

{kind=link}
{kind=link}
{kind=link}
玉米不同组织器官谷氨酰胺合成酶同工酶表达差异及聚合方式
[王小纯1, 2, 3  , 张浩然
, 张浩然3 , 韦一昊1 , 贾喜婷3 , 谷明鑫3 , 马新明1, * ]
, 张浩然]
|
|


第一作者联系方式: E-mail:xiaochun.w@163.com, Tel: 13783586761
谷氨酰胺合成酶(GS)是作物氮同化及转移利用的关键酶, 本试验研究了玉米灌浆期不同组织器官的GS同工酶表达特性, 鉴定了玉米GS同工酶的聚合方式。Western blot结果表明, 玉米不同组织器官的GS同工酶亚基表达存在明显差异, 分子量约40 kD的GS1亚基在所有组织中均大量表达, 39 kD的GS1亚基仅在穗位节及穗柄中大量表达, 分子量约44 kD的GS2亚基在叶片等光合组织中微量表达。通过改进BNE技术, 结合胶内转移酶活性的测定, 分析了玉米GS同工酶全酶的大小; 利用2-D胶结合Western blot鉴定了GS同工酶相应的亚基组成。结果表明, 在玉米组织鉴定出3种分子量不同的GS同工酶, GS2全酶分子量约460 kD, 为十聚体; GS1全酶有2种聚合状态, 一种是分子量约410 kD的十聚体, 另一种是分子量约240 kD的五聚体形式, 可见玉米GS同工酶表达存在多种方式。
Glutamine synthetase (GS) is a key enzyme in nitrogen assimilation and recycling in cereals. In this study, the expression characteristics of GS isoenzymes in different tissues and organs of maize in grain-filling period were analyzed, and the assembly of GS isoenzymes were indentified. The GS isoforms expressed differentially in different organs were shown by Western-blot obviously; GS1 with a molecular weight of about 40 kD expressed highly in all tissues, and GS1 with a molecular weight of about 39 kD was merely expressed in the node of ear position and pedical, and GS2 with a molecular weight of about 44 kD was weakly expressed in the photosynthtic tissue such as leaf. With a modified blue naive PAGE (BNE) technique and in-gel activity analysis, the size of GS holoenzyme was calibrated; combined the 2-D gel with western-blot analysis, the subunits composition of GS isoenzymes were identified. Three GS isoenzymes with different sizes were identified in maize. GS2 holoenzyme was about 460 kD and likely a decamer, GS1 holoenzyme existed two kinds of assembly state, one was about 410 kD and likely a decamer, another was about 240 kD and more likely a pentamer; therefore, the expression of GS isoenzymes exists diversity in maize.
