

{kind=link}
{kind=link}
{kind=link}
{kind=link}
源于陕A群、陕B群玉米自交系在不同密度条件下配合力分析
[王博新 , 王亚辉, 陈朋飞, 刘徐冬雨, 冯志前, 郝引川, 张仁和, 张兴华, 薛吉全
, 王亚辉, 陈朋飞, 刘徐冬雨, 冯志前, 郝引川, 张仁和, 张兴华, 薛吉全* ]
, 王亚辉, 陈朋飞, 刘徐冬雨, 冯志前, 郝引川, 张仁和, 张兴华, 薛吉全]
|
|


第一作者联系方式: E-mail:wangboxin019@163.com
采用NC-II遗传设计, 以郑58、昌7-2为测验种, 与17份高密度条件下筛选的玉米自交系组配成34份杂交组合, 2014—2015年分别于陕西杨凌、长武、榆林进行3种密度(45 000、67 500和90 000株 hm-2)配合力分析试验。采用PROC VARCOMP分析不同密度条件下产量及耐密性相关性状的遗传效应, 采用频率直方分布图研究不同密度条件下产量及耐密性相关性状一般配合力(GCA)平均数的变化规律, 利用AMMI评价玉米自交系与杂交组合的稳定性。结果表明, 产量、倒伏率、茎秆强度主要受加性遗传效应控制, 空秆率主要受非加性遗传效应控制。加性遗传效应对产量及耐密性相关性状的贡献率随种植密度的增加呈上升趋势。玉米自交系产量、空秆率、倒伏率、茎秆强度的一般配合力频率均属于正态分布, 随着种植密度的增加, 产量GCA的平均值提高了0.28, 空秆率GCA平均值降低了0.21, 倒伏率GCA平均值降低了0.03, 茎秆强度GCA平均值增加了0.02。玉米杂交组合产量与玉米自交系产量GCA密切相关( r=0.877**, r=0.811**, r=0.672**)。随着种植密度的增加, 表现稳定的玉米自交系及杂交组合的数量呈上升趋势。因此, 强化逆境选择压力, 实施高密度选择策略, 是增强玉米自交系耐密性和抗倒性, 提升一般配合力, 实现产量增益的有效措施。
, WANG Ya-Hui, CHEN Peng-Fei, LIU Xu-Dong-Yu, FENG Zhi-Qian, HAO Yin-Chuan, ZHANG Ren-He, ZHANG Xing-Hua, XUE Ji-Quan
The objective of this study was to investigate the effectiveness of high-density breeding strategy through analyzing genetic components and general combining ability (GCA) effects under different planting densities. Seventeen maize inbred lines selected under high density were test-crossed with two testers (Zheng 58, Chang 7-2) in the North Carolina Design II. Thirty-four crosses were evaluated under different density conditions (45 000, 67 500, and 90 000 plants ha-1) in 2014 and 2015 at Yangling, Changwu and Yulin in Shaanxi province. The genetic variance components for grain yield and other traits were estimated using PROC VARCOMP method. Frequency distribution was used to analyze the response of GCA for grain yield and other traits under different densities. The stability of maize inbred lines and crosses was evaluated based on AMMI model. The grain yield, stalk lodging rate and rind penetration strength were mostly controlled by additive gene action, while the inheritance of sterile plant rate controlled by non-additive gene action. Also, the contribution of additive gene action to total variance increased with increasing planting density. The average of general combining ability for grain yield, sterile plant rate, stalk lodging rate and rind penetration strength increased by 0.28, -0.21, -0.03, and 0.02, respectively, from low density to high density. The correlations between GCA of the inbred lines and their crosses were significant ( r=0.877**, r=0.811**, r=0.672* *) under different density. As the density increased, the number of stable maize inbred lines and crosses increased. These results indicate that increasing plant density to improve GCA for grain yield and other traits of inbred lines is an effective strategy to enhance tolerance to high plant density and yield stability.
