




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
簇毛麦6VS特异转录序列 P21461及 P33259的获得及其分子标记在鉴定小麦-簇毛麦抗白粉病育种材料中的应用
[刘畅, 李仕金, 王轲, 叶兴国, 林志珊*  ]
]
]
|
|


第一作者联系方式: E-mail: 15120067976@163.com
簇毛麦6V#2S和6V#4S染色体臂分别携带抗白粉病基因 Pm21和 PmV, 在与小麦的杂种后代中, 抗病基因与外源染色体臂共分离。开发鉴定2条外源染色体臂间多态性的序列, 尤其是遗传信息相对缺乏的6V#4S染色体臂的序列, 对于其在遗传与育种上的应用具有重要意义。本研究以携带6V#4S·6DL染色体的小麦易位系Pm97033及感病小麦亲本宛7107接种白粉菌的叶片转录组数据为资源, 通过差异基因筛选、共线性分析、簇毛麦基因组扩增及测序验证的方法, 鉴定出来自6V#4S的表达序列 P21461和 P33259, 其中基于 P21461序列设计的引物P461-5在簇毛麦6V#2S和6V#4S染色体臂的扩增产物具有30 bp的InDel和4 nt的多态性。用该引物转化的标记P461-5a可以鉴定抗白粉病小麦品种和高代品系所含的外源染色体, 显示其在簇毛麦抗源鉴别和小麦抗病育种辅助选择中潜在的应用价值。根据 P33259开发的标记P259-1可以对含有6V#4S染色体臂的材料进行特异扩增, 但对6V#2S·6AL易位染色体没有扩增产物, 因此P259-1可作为6V#4S·6DL易位染色体的特异分子标记。qRT-PCR分析结果显示, P21461的表达不受白粉菌诱导, 而 P33259在接菌后12 h和24 h的转录水平比接菌前提高约2倍, 推测其可能参与Pm97033与白粉菌的早期互作。
Dasypyrum villosum carries powdery mildew (PM) resistance gene Pm21 and PmV on its chromosome arms of 6V#2S and 6V#4S, respectively. The two resistant genes are co-segregated with exogenous chromosome arms in the offspring of hybrids between translocation lines and common wheat varieties. Developing polymorphic expression sequences to identify the two exogenous chromosomal arms is of great significance for their application in genetics and breeding, especially for 6V#4S chromosome, on which hereditary informations relatively lack. In this study, transcriptome of leaves of 6V#4S·6DL translocation line Pm97033 and common wheat line Wan7107 inoculated with Bgt was used as resources for sequences screening. As results, two unigenes of P21461 and P33259 from 6V#4S were identified by differential gene screening, synteny analysis, genomic DNA amplification and products sequencing of D. villosum accordingly. By using a pair of designed primer P461-5 based on P21461 sequence, it was found that there were 30 bp of InDel and 4 nt polymorphism between chromosomal arms of 6V#2S and 6V#4S. By the newly developed marker P461-5a, which retains the InDel polymorphism and truncates the product size amplified by P461-5, several wheat varieties and advanced lines with strong resistance to powdery mildew were identified to contain Pm21 or PmV. This marker showed potential application in discriminating D. villosum resources and assisted selection breeding for PM resistance in wheat. A marker P259-1 developed according to sequence of P33259 could specifically amplify a fragment from the wheat lines containing 6V#4S chromosome arm, but not amplify any fragment in the wheat lines containing 6V#2S·6AL translocation chromosome. Thereby, P259-1 can be used as a 6V#4S·6DL specific molecular marker in wheat backgrounds. The qRT-PCR assay showed that the expression of P21461 was not induced by PM. While transcriptional levels of P33259 increased about two times at 12 h and 24 h after inoculating the pathogens, suggesting that it might be involved in early interaction between Pm97033 and the pathogen.