氮是玉米生长发育必须的大量矿质营养元素, 也是玉米产量的一个主要限制因素。在高等植物中, 谷氨酰胺合成酶(GS)/谷氨酸合酶(GOGAT)循环是氮素同化的主要途径, 是无机氮转化为有机氮的枢纽[1], GS是GS/GOGAT循环中的关键酶, 因此, GS同工酶表达成为提高作物氮素利用率的一个研究热点。
高等植物中有2种GS同工酶, 定位于细胞液的GS1和质体的GS2。研究表明GS同工酶的表达受组织器官、生长发育、新陈代谢及环境因素等的调控[2, 3, 4]。GS1主要参与蛋白质等含氮有机化合物降解产生氨的再同化及转移利用[1], GS2主要参与光呼吸和硝酸盐还原产生的氨的同化[5]。高等植物GS2亚基较大(42~45 kD), 由单一核基因编码, 而GS1亚基较小(38~40 kD), 由2~5个核基因编码[6, 7, 8, 9, 10, 11, 12]。玉米GS2亚基约44 kD, 由单一核基因编码, GS1亚基39~40 kD, 由5个核基因编码[13]。
植物GS必须组装成聚合体才具有催化活性, 早期电子显微镜研究表明大豆GS1全酶是八聚体, 由2个平面环组成, 每个平面环由4个亚基组成[14]。Llorca等[10]利用X-ray晶体技术研究菜豆重组GS1结构, 发现其与大豆GS1全酶结构相同, 即由两个4元环聚合体组成的八聚体; 用分析离心机测定菜豆GS1全酶相对分子质量为344 kD, 与晶体研究结果一致。X-ray晶体分析表明玉米GS1、苜蓿GS1全酶是由2个五元环聚合体组成的十聚体[15, 16, 17], 但没有关于GS2全酶结构的报道。
利用凝胶过滤及分析离心机测定纯化GS全酶的分子量, 也可以初步判断GS的聚合状态, 但是所用材料多, 费时长, 仪器设备昂贵, 而且分辨率低。近年来Blue native PAGE (BNE)快速发展[18, 19], 利用凝胶电泳对蛋白质复合体依据其大小进行分离, 具有样品需求少、分辨率高、简便快捷且保持蛋白质聚合状态等特点, 越来越多地应用于活性蛋白的低聚物状态研究[20, 21, 22]。本研究通过改良BNE, 结合蛋白质免疫印迹等技术, 分析了玉米不同组织部位GS的表达特性, 并快速鉴定了GS同工酶的聚合状态, 与玉米GS1晶体研究结果一致, 为简便快捷及时研究GS同工酶表达调控方式及其与玉米氮素利用的关系提供了技术保障。
将玉米品种豫单916种植于河南农业大学农场(郑州), 常规田间管理, 于灌浆期选取根(地下节根和地上气生根)及不同叶位的叶片、叶脉、叶鞘、节、节间、穗柄、苞叶和籽粒, 快速清洗、剪碎, 于液氮中速冻, 置-80℃保存备用。
称取0.5 g不同组织部位的样品, 加液氮研磨, 再加3倍体积的提取缓冲液(100 mmol L-1 Tris-HCl, pH 7.6, 1 mmol L-1 EDTA, 1 mmol L-1 MgCl2和10 mmol L-1 β -巯基乙醇[13, 22])混成匀浆, 冰浴静置30 min后, 4℃、13 000× g离心30 min, 上清液即粗酶提取液。
利用3种凝胶电泳系统分离鉴定GS同工酶。
1.3.1 不连续活性聚丙烯酰胺凝胶电泳系统(Native- PAGE) 由3%的浓缩胶(pH 6.7)和5%的分离胶(pH 8.7)组成, 用于玉米不同组织器官的蛋白提取液中GS同工酶亚型的分离和酶活性检测[22]。4℃预冷电极缓冲液(25 mmol L-1 Tris, 192 mmol L-1甘氨酸), 粗酶液与5× 上样缓冲液[25 mmol L-1 Tris-HCl, pH 7.6, 5% (w/v) β -巯基乙醇, 0.05% (w/v)溴酚蓝, 50% (v/v)甘油]混匀后上样, 于4° C条件下电泳, 浓缩胶稳压80V, 分离胶稳压120 V。