自20世纪30年代开始, 世界玉米产量每年平均以61 kg hm-2的速率增加[1]。其中, 美国、巴西、中国、阿根廷的玉米产量年均增益速率分别为75、80、94和132 kg hm-2 [2, 3, 4, 5]。Duvick[6, 7]指出, 美国玉米产量提高的主要原因是抵抗逆境胁迫能力的增强, 一定意义上说产量就是抗逆性, 特别是耐密性。Troyer[8]强调适应性比杂种优势更重要。Tollenaar等[9]认为产量遗传改良归功于忍耐逆境胁迫能力的提高。张世煌[10]认为非杂种优势对产量贡献大于杂种优势的贡献, 可通过增加选择压力如密植、低氮、高温、干旱来增加选择响应。薛吉全等[11]研究发现耐密性较好的玉米品种如先玉335、郑单958对高密、低氮和干旱等逆境具有良好的适应性。玉米自交系对逆境胁迫比杂交种更敏感[12]。玉米杂交种的抗逆性来源于亲本自交系[13]。基于此, 实施高密度选择策略已成为选育耐密型玉米自交系及耐密型玉米品种的重要手段。研究不同密度条件下玉米自交系产量及耐密性相关性状的遗传效应, 以及配合力效应, 对选育耐密型玉米自交系与玉米杂交种具有重要的科学意义。
本研究采用NC-II遗传设计, 以郑58、昌7-2为测验种, 与17份高密度条件下筛选的玉米自交系组配, 通过2年3点3种密度条件下配合力分析, 明确不同密度条件下控制玉米产量及耐密性相关性状的遗传效应, 评价不同密度条件下玉米自交系产量及耐密性相关性状的配合力效应, 为开展高密度筛选策略和选育耐密型玉米品种提供科学依据。
17份玉米自交系来源于陕A群、陕B群两大杂种优势群, 均在120 000株 hm-2高密度条件下选育而来。陕A群、陕B群由西北农林科技大学玉米遗传育种课题组构建, 以提高适应性为主线, 以国内外优良种质形成的综合种为基础, 简化杂优模式, 采用两边推、两向分群的育种思路, 强化逆境人工选择, 通过多地点、高密度、少施肥和少灌水的技术路线, 2009— 2013年开展5轮30点的联合改良。以单穗粒重(大于100 g)、出籽率(大于87%)和穗行果穗入选数(大于3个以上)为指标, 经陕西、海南(每年2代)连续自交选育而成(详见表1)。
以17份高密度条件下筛选的玉米自交系为父本, 郑58、昌7-2为母本, 采用NC-II遗传交配设计, 2013年冬季在西北农林科技大学海南三亚育种基地测配出34份F1杂交组合, 供2年田间试验。2014— 2015年将34份F1杂交组合种植于陕西杨凌、长武、榆林3个试验基地。设置45 000、67 500和90 000株 hm-23种密度。采用完全随机区组设计, 3次重复, 2行区, 行长5 m, 行距0.6 m。田间管理措施同当地大田生产管理。
| 表1 陕A群、陕B群17个玉米自交系 Table 1 Seventeen maize inbred lines from Shaan A group and Shaan B group in this study |
吐丝期用茎秆强度测定仪(YYD-1型)探头垂直缓慢插入地上第3节节间中部, 茎秆强度仪上显示的记录值为茎秆穿刺强度(rind penetration strength, RPS), 于每小区随机测10株, 取平均值。