簇毛麦[Dasypyrum villosum(L.) P. Candargy (syn. Haynaldia villosaSchur)]是一个起源于地中海东北部的小麦族簇毛麦属二倍体种(2n = 2x = 14, VV) [1, 2], 长期生长在恶劣、干旱的环境中, 含有许多生物和非生物胁迫抗性基因及优质基因[3], 是改良小麦的优良基因源。已有学者先后将簇毛麦基因组转移到小麦的遗传背景, 分别育成3套染色体附加系[4, 5, 6]。根据簇毛麦来源的不同, Qi等[7]将Sears培育的小麦-簇毛麦附加系用DA1V#1~DA7V#1表示, 将南京农业大学培育的小麦-簇毛麦附加系用DA1V#2~DA7V#2表示; Liu等[5]将Lukaszewski等培育的附加系用DA1V#3~DA7V#3表示; 本课题组把来自前苏联的簇毛麦No.1026的6V染色体命名为6V#4 [8]。来源不同的6V染色体短臂(6V#1S、6V#2S、6V#3S和6V#4S)对小麦白粉病的抗性表现不同, 携带6V#1S和6V#3S的小麦不抗白粉病, 而携带6V#2S和6V#4S的小麦对白粉病菌表现免疫[5, 7]。
由Blumeria graminisf. sp.tritici引起的小麦白粉病是一种严重威胁小麦生产, 且在世界范围内普遍发生的真菌性病害。抗病品种对控制小麦白粉病和减少产量损失发挥了极其重要的作用, 因而, 选育高抗新品种和改良现有推广品种的白粉病抗性, 是小麦育种的一项重点任务, 也是保障小麦安全生产最经济、有效的措施之一。虽然迄今已在小麦农家品种和野生近缘种中发掘了多个白粉病抗性基因, 并开发了一些抗病基因的分子标记, 但白粉菌菌系变异快, 很多抗病基因在生产上应用不久就被新致病菌系所克服。簇毛麦-普通小麦6V#2S· 6AL和6V#4S· 6DL易位系对白粉病菌所有菌株免疫, 表现出广谱抗性, 尤其是含6V#2S的易位系已被广泛用于小麦育种[9], 而对6V#4S易位系抗病基因鉴定和应用的研究还比较少。目前, 国内培育的一些小麦品种或高代品系, 系谱显示包含这2个不同的易位系, 但其后代的抗源归属还有待鉴定。
随着组学技术的发展, 转录组测序技术正在被广泛应用[10, 11, 12, 13]。通过大规模的基因表达分析, 鉴定植物对生物与非生物胁迫的响应基因及其代谢通路, 使人们可以在新的角度上深入了解植物适应环境的分子机制。Zhang等[14, 15]对接种条锈菌及白粉菌的小麦N9134的大规模转录组测序比较分析发现, 在响应条锈病菌及白粉菌侵染时小麦不同基因被激活, 并对新的及响应真菌的lincRNA进行了全基因组鉴定和功能预测。Hao等[16]对小麦品种Xingzi 9104的转录组测序分析, 为更深入地了解小麦条锈病成株期的抗病机制奠定了基础。Li等[17]将小麦品种YN15和小麦-中间偃麦草二体异附加系SN306接种白粉病菌, 分别对其进行转录组测序, 鉴定了一些在SN6306中表达而在YN15中不表达的unigene。可见, 植物-病原菌互作的转录组提供了互作双方在特定时空下相互应答的表达基因, 是抗病候选基因的鉴定和分子标记开发的基础。
Pm97033是利用硬粒小麦Mexicali75和簇毛麦No.1026合成的双二倍体TH3 (AABBVV)为亲本与小麦宛7107杂交、连续3次回交, 结合未成熟胚和花药培养而选出的抗白粉病的稳定品系[18], 其体细胞染色体数2n = 42, 花粉母细胞减数分裂中期I显示21II。原位杂交和重双端体分析表明, Pm97033中簇毛麦6V#4S染色体臂易位到小麦6DL上[19]。由于簇毛麦与小麦亲缘关系较远, 其杂种6V#4S染色体与小麦6DS染色体难以配对和交换, 整个易位的外源染色体臂形成一个完整的遗传单位传递。因此, 在常规杂交育种中, 6V#4S上的特异序列都与白粉病抗性共分离, 用常规方法对6V#4S的抗病基因的鉴定与分离十分困难。