1.3.2 Blue native PAGE (BNE) 依据Wittig等[18]改良BNE电泳方案, 根据GS同工酶全酶大小进行分离并鉴定其相对分子质量。凝胶由3%的浓缩胶和4%~13%的梯度分离胶组成, 样品处理同Native-PAGE, 加入预冷阳极缓冲液(25 mmol L-1咪唑-HCl, pH 7.0), 首先使用阴极电泳缓冲液A (0.02% Coomassie Blue G250, 50 mmol L-1 Tricine-HCl, 5 mmol L-1咪唑, pH 7.0), 于4° C、100 V电泳20min后, 更换为阴极电泳缓冲液B (50 mmol L-1 Tricine-HCl, 5 mmol L-1咪唑, pH 7.0), 继续电泳至样品进入4%~13%的梯度分离胶, 然后进行稳流(15 mA)电泳至蓝色指示剂出胶[22]。
1.3.3 Clear native PAGE (CNE) 参考Wittig等[19], 凝胶体系和BNE一样, 但只使用阴极电泳缓冲液B, 于100V电泳至样品进入分离胶, 然后进行稳流(15 mA)电泳至蓝色指示剂出胶。
胶内GS活性依据Wang等[22]方法检测, 活性染色结束后扫描结果, 然后再用考马斯亮蓝R250染色。BNE胶中, 以高分子蛋白Marker (Amersham HMW Calibration Kit For Native Electrophoresis)为标准, 利用Gel-Pro analyzer计算GS同工酶全酶的相对分子质量。
取适量粗酶提取液与等体积2× 的上样缓冲液[100 mmol L-1 Tris-HCl, pH 6.8, 4% (w/v) SDS, 10% (v/v) β -巯基乙醇, 0.2% (w/v)溴酚蓝, 20% (v/v)甘油]混合, 沸水浴5min变性处理, 室温条件下利用SDS-PAGE (5%浓缩胶, 12%分离胶)分离蛋白质, 将蛋白转移至PVDF膜上进行Western-blot检测。使用本实验室制备的小麦GS多克隆抗体检测玉米GS亚基, 使用Bio-Rad Clarity Western ECL试剂盒显色, 以蛋白Marker (Thermo Scientific PageRuler Prestained Protein Ladder)为标准, 利用Gel-Pro analyzer计算GS同工酶亚基分子质量。
切取BNE胶条, 浸泡于含1% SDS和1% β -巯基乙醇的变性液中, 37℃变性处理2 h[18], 然后用去离子水冲洗凝胶3~5次, 置SDS-PAGE浓缩胶顶部, 室温条件下电泳。电泳结束后, 将蛋白质转移至PVDF膜上进行Western blot分析。
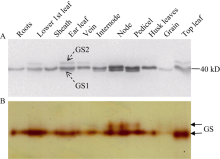
Western blot结果显示玉米中有3种GS亚基, 即40 kD和39 kD的胞液型GS (GS1)及44 kD的质体型GS (GS2), 且不同组织器官中GS同工酶表达差异较大(图1-A)。40 kD的GS1在玉米中起主导作用, 不同组织器官均有较大的表达量(籽粒除外); 39 kD的GS1仅在穗位节间、节和果穗柄中表达, 且后2个部位表达量较高, 可能与玉米灌浆期氮素营养的运输有关; GS2只在功能叶的叶片、叶脉中少量表达, 可能与玉米是C4植物, 光呼吸强度低有关。Native-PAGE结合胶内GS酶活性分析显示(图1-B), 玉米根系、叶片、茎节、穗柄等组织中均检测到GS活性带, 叶片中最高, 籽粒中最低; 但叶片提取液中仅检测到1条GS活性带, 可能因GS2活性太低或不连续Native-PAGE分离蛋白质的特点导致的2种GS同工酶没有被分离开; 此外与Western blot结果一致, 在含两种GS1亚基的穗位节和果穗柄中检测到2个GS同工酶。