成熟期调查试验小区的总株数、空秆数(不结穗或结粒数少于20的株数)、倒伏数(茎秆倾斜45° )。收获小区全部果穗称重计产, 用PM-8188测定玉米籽粒含水量, 按照标准含水量14%计算产量。
1.4.1 联合方差分析 利用SAS 9.2软件的PROC GLM程序分别在不同密度条件下对单一环境产量及耐密性相关性状进行方差分析, 再对混合环境进行联合方差分析。各性状校正值由PROC MIXED和LSMEANS of SAS计算获得。环境(年份与地点互作)、重复视为随机效应, 品种为固定效应, 其联合方差分析模型如下[14]。
Yijg=μ +Ei+Rj(i)+Gg+EGig+ε ijg
其中, Yijg为第i环境第j重复中第g个品种的观察值; μ 为均值; Ei为环境效应; Rj(i)为重复效应; Gg为基因型效应; EGig为基因型与环境互作效应; ε ijg为误差项。
1.4.2 遗传效应分析 利用SAS 9.2软件的PROC VARCOMP程序分别分析不同密度条件下各组分方差分量。利用Anita等[15]提出的line× tester模型计算出一般配合力方差(Vg)、特殊配合力方差(Vs), 利用Baker[16]提出的模型估算加性遗传效应比值(Va/Vt)。参考Nyquist等[14]提出的公式计算广义遗传力(h2)。
Vg=$\sigma _{m}^{2}$+$\sigma _{f}^{2}$; Vs=$\sigma _{mf}^{2}$; Va/Vt=$\frac{2(\sigma _{m}^{2}+\sigma _{f}^{2})}{2(\sigma _{m}^{2}+\sigma _{f}^{2})+\sigma _{mf}^{2}}$
${{h}^{2}}=\frac{\sigma _{\text{G}}^{2}}{\sigma _{\text{G}}^{2}+\sigma _{\text{GE}}^{2}/n+{{\sigma }_{\varepsilon }}/nr}$
其中, $\sigma _{m}^{2}\sigma _{f}^{2}\sigma _{mf}^{2}$分别为母本方差、父本方差、父本与母本互作方差, $\sigma _{G}^{2}\sigma _{GE}^{2}\sigma _{\varepsilon }^{2}$分别为基因型方差、基因型与环境互作方差、误差项。n为环境数, 本试验为6。r为重复数, 本试验为3。
1.4.3 配合力分析 依据多环境增广NC-II设计, 利用line× tester模型对不同密度条件下产量及耐密性相关性状的GCA、SCA效应值进行估算, 其配合力分析模型[17]如下。
Yijk=μ +Ee+gi+gj+sij+gEeg+sEes+ε ijk
其中, Yijk为第k个环境第ij杂交组合的观察值; μ 为杂交组合均值; Ee为环境效应; g为GCA效应; s为SCA效应; gEeg为GCA与环境互作效应; sEes为SCA与环境互作效应; ε ijk为第k个环境第ij杂交组合的误差项。
1.4.4 稳定性分析 利用方差分析与主成分分析方法, 将基因型× 环境互作效应分解为多个主成分相互作用的主成分分析轴。其AMMI模型[18]如下。