本研究利用Pm97033与白粉菌互作早期的转录组数据, 筛选、鉴定来自6V#4S染色体臂上转录的基因序列, 分析它们在簇毛麦6V#2S和6V#4S染色体臂间的序列多态性, 进而开发分子标记, 并鉴定携带不同簇毛麦白粉病抗性基因的小麦品种及高代品系。同时, 初步分析外源特异基因在寄主与病原菌互作过程中的表达模式, 为进一步对抗病相关基因的鉴定和筛选提供信息。
包括携带6V#4S染色体臂的小麦品系Pm97033及其簇毛麦亲本No.1026, 已知抗病基因的小麦品种, 抗白粉病高代品系(表1)。其中, Pm97033及其簇毛麦亲本No.1026, 小麦感白粉病亲本宛7107, 以及6V#2S· 6AL易位系10SR3124由本课题组保存; 16网簇毛麦-1由南京农业大学陈佩度教授惠赠; 扬麦18、扬麦22由江苏省里下河地区农业科学研究所张伯桥研究员提供; 金禾9123由河北农林科学院遗传生理研究所刘永伟助理研究员提供; 抗白粉病高代品系由中国农业科学院作物科学研究所小麦分子育种课题组培育。
| 表1 试验材料的名称、系谱/来源及对白粉病菌的反应 Table 1 Name, pedigree/origin, and reaction to powdery mildew of plant materials used in the study |
白粉菌为北京流行的小麦白粉菌混合菌株, 由本实验室在温室感病小麦植株上收集, 在花盆密植的感病品种上大量繁殖, 以保证接种时白粉菌孢子充足、新鲜。
取宛7107和Pm97033各60粒种子, 于培养皿中浸种萌发, 选发芽均匀一致的种子种于长方形塑料盆, 每品种2行, 每行各25粒。罩以保鲜膜, 防止空气中的白粉菌孢子落入, 并在人工气候培养箱中(光照16 h/黑暗8 h, 温度24℃/20℃)培养。在小麦幼苗长至二叶一心时, 将第2片叶固定于平板上, 背面朝上, 在叶片中间叶段采取抖落法高密度均匀接种白粉菌分生孢子, 其后用保鲜膜罩保湿, 放在人工气候培养箱中(光照16 h/黑暗8 h, 温度22℃/20℃)培养。分别于接菌后20 h和48 h剪取接菌的叶段, 保存于液氮中用于转录组测序。
委托北京诺禾致源生物信息科技有限公司基于Illumina技术测序平台, 构建常规转录组文库, 利用双末端测序方法进行转录组测序。
转录组测序后, 去除小麦和白粉菌参考基因组序列后的数据进行无参拼接, 从差异表达基因中, 筛选宛7107无表达的基因序列, 在https://urgi. versailles.inra.fr/blast/blast.php进行比对, 选择与小麦第6同源群染色体短臂的基因序列具有最高匹配值的序列。基于该候选目的序列设计引物(表2), 用其对Pm97033的簇毛麦亲本No.1026进行扩增, 将PCR产物回收、测序。
扩增反应体系25 µ L, 含1 μ L基因组DNA (80 ng μ L-1), 2.5 μ L 2× Taq MasterMix, 2 µ L引物(表2, 10 mol L-1), 19.5 μ L ddH2O。扩增反应条件为95℃ 5 min; 94℃ 30 s, 60℃ 30 s, 72℃ 1 min, 39个循环; 72℃ 5 min。引物P461-5a的PCR扩增的反应条件为95℃ 5 min; 94℃ 30 s, 61℃ 30 s, 72℃ 1 min, 38 个循环; 72℃ 5 min。PCR产物经3%琼脂糖凝胶, 120 V条件下电泳40 min。
将目的片段回收后进行pZeroBack载体连接, 总反应体系为20 µ L, 其中连接体系包括10× T4 Ligase buffer 2 µ L, 0.5 µ L PLB Vector (35 ng μ L-1), 0.5 µ L T4 DNA Ligase (5 U μ L-1, Thermo Scientific), 2 µ L目的片段(30 ng μ L-1), 4℃过夜连接。将连接产物转化TOP10大肠杆菌, 涂于氨苄霉素抗性的LB平板上, 至37℃培养箱过夜培养16 h, 对转化得到的目的片段大肠杆菌单克隆进行初筛检测、测序验证。