 | 图1 玉米不同组织GS同工酶表达和活性分析(A) Western blot鉴定GS同工酶亚基的表达。12% SDS-PAGE分离玉米蛋白, 转膜后利用小麦GS多克隆抗体检测GS亚基。(B)胶内GS酶活性检测。以5% Native-PAGE分离组织蛋白, 胶内GS酶活性分析检测GS同工酶活性。Roots: 根; Lower 1th leaf: 穗下第1叶; Sheath: 穗位叶鞘; Ear leaf: 穗位叶; Vein: 穗位叶叶脉; Internode: 穗位节间; Node: 穗位节; Pedicel: 果穗柄; Husk leaves: 苞叶; Grain: 籽粒; Top leaf: 顶部功能叶。Fig. 1 Analysis of the expression and activity of GS isoforms from different tissues of maize(A) Expression of GS isoforms detected by Western blot. Protein extracts separated by 12% SDS-PAGE, and then probed with GS ployclonal antibodies against wheat after electrobloting. (B) In-gel detection of GS activity. Proteins from different tissues were separated by 5% Native-PAGE, and GS isoforms were detected with the in-gel GS activity analysis. |
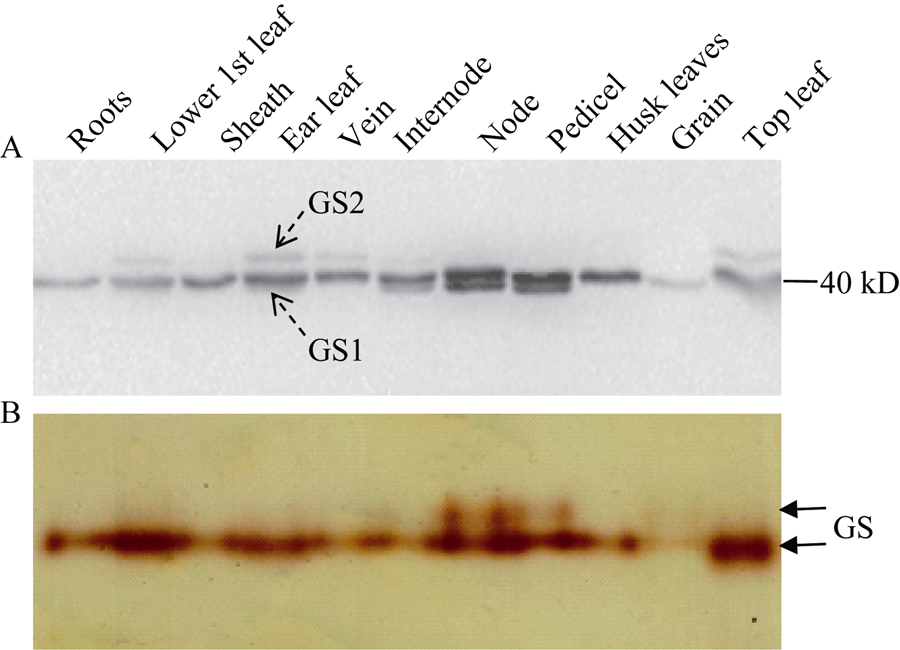
通过改良BNE方法, 在玉米叶片和穗位节间、节和果穗柄中检测到2个大小不同的GS活性带, 其他组织中则只检测到一条GS带(图2-A)。其中, 迁移率较大的GS同工酶全酶分子量约240 kD, 活性较低; 迁移率较小的GS同工酶分子量约410 kD, 活性非常高。利用BNE分离玉米叶片总蛋白后, 进行Western blot, 检测到一个相对分子质量约460 kD的GS同工酶(图2-C), 可能因其活性低, 且分子量接近RuBP羧化酶, 其活性被羧化酶的蓝色条带遮盖[22]。
 | 图2 玉米GS同工酶全酶大小鉴定(A)以BNE分离玉米不同组织可溶蛋白, 以胶内转移酶活性检测GS同工酶。(B) BNE胶考马斯亮蓝染色, 参照蛋白marker, 利用Gel-Pro analyzer计算GS同工酶全酶大小。(C)以BNE/CNE分离玉米穗位叶可溶蛋白, 转膜后进行Western blot检测。Fig. 2 Identification of the molecular weight of GS holoenzymes(A) Soluble proteins from different tissues of maize were separated by BNE, and GS isoforms were detected with an in-gel transferase activity assay. (B) CBB-staining after BNE, the molecular weight of GS holoenzymes were estimated using Gel-Pro analyzer with the calibration of the protein marker. (C) Proteins from the ear leaf of maize were separated by BNE/CNE, and GS isoforms were detected by Western blot. |
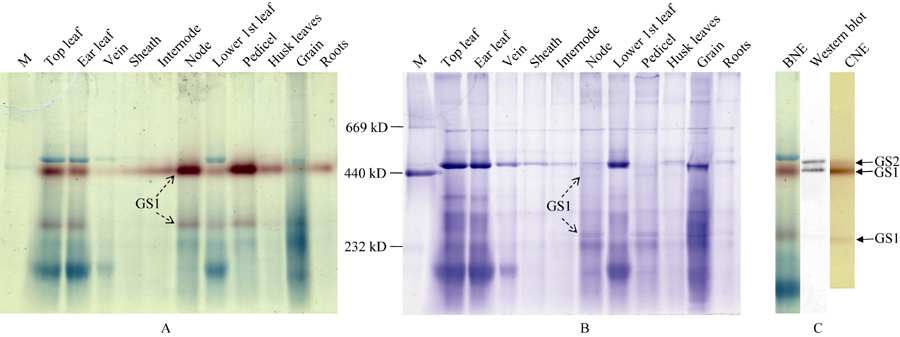
第一相胶为BNE, 用以分离玉米组织蛋白复合体, 切取相应组织的泳道进行蛋白质变性处理; 第二相胶为SDS-PAGE, 对BNE上的蛋白质复合体的亚基进行电泳分离; 然后转膜, 利用Western blot鉴定玉米组织中GS同工酶的亚基组成。玉米叶片中分子量约460 kD的GS同工酶(图2-A)由44 kD的亚基组成且信号非常弱(图3-A), 为GS2; 分子量约410 kD和240 kD的GS同工酶由39~40 kD的亚基组成且信号非常强, 为GS1, 表明胞液型GS存在两种不同的聚合方式。玉米穗柄中只有分子量约410 kD和240 kD的GS同工酶, 也是由39~40 kD的亚基组成, 说明穗柄中GS1全酶也存在两种不同的聚合方式。