${{Y}_{ger}}=\mu +{{\alpha }_{g}}+{{\beta }_{e}}+\sum\limits_{i=1}^{n}{{{\rho }_{n}}{{\gamma }_{gn}}{{\delta }_{en}}+{{\theta }_{ger}}}$
其中, ${{Y}_{ger}}$表示第e个环境下第g个基因型第r次重复观测值, $\mu $表示总体平均值, ${{\alpha }_{g}}$表示基因型平均偏差, ${{\beta }_{e}}$表示环境平均偏差, ${{\rho }_{n}}$表示第n主成分分析特征值, ${{\gamma }_{gn}}$表示第n主成分基因型特征值, ${{\delta }_{en}}$表示第n主成分环境特征值, ${{\theta }_{ger}}$表示误差。
对2年3点数据进行误差方差齐性检验, 各年份、地点间误差方差齐次(P> 0.05), 说明不同年份不同地点的数据可以合并进行联合方差分析。不同密度条件下产量及耐密性相关性状联合方差分析(表2)表明, 3种密度条件下倒伏率与空秆率在环境间差异不显著, 而产量和茎秆强度在环境间差异达到显著或极显著水平。3种密度条件下产量及耐密性相关性状在基因型间存在显著或极显著差异, 表明参试组合内存在真实性遗传差异。配合力联合方差分析(表2)显示, 除45 000株 hm-2密度条件下空秆率的一般配合力外, 3种密度条件下的产量及其余耐密性相关性状的一般配合力均存在显著或极显著差异。3种密度条件下产量、倒伏率、空秆率的特殊配合力都存在显著或极显著差异, 而茎秆强度的特殊配合力差异不显著。
| 表2 不同密度条件下玉米自交系产量及耐密性相关性状联合方差分析 Table 2 Mean squares from combined analysis of variance for grain yield and other traits of maize inbred lines under different densities |
遗传方差组成分析(表3)显示, 45 000株 hm-2密度条件下产量、空秆率、倒伏率、茎秆强度的加性遗传方差占总基因方差的比值分别为55.23%、28.05%、87.12%和67.66%; 67 500株 hm-2密度条件下各性状的加性遗传方差占总基因方差的比值为61.81%、23.86%、87.87%和87.30%; 90 000株 hm-2密度条件下各性状的加性遗传方差占总基因方差的比值为65.77%、47.20%、85.33%和75.52%。这说明产量及耐密性相关性状受基因加性效应和非加性效应共同控制。产量、倒伏率、茎秆强度的加性遗传方差占总基因型方差的55.23%~87.87%, 主要受加性遗传效应控制。空秆率的加性遗传方差占总基因型方差的23.86%~47.20%, 主要受非加性遗传效应控制。同时, 随着种植密度的提高, 加性遗传效应对产量及耐密性相关性状的贡献率均呈上升趋势。在不同密度条件下产量、空秆率、倒伏率的遗传力较为稳定, 在58.39%~72.48%区间。茎秆强度的遗传力随着种植密度的增加呈下降趋势, 在70.33%~31.22%区间(表3)。