在Pm97033接菌后0、12、20、24、48和72 h取样, 利用TRIzol试剂(Life Technologies, USA)提取RNA, 利用HiFiScript gDNA Removal cDNA Synthesis Kit (北京康为世纪生物科技有限公司)合成第一链cDNA。以P21461为目的基因, 其特异引物为21461-F和21461-R; 内参基因为Ta2291[27], 其引物为ARF-F和ARF-R (表2)。反应条件为95℃ 15 s, 60℃ 20 s, 72℃ 30 s, 40个循环, 然后95℃ 15 min, 60℃ 1 min, 95℃ 30 s, 60℃ 15 s。反应在ABI 7500荧光定量PCR仪上进行。设置每个反应3个重复, 采用2-Δ Δ Ct法进行基因相对定量分析[28]。参照Li等[17]描述的方法, 如果某基因相对表达量达Pm97033在0 h (对照)的表达水平2倍或2倍以上, 则认为该基因受白粉菌诱导上调表达; 相反, 如果基因相对表达量低于对照的表达水平, 则认为该基因受诱导下调表达; 当基因相对表达量介于上调与下调表达之间时, 认为该基因表达量无变化。
| 表2 本研究设计的引物 Table 2 Primers used in this study |
序列P21461和P33259是转录组数据中2条在Pm97033和宛7107间差异表达的序列。在抗病品种Pm97033接种白粉菌20 h后, 序列P21461和P33259的Read count值分别为288和320, 在接菌48 h后的Read count值分别为353和10; 而感病品种宛7107在相同时间点, 样品转录测序Read count值都为0。将这些序列在https://urgi.versailles.inra. fr/blast/blast.php中进行Blast, 其最高分值的匹配片段均位于小麦6DS, 说明这些序列很可能位于小麦族第6同源群染色体短臂上。
P21461测序获得的序列长度为3135 bp。针对P21461序列的不同区域设计引物, 分别在Pm97033及其抗、感亲本(簇毛麦No.1026和宛7107)中扩增, 结果有3对引物在No.1026和Pm97033中有一致带谱, 其中P461-5在Pm97033和No.1026中有一条扩增带, 而在宛7107中无扩增产物(图1-A)。分别对P461-4和P461-5在Pm97033和No.1026中的扩增产物回收和测序, 测序结果与原转录组测序信息完全一致。
同理, 对P33259序列设计了2对引物(P259-1和P259-3), 扩增结果显示, 两对引物都只能在Pm97033和No.1026中扩增出大小一致的片段(图1-B), 并获得测序验证。因此认为, P21461和P33259均位于Pm97033携带的6V#4S染色体臂上。
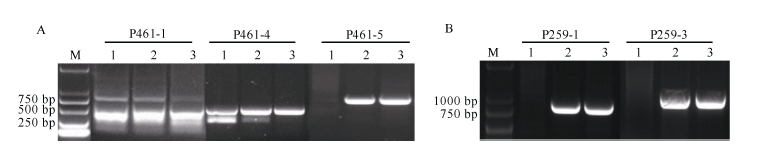 | 图1 不同引物在Pm97033及其簇毛麦和小麦亲本中的扩增谱带Fig. 1 Banding patterns of different primers amplified in Pm97033 and its parents of D. villosumand common wheat |
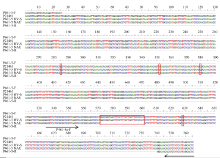
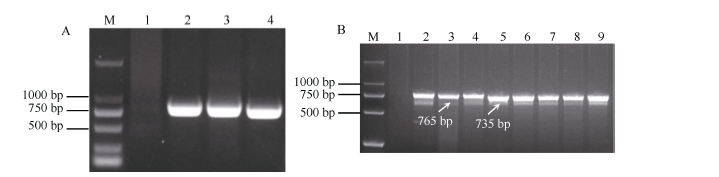
2.2.1 引物P461-5扩增产物在不同易位系中的多态性 利用P21461的特异引物P461-5检测6V#2S· 6AL易位系10SR3124, 可见其扩增产物与在6V#4S· 6DL易位系Pm97033和簇毛麦No.1026中的扩增带型非常接近, 而宛7107没有该扩增条带(图2-A)。回收P461-5对10SR3124的扩增片段, 连接到pZeroBack载体上测序验证, 结果该扩增片段与原始序列存在30 bp的InDel和4个碱基的替换(图3)。
