_Os01g45910(5 GGT: 5 GGT repeat; 6 GGT: 6 GGT repeat.) |
 | 图6 LOC_Os01g45910在日本晴和Z519中的表达量Fig. 6 Expression level of LOC_Os01g45910in Nipponbare and Z519 |
| 表4 PSH1候选基因的DNA的测序引物序列及qRT-PCR的引物序列 Table 4 Primers of DNA sequencing and qRT-PCR |
水稻染色体片段代换系与受体亲本只存在代换片段的差异, 可将复杂性状分解为单个孟德尔因子, 是用于QTL定位的良好材料, 尤其作为一个强大的工具, 这些代换系可很好地将基于优良品种遗传背景下鉴定出的高产、优质、抗性基因与水稻育种计划紧密结合[25, 26]。本研究采用已全基因组测序的粳稻日本晴为受体亲本、西南大学选育的优良紫鞘恢复系R225为供体亲本, 通过高代回交, 并结合分子标记辅助选择及表型双重选择的方法, 鉴定了一个紫鞘染色体片段代换系Z519。Z519包含16个代换片段, 分布于除第10染色体外的其他11条染色体, 平均代换长度为6.85 Mb。该代换系的株高显著低于日本晴, 主穗总粒数、实粒数和千粒重显著高于受体日本晴, 有效穗数、主穗粒长和结实率与受体无显著差异, 因而Z519在基因聚合育种、有利基因的进一步分离和功能研究方面具有重要的应用价值。
花青素属于类黄酮类物质, 在植物生长发育、防御敌害等方面意义重大, 具有清除自由基、抗癌和抗氧化等多种生物活性; 对肿瘤和心血管等疾病的治疗与预防有着重要的作用[27]。目前, 关于水稻紫色性状的遗传已有相关研究, 但由于所用紫色稻材料的不同, 在不同遗传背景下水稻紫色基因及紫色抑制基因数目的不同, 导致研究结果也不尽一致, 但基本上紫色为显性[16, 28, 29, 30, 31, 32, 33]。本研究鉴定的水稻染色体片段代换系Z519的叶鞘颜色随生育进程逐渐加深, 紫色条纹也在柱头、茎维管束、叶边缘等部位呈现。经遗传分析, 该紫鞘表型受单基因显性调控。进一步以日本晴和Z519回交并自交产生的F2隐性群体为材料, 将PSH1定位于第1染色体长臂分子标记L03和L01之间37.8 kb的区域。PSH1不同于Hu等[34]从紫叶Purple522的cDNA文库中分离出的2个有bHLH结构的Ra和Rb基因(位于第1染色体的R2347-C808之间)。同时, 也完全不同于定位于第6染色体的PSH(t)[16]和OsC1[2]及第11染色体的OsA1[2]。PSH1与Wang等[16]定位于第1染色体23.5 kb区域的PSH1(t)位置相近, 但其没有确定候选基因。本研究经基因预测和测序, 发现Z519在一个编码质体ATP/ADP转运蛋白的LOC_Os01g45910基因第一外显子的第238~252碱基的5个G、T、G重复区又多插入了3个碱基G、T、G, 导致增加了一个甘氨酸, 而且其表达量在Z519中明显降低。我们初步认为该基因可能是PSH1的候选基因。该蛋白主要存在于细胞膜上, 影响物质的跨膜运输。已有研究表明, 与花青素转运有关的MATE家族, 通过H+/Na+的逆向转运, 在ATP的作用下把花青素转入液泡[35]。因而PSH1可能是影响花青素转运的一个新基因, 至于PSH1编码蛋白跨膜转运的机制尚不清楚, 还需进一步深入研究。因而克隆该基因对花青素的分子遗传机制研究具有重要意义。
鉴定了一个以日本晴为受体、优良恢复系R225为供体的水稻染色体片段代换系Z519。Z519包含16个代换片段, 分布于除第10染色体外的其他11条染色体, 平均代换长度为6.85 Mb。Z519表现紫鞘, 并在叶缘、茎秆维管束和柱头等部位呈现紫色条纹, 其叶鞘的花青素含量和剑叶中的叶绿素含量显著高于日本晴。该代换系的株高显著降低, 千粒重、主穗总粒数和实粒数显著增加, 有效穗数、主穗长和结实率与受体日本晴无显著差异。该紫鞘性状受1对显性核基因控制, 被定位于水稻第1染色体L03和L01之间37.8 kb的区域, 被命名为PSH1。初步确定编码质体ATP/ADP转运蛋白的基因LOC_Os01g45910为PSH1候选基因, 目前尚未被克隆, 可能是一个调控花青素转运的新基因。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|