| 表3 不同密度条件下产量及耐密相关性状的一般配合力、特殊配合力方差分量及遗传力 Table 3 The variance component of general combining ability and special combining ability and heritability for grain yield and other traits under different density |
2年3点不同密度条件下各性状的一般配合力频率分布直方图(图1)显示, 玉米自交系产量、空秆率、倒伏率、茎秆强度的一般配合力频率均属于正态分布, 并且随着种植密度的增加, 产量及耐密性相关性状的一般配合力平均数发生了移动。3种密度条件下产量GCA的平均数分别为0.06、0.22和0.34, 高密度较低密度增加了0.28 (图1-A); 空秆率GCA平均数分别为-0.07、-0.53和-0.29, 高密度较低密度降低了0.21 (图1-B); 倒伏率GCA的平均数为-0.30、-0.32和-0.33, 高密度较低密度降低了0.03 (图1-C); 茎秆强度GCA平均数分别为0.41、0.30和0.43, 高密度较低密度增加了0.02 (图1-D)。
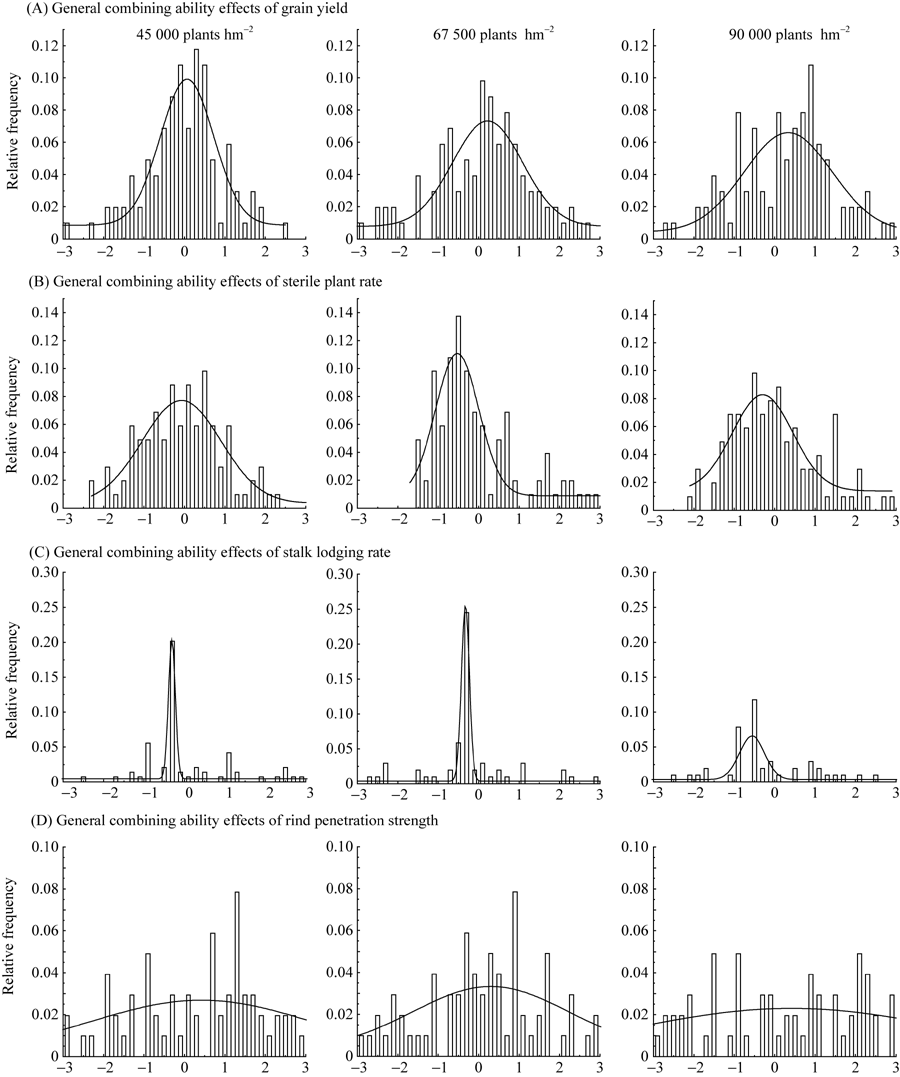 | 图1 不同密度条件下玉米自交系产量(A)、空秆率(B)、倒伏率(C)和茎秆强度(D)一般配合力的频率分布直方图Fig. 1 Frequency distribution of general combining ability effects of grain yield (A), sterile plant rate (B), stalk lodging rate (C), and rind penetration strength (D) of maize inbred lines under different densities |
不同密度条件下玉米自交系产量的一般配合力(图2), 除HN008、Z666、KA027、KA203、KA034表现为负效应外, 其余玉米自交系均表现为正效应, 其中6份玉米自交系KA103、KA105、KB081、KB024、KB215、YLB-6产量GCA随着密度的增加而显著提高。说明实施高密筛选策略, 可有效改良空秆率、茎秆强度和倒伏率的一般配合力, 进而提升产量的一般配合力。
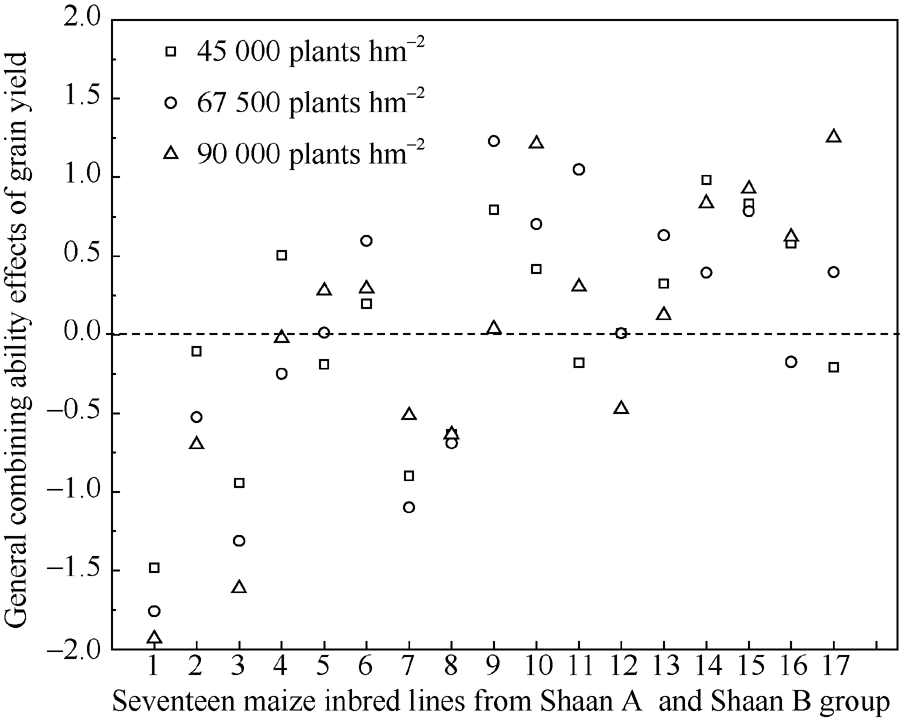 | 图2 不同密度条件下玉米自交系产量GCA表现Fig. 2 General combining ability for grain yield of maize inbred lines under different densities |
图3可知, 随着玉米自交系产量GCA的提高, 其杂交组合产量也随之提高, 而且玉米自交系产量GCA与其杂交组合的产量之间均存在极显著正相关(r=0.877* * , r=0.811* * , r=0.672* * ), 回归系数间存在显著性差异(P< 0.05), 说明玉米杂交组合的产量与玉米自交系的一般配合力密切相关。通过高密度选择, 可显著改善玉米自交系的农艺性状, 增强耐密性和抗倒性, 提升了一般配合力, 特别是高密度条件下的变化幅度优于低密度条件的变化幅度。