 | 图2 引物P461-5在携带6V#2S和6V#4S的小麦易位系及已知系谱的抗白粉病小麦品种中的扩增谱带Fig. 2 Amplification profile of primer P461-5 in the 6V#2S and 6V#4S translocation lines and wheat varieties with known pedigree and powdery mildew resistance |
进一步用引物P461-5对一些已知系谱的小麦白粉病抗性品种或品系进行扩增, 获得3种带型。结合抗源系谱信息(表1), 可知在宛7107不含6V#4S或6V#2S, 无P461-5扩增产物; 簇毛麦No.1026、Pm97033和扬麦22携带6V#4S, 其扩增产物为765 bp; 10SR3124、内麦836、扬麦18、CB037和金禾9123携带6V#2S, 其扩增产物为735 bp (图2-B)。这一结果表明, P461-5在小麦不同遗传背景中都可有效检测来自不同簇毛麦6VS染色体。
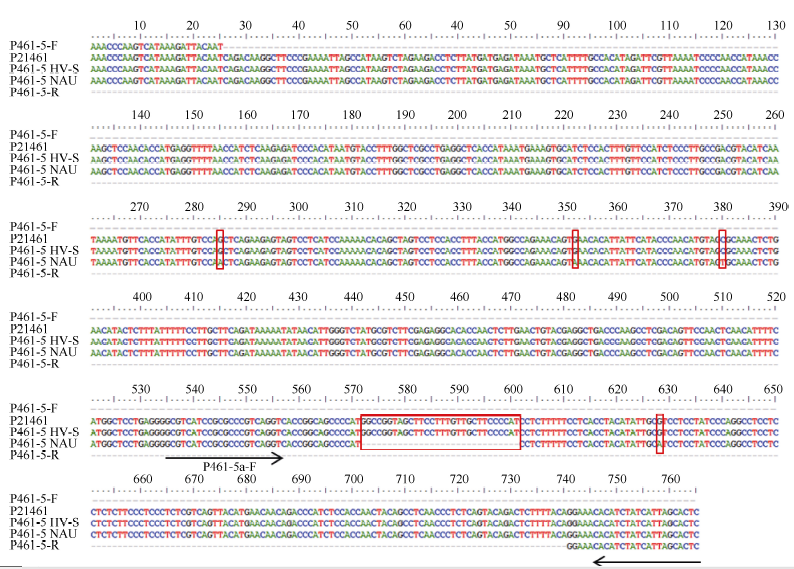 | 图3 引物P461-5在簇毛麦No.1026及6V#2S· 6AL易位系10SR3124中的扩增片段与原转录组序列的核苷酸序列比对图Fig. 3 Alignment of nucleotide sequences between the original transcriptome and the amplified fragments in D. villosum No.1026 and the 6V#2S· 6AL translocation line 10SR3124 using primer P461-5 |
2.2.2 引物P259-1扩增产物在不同易位系中的多态性 利用P33259特异引物P259-1对不同来源簇毛麦6V染色体易位系扩增表明, 在簇毛麦No.1026及其衍生易位系Pm97033中可扩增出829 bp的目标带, 而在16网簇毛麦-1及其衍生的易位系10SR3124中没有任何扩增带(图4-A)。用一些已知系谱的小麦白粉病抗性品种检验该引物的有效性, 发现829 bp的目标带只出现在携带6V#4S的3个材料(簇毛麦No.1026、Pm97033和扬麦22)中, 而在宛7107及携带6V#2S的10SR3124、内麦836、扬麦18、CB037和金禾9123中未检测到扩增产物(图4-B)。该结果与系谱(表1)分析相吻合, 说明在小麦不同的遗传背景中, P259-1可稳定扩增来自簇毛麦6V#4S染色体的特异带。
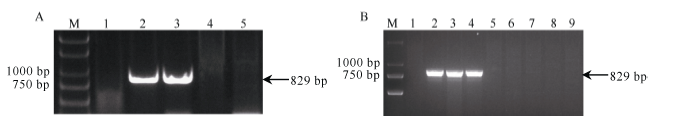 | 图4 引物P259-1在携带6V#2S和6V#4S的小麦易位系及已知系谱的抗白粉病小麦品种中的扩增谱带Fig. 4 Amplification profile of primer P259-1 in the 6V#2S and 6V#4S translocation lines and wheat varieties with known pedigree and powdery mildew resistance |
引物P461-5对6V#2S· 6AL和6V#4S· 6DL的扩增分别获得大小为765 bp 和735 bp的片段, 在这一长度范围内, 30 bp的差异在1%~3%琼脂糖凝胶中较难区分(图2-A)。为了便于在育种上的利用, 我们在30 bp的InDel区域附近, 重新设计一对引物P461-5a (图3), 缩短了扩增片段长度, 使其对6V#4S和6V#2S的特异扩增差异更明显(图5-A)。利用该引物对白粉病免疫的高代品系进行鉴定, 只有16Ma385扩增出与Pm97033相同的231 bp大小的片段, 表明其含有6V#4S· 6DL染色体, 其余4个品系均扩增出201 bp的片段, 表明含有6V#2S· 6AL染色体(图5-A)。值得一提的是, 系谱显示在CB033选育过程中, 先后利用了含有6V#4S· 6DL和6V#2S· 6AL染色体易位系作为亲本, 但CB033中仅保留了6V#2S· 6AL易位染色体。