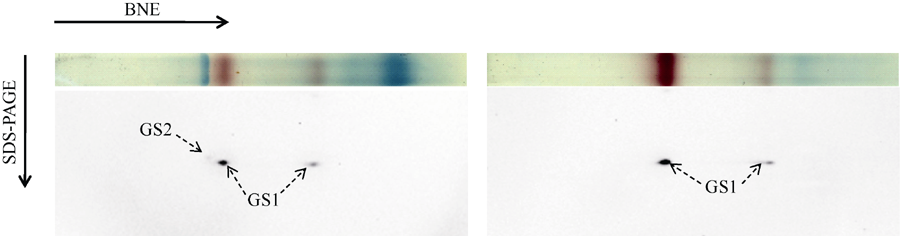 | 图3 玉米GS同工酶亚基组成的鉴定(A)玉米穗位叶GS同工酶亚基组成鉴定, 利用BNE分离叶片可溶蛋白, 切取相应泳道进行变性处理, 利用SDS-PAGE分离复合体亚基, Western blot检测GS亚基。(B)穗柄中GS同工酶亚基鉴定, 方法步骤同(A)。Fig. 3 Identification of the subunit composition of GS isoenzymes in maize(A) Identification of the subunit composition of GS isoenzymes in the ear leaf of maize. Proteins were separated by BNE, the corresponding gel lane was cut out and denaturalized, and the protein complexes were separated by SDS-PAGE, then the GS subunits were identified by Western blot. (B) Identification of the subunit composition of GS isoenzymes from the pedicel of maize, the procedure was the same as that of (A). |
利用BNE计算GS同工酶的全酶大小(图2), 利用BNE与SDS-PAGE结合进行两相电泳分离GS同工酶亚基, 利用Western blot检测鉴定GS同工酶亚基组成(图3)。利用GS同工酶分子量除以相应亚基的分子量, 在此基础上计算GS同工酶聚合状态, GS2为十聚体, 胞液型GS1有2种聚合方式, 一种为十聚体, 和前人结果一致[15]; 此外, GS1同工酶还存在另外一种聚合状态, 即五聚体。
高等植物2种GS同工酶分别定位于细胞液(GS1)和质体(GS2)[2, 23]。GS2缺失在正常条件下是致死性突变; 但通过抑制光呼吸, GS2突变体能够正常生长发育, 证明GS2参与光呼吸过程中的氨同化[2]。C3植物和C4植物光呼吸强度差异巨大[24], 小麦、水稻等C3植物光呼吸强度很高, GS2表达量更丰富[25], 而玉米等C4植物光呼吸强度要小得多, 本研究结果显示仅在玉米叶片中检测到GS2少量表达, 远远低于GS1表达量, 与C4植物光呼吸弱的生理现象一致。
玉米GS1基因由5个核基因编码, 其表达因组织、叶龄及氮量而异, Gln1-3和Gln1-4在叶片表达量高[26], Gln1-3决定穗粒数, Gln1-4调控粒重[13]。本研究发现玉米的穗位节、节间和穗柄组织中有2种高表达GS1亚基, 其中39 kD的GS1亚基是组织特异表达。推测39 kD的GS1可能在玉米灌浆期氮素营养转运方面起着重要作用。
由于从植物组织分离纯化足量蛋白的难度非常大, 因而, 通常采用异源重组植物GS研究其结构和聚合方式[10, 15, 16, 17]。BNE/CNE电泳技术用于分离膜蛋白等蛋白质复合物、测定天然蛋白复合体的相对分子质量, 且具有极高的分辨率[18, 19]。近年来广泛运用于植物可溶性蛋白的分离鉴定[20, 21, 22]。本研究通过改良BNE电泳技术分离玉米可溶性蛋白, 结合胶内活性测定、2-D胶和Western blot技术, 首次发现玉米叶片有3种分子量不同的GS同工酶, GS2为十聚体, GS1主要是十聚体, 与Unno等[15]利用晶体学X-ray分析的玉米GS1a是由2个五聚环组成的十聚体的结果一致; 此外, GS1还存在少量的五聚体, 推测玉米GS1的聚合状态也是一种在氮素代谢中广泛存在的GS同工酶调控方式。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|