 | 图3 不同密度下玉米自交系的产量GCA与杂交组合产量的相关性Fig. 3 Correlation between general combining ability effects for grain yield of inbred lines and grain yield of their crosses under different densities |
图4可知, 第1主成分与第2主成分之和均大于80%, 可以有效评价玉米自交系及其杂交组合的稳定性。图中玉米自交系或杂交组合离原点位置较近, 表明玉米自交系或杂交组合稳定性较好。在3种密度条件下, 表现相对稳定的玉米自交系分别为9、8、14个, 比例为52.94%、47.06%、82.35%; 在3种密度条件下, 表现相对稳定的杂交组合分别为24、24、28个, 比例为70.58%、70.58%、82.35%。随着种植密度的增加, 稳定性良好的玉米自交系数量及杂交组合的数量呈上升趋势。高密度下筛选的玉米自交系, 在高密度条件下的稳定性较好, 组配的杂交组合表现出同样的趋势。说明, 耐密性良好的玉米自交系组配的玉米杂交组合适应性较好。
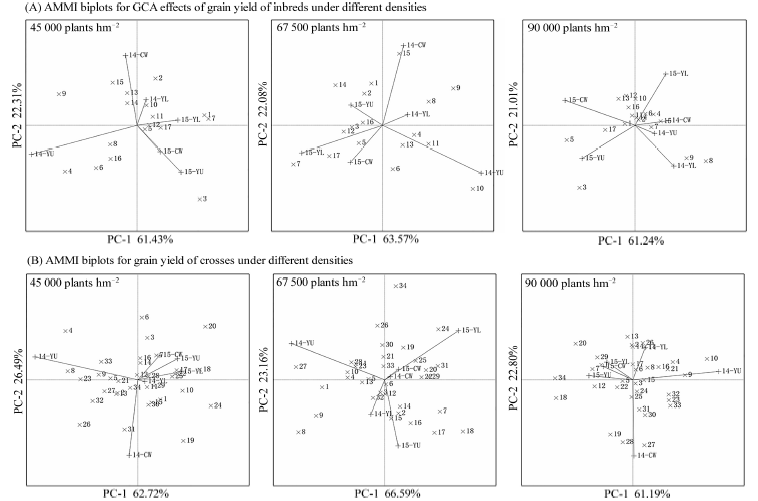 | 图4 不同密度条件下(A)玉米自交系的产量GCA, (B)杂交组合的产量AMMI交互作用双标图14-YL: 2014年杨凌, 14-CW: 2014年长武, 14-YU: 2014年榆林, 15-YL: 2015年杨凌, 15-CW: 2015年长武, 15-YU: 2015年榆林。Fig. 4 AMMI biplots for GCA effects for grain yield of inbred lines (A), grain yield of crosses (B) under different densities14-YL: at Yangling in 2014, 14-CW: at Changwu in 2014, 14-YU: at Yulin in 2014, 15-YL: at Yangling in 2015, 15-CW: at Changwu in 2015, 15-YU: at Yulin in 2015. |
玉米产量的提高关键在于群体内有利等位基因累积的多少, 而加性遗传效应反映群体内有利等位基因的累积量[19]。Edmeades等[20]曾用17个来自Tuxpeň o群体的玉米自交系进行完全双列杂交, 发现非加性遗传效应主要控制玉米产量, 但加性遗传效应控制随着种植密度的增加呈上升趋势。Badu-Apraku等[21]采用完全双列杂交对9份早熟玉米自交系分析发现, 在干旱环境下, 产量一般配合力方差大于特殊配合力方差, 表明产量主要受加性遗传效应控制。梁文科等[22]对2个热带群体及7个温带群体育种价值的评估认为, 非加性遗传效应对产量的贡献较大。本研究通过高密度条件下筛选的17份玉米自交系与郑58、昌7-2进行NC-II测配, 发现产量、倒伏率、茎秆强度主要受加性遗传效应控制, 空秆率主要受非加性遗传效应控制。随着种植密度的增加, 加性遗传效应对玉米产量及耐密性相关性状的贡献率呈上升趋势。王文斌等[23]基于SNP分析遗传多样性发现, 陕A群、陕B群选育的玉米自交系遗传背景广泛, 具有丰富的有利等位基因。通过表型鉴定与分子标记验证, 明确通过高密度选择, 有利于增加群体内遗传变异, 优化了与耐密性相关的倒伏率和茎秆强度等农艺性状, 有助于选育出优异的耐密型玉米品种。