利用引物P259-1检测上述抗白粉病小麦高代品系显示, 以Pm97033为亲本育成的16Ma385可扩增出829 bp的目的片段, 而在宛7107及以6V#2S· 6AL易位系为亲本培育的CB034、16Ma401中均无扩增产物。与P461-5a的鉴定结果一致, 在CB033及其衍生系16Ma409中, 也只检测到6V#2S特异带, 没有检测到6V#4S特异带(图5-B)。
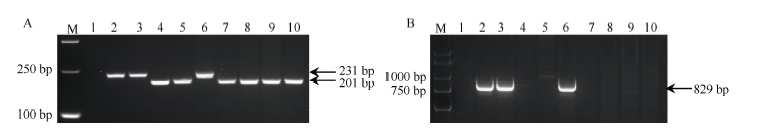 | 图5 引物P461-5a (A)和P259-1 (B)在不同小麦易位系、簇毛麦亲本及抗白粉病高代品系中的扩增Fig. 5 Amplification and identification of different wheat translocation lines, D. villosum parents, and wheat advanced lines with PM-resistance using primers P461-5a (A) and P259-1 (B) |
为了解P21461和P33259是否受白粉菌的诱导, 对接菌后不同时间点的小麦抗病易位系Pm97033的样品进行qRT-PCR分析, 结果表明, 与接菌前(0 h)的转录水平相比, P21461在接菌后12、20、24和48 h的转录水平没有明显变化, 直到接菌后72 h转录水平略有提高(图7-A), 推测该基因与Pm97033对白粉菌的早期抗性没有直接关系。而P33259的转录则呈现出受白粉菌诱导的趋势, 与接菌前(接菌0 h)的转录水平相比, 接菌后12 h转录水平提高(2.5倍), 20 h时转录水平虽比12 h的转录水平有所降低, 但到24 h时转录水平再次提高(2.3倍), 接菌后48 h和72 h转录水平再次降低(0.6倍和0.5倍) (图7-B), 推测该基因可能与Pm97033和白粉菌的早期互作相关。
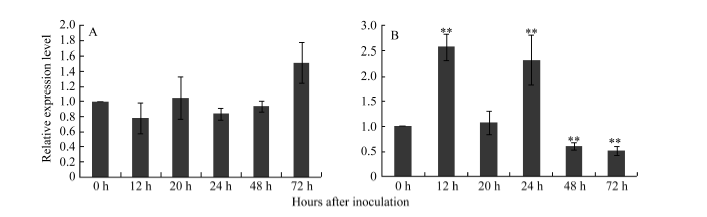 | 图6 Pm97033接种白粉病菌后72 h内P21461 (A)和P33259基因(B)的表达模式Fig. 6 Expression patterns of P21461 (A) and P33259 (B) in Pm97033 after inoculating Blumeria graminis f. sp. tritici (Bgt) for 72 hours |
如何从巨大的转录组数据中鉴定6VS的基因序列是一个很大的挑战。由于宛7107不携带6V#4S, 因此大多数目标序列必然在Pm97033中表达, 而宛7107中无表达或低表达(来自内源基因)。同时, 根据禾本科物种间基因序列的保守性和共线性关系, 推测目标序列与小麦第6群染色体短臂上的同源序列匹配度应该具有最高的分值。因此, 我们通过选择符合上述条件的序列, 进一步通过设计引物, 对Pm97033的簇毛麦亲本基因组特异扩增和测序验证。P21461和P33259的成功鉴定充分表明这一方法用于筛选来自6V#4S的基因是有效的, 它为进一步从转录组数据中快速鉴定目标基因奠定了基础。6V#4S特异序列的获得也为该易位染色体的遗传育种研究提供了简易实用的分子标记。Li等[17]在对小麦品种YN15 和小麦-中间偃麦草二体异附加系SN6306接种白粉菌的材料进行转录组测序分析中, 主要通过对unigene的筛选, 基因表达模式的荧光定量分析及对YN15、SN6306和中间偃麦草的基因组的扩增和测序验证, 鉴定出若干候选的中间偃麦草抗白粉病基因。本研究在采用与此类似的方法前, 先将候选序列进行染色体的定位和验证, 减少了大量小麦内源基因进入后续分析。本研究的qRT-PCR结果表明, 在寄主与白粉菌的互作早期, P21461的转录水平不受白粉菌侵染的诱导, 而P33259则在接菌早期不同时间点转录水平有所提高, 推测其可能参与了寄主与病原菌早期的互作。这一结果需要进一步的验证。