非杂种优势对产量贡献大于杂种优势的贡献, 一般配合力对产量贡献大于特殊配合力的贡献[9, 10]。杨泽勇等[24]对不同密度条件下选育出的优良自交系鉴定, 发现高密度选系在60 000、90 000和120 000株 hm-2密度条件下产量GCA为正值的比值为33.3%、50.0%和66.6%。高瑞景等[25]对3个育种材料的S3代选系研究发现, 一般配合力最高的材料均出现在高密度选系中。梁玉娟等[26]对Reid与Laneaster类群中优良自交系研究发现, Reid类群中玉米自交系在高密度条件下具有较高的一般配合力。本研究发现, 产量、空秆率、倒伏率、茎秆强度的一般配合力均属于正态分布, 随着种植密度的增加, 产量及农艺性状的一般配合力平均数发生了移动, 产量GCA的平均值提高了0.28, 空秆率GCA平均值降低了0.21, 倒伏率GCA平均值降低了0.03, 茎秆强度GCA平均值增加了0.02。玉米自交系的产量GCA, 随着种植密度的增加而显著提高, 且玉米自交系的产量GCA与其杂交组合的产量之间存在极显著正相关(r=0.877* * , r=0.811* * , r=0.672* * ), 说明玉米杂交组合的产量与玉米自交系的一般配合力密切相关。通过高密度选择, 可显著改善玉米自交系的农艺性状, 增强耐密性和抗倒性, 提升产量及耐密性相关性状的一般配合力。
在环境优越的条件下, 植株间性状接近, 表现都较好, 差异不明显。而在逆境条件下, 个体间差异扩大, 增加了选择效率[27]。Francis等[28]研究发现高密度条件下选育的耐密型品种, 其产量明显高于老杂交品种, 而产量的变异系数明显低于老杂交组合。Matteo等[29]研究认为产量的稳定性与耐密性存在显著正相关。Dow等[30]指出高密度条件下筛选自交系与杂交组合是增强适应性的有效手段。提高筛选密度, 强化人工逆境选择是培育耐密型品种有效途径。Troyer[8]提出在选种圃中通常使用当地生产上种植密度的2倍作为选系密度, 杂交种的鉴定则采用超过平均密度的20%, 而产量试验中的最高密度要超过至少30%。本研究发现, 高密度下筛选的玉米自交系, 在高密度条件下稳定性较好, 组配的杂交组合表现出同样的趋势, 随着种植密度的增加, 稳定性良好的玉米自交系数量及杂交组合的数量呈上升趋势。研究证实高密度选择有利于优异基因的聚合, 可筛选出优异玉米自交系及优异杂交组合。
玉米杂交组合的产量与玉米自交系的一般配合力密切相关, 实施高密度选择策略对群体内基因加性效应的累积是有效的。随种植密度的增加, 加性遗传效应对产量及耐密性相关性状的贡献率呈上升趋势, 产量及耐密性相关性状的一般配合力平均数逐步朝着有利方向移动, 表现稳定的玉米自交系及测配杂交组合的数量呈上升趋势。因此, 强化逆境选择压力, 实施高密度选择策略, 是增强玉米自交系耐密性和抗倒性, 提升一般配合力, 实现产量增益的有效措施。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|