我们曾经从簇毛麦No.1026基因组中分离到定位于6V#2S 染色体臂的Pm21基因位点上的关键抗白粉病基因Stpk-V的同源基因Stpk-V3[8], 证明2个同源基因序列间在内含子和启动子区存在多态性。本研究发现, 2类不同的簇毛麦易位系在转录的P21461片段中至少含有一个30 bp的InDel以及4 nt的差异。P21461的基因注释为一个未知功能的假定蛋白, 表明P21461是一个编码蛋白的基因序列, 6V#2S与6V#4S间存在某些基因编码区的多态性。
标记P259-1可从Pm97033及其簇毛麦亲本No.1026中扩增出序列一致的6V#4S特异的片段, 但对6V#2S· 6AL易位染色体及其簇毛麦亲本都没有扩增产物。基于P33259的序列, 我们设计了另一对引物也不能对6V#2S进行特异扩增, 似乎表明P33259在6V#2S中缺失, 这也再次表明两条外源染色体臂具有一定的基因结构歧化。
6V#4S和6V#2S染色体臂分别易位到了小麦的不同染色体臂上, 在各自与小麦的杂交组合中表现出了不同的遗传行为。如携带6V#4S· 6DL易位染色体的小麦品种扬麦22与扬麦16杂交时, 杂种F2的抗、感单株之比为129∶ 80, 严重偏离一对抗病基因的分离规律(χ 23:1 = 18.95, P < 0.01)[29]。我们在Pm97033与宛7107杂种F2也观察到相似的结果(χ 23:1= 7.456, P < 0.01)。而根据李桂萍等[30]的研究报道, 在6V#2S· 6AL易位系与感病小麦5个不同组合的杂种F2中, 抗、感病植株的比例符合3∶ 1的理论比值。我们最近的观察发现, 当两类不同的易位系相互杂交时, F2抗、感病株的比例符合15∶ 1, 且根据分子标记辅助分析, 发现该F2群体中含纯合、杂合或不含6V#4S· 6DL易位染色体的单株, 其分离比符合1∶ 2∶ 1 (χ 21:2:1 = 0.551, P > 0.01), 推测杂种中由于6V#4S与6V#2S间低频率的配对减少了6V#4S· 6DL染色体的丢失, 从而提高了6V#4S在该F2群体的频率[31]。尽管如此, 本研究发现高代品系CB033和CB037的杂交组合中包含了两类不同的易位系, 但CB033和CB037都仅保留了6V#2S· 6AL染色体。以CB033 (系谱为CA9211/6V# 4S· 6DL//辽春10号/3/6V#2S· 6AL)为例, 我们认为, 首先利用CA9211/6V#4S· 6DL//辽春10号杂交分离出的抗病植株作为母本再与6V#2S· 6AL易位系杂交, 此时作为母本的抗病株中6V#4S· 6DL染色体处于杂合状态, 在与纯合体的6V#2S· 6AL易位系的杂种群体中, 含有6V#4S· 6DL染色体的植株频率低于群体的50%, 而含有6V#2S· 6AL染色体的植株则为100%。因此, 在无分子标记辅助选择的情况下, 仅依靠抗病表型获得含有6V#2S· 6AL易位染色体植株的频率远高于含有6V#4S· 6DL染色体的植株。
利用转录组测序数据成功筛选鉴定了2个来自Pm97033中簇毛麦6V#4S染色体臂的基因片段P21461和P33259, 将P21461转化成能在不同小麦遗传背景中区分6V#2S和6V#4S染色体臂的特异分子标记P461-5a, 而基于P33259的序列开发的标记P259-1可特异跟踪小麦遗传背景中的6V#4S· 6DL染色体, 并将2个标记用于鉴定育成的不同小麦-簇毛麦抗白粉病品种(系)。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|