
{kind=link}
{kind=link}
玉米大斑病广谱抗性外引自交系的发掘与抗病基因初步鉴定
[肖明纲1, 2  , 宋凤景
, 宋凤景3 , 孙兵2 , 左辛4 , 赵广山5 , 辛爱华2 , 李柱刚2, * ]
, 宋凤景]
|
|


作者简介: E-mail: xiaoyang8076@163.com
从2014—2016年连续3年对43份来自美国、法国、俄罗斯和德国的玉米资源进行了抗大斑病人工接种鉴定, 筛选到高抗玉米大斑病材料7份, 抗病材料1份, 中抗材料6份, 抗性材料占鉴定总材料的比例为32.6%。利用F2群体, 对7份高抗材料进行了大斑病抗性遗传分析, 抗感植株分离比例和适合性测验证明, 自交系A04、F02、F05和R01对大斑病的抗性可能是由一对显性基因控制的。抗谱分析表明, 自交系A04、F02、F05和R01携带的抗大斑病基因不同于 Ht1、 Ht2、 Ht3和 HtN, 可能是新的抗病基因。该研究结果可为今后我国玉米大斑病抗性种质的引进及改良提供重要参考。
, SONG Feng-Jing
From 2014 to 2016, forty-three maize inbred lines introduced from the United States, France, Russia and Germany, were identified and evaluated for resistance to northern corn leaf blight (NCLB). The results showed that seven, one and six maize inbreds were highly resistant, resistant or moderately resistant to NCLB, accounting for 32.6%. Exserohilum turcicum was used to test the F2 populations for analyzing the inheritance of NCLB resistance in the seven highly resistance maize inbred lines. Genetic analysis suggested that A04, F02, F05, and R01 of these inbred lines probably carried a single dominant gene conferring their resistance to NCLB. The reaction patterns to isolates 0, 1, 2, N, and 123N of E. turcicum showed that new genes resistance to NCLB were most likely present in A04, F02, F05, and R01. The results provided useful information on the introduction of maize germplasms and genetic improvement for resistance to NCLB.
玉米大斑病(northern corn leaf blight, NCLB)是由大斑凸脐蠕孢(Exserohlium turcicum Pass. Leonard et Suggs)侵染引致的玉米生产中一种重要的叶部病害, 在我国主要分布于东北地区、华北北部和西南地区等气候较凉爽的玉米种植区, 常造成大面积减产[1]。20世纪80年代, 由于感病杂交种的淘汰, 有效遏制了大斑病在我国的大范围流行[2, 3]。20世纪90年代, 因抗病杂交种的大面积推广种植, 病原菌产生了新的生理小种, 大斑病危害再次升级[4]。2002年以来, 大斑病危害呈回升趋势, 2003— 2006年在东北春玉米区大斑病大范围严重发生, 对生产造成较大影响[5]。因气候条件适宜, 病原菌致病力分化, 感病品种及其相似品种的大面积种植, 2012— 2017年玉米大斑病连续重度发生, 对玉米生产造成严重影响。
迄今, 已报道的玉米抗大斑病主效基因有10个, 被分别定位在第1、第2、第3、第7和第8染色体上, 其中Ht1、Ht2、Ht3、HtN、HtM、HtP、NNc和St为显性单基因, ht4和rt为隐性基因[6, 7, 8, 9, 10, 11, 12]。Ht1基因已经广泛应用于国内外的育成品种中, 在20世纪80年代, 我国利用抗大斑病自交系Mo17为亲本选育出一批抗病杂交种, 有效遏制了大斑病在我国的大范围流行。
在我国东北地区、华北北部和西南等玉米种植区, 凉爽的气候条件利于玉米大斑病流行爆发而造成大面积减产[1, 2, 3, 4, 5]。培育和种植抗病品种是控制玉米病害、减少产量损失的有效途径, 而优异的抗性资源则是进行玉米抗病育种的基础。目前玉米大斑病在我国各玉米区发生越来越普遍, 危害也越来越严重, 针对这种情况, 我们利用人工接种方法, 对引自美国、德国、法国和俄罗斯的43份玉米种质资源进行抗性鉴定和评价, 筛选抗性资源, 探究其真实抗性水平, 为我国玉米抗大斑病育种提供基础材料和抗性信息; 利用F2群体对高抗大斑病材料进行了初步的抗性遗传分析, 利用生产上的优势小种和强致病力小种进行了抗谱分析, 为进一步定位和利用其抗大斑病基因奠定基础。
43份外引自交系, 其中美国30份、法国6份、俄罗斯4份、德国3份; 自交系Mo17 (MR)和获白(HS)为抗感对照。
大斑凸脐蠕孢(Exserohilum turcicum), 分离自黑龙江省玉米发病叶片。
玉米大斑病抗性鉴定圃设置在位于哈尔滨市民主乡的黑龙江省农业科学院耕作栽培研究所实验基地。每份材料种植2行, 行长5 m, 行距0.65 m, 每行留苗25~30株, 株距略小于大田生产, 田间正常管理。2015年和2016年对2014年鉴定得到的抗病材料进行重复鉴定, 以探究其真实抗性。
1.4.1 玉米大斑病接种方法 将菌种接种于经高压灭菌的高粱粒上, 在23~25℃黑暗条件下培养至菌丝布满高粱粒, 用水洗去高粱粒表面菌丝体, 摊铺于洁净瓷盘中, 保持高湿度, 在室温和黑暗条件下培养。镜检确认大量产生分生孢子后, 直接用水淘洗高粱粒, 配制接种悬浮液。待玉米展13叶期时利用孢子悬浮液喷雾接种, 乳熟后期调查[1]。
1.4.2 玉米大斑病抗性评价 调查每份鉴定材料群体的发病状况, 调查重点部位为玉米果穗的上方叶片和下方3叶。病斑占叶面积少于或等于5%为1级(HR), 占叶面积6%~10%为3级(R), 占叶面积11%~30%为5级(MR), 占叶面积31%~70%为7级(S), 全株叶片基本为病斑覆盖, 叶片枯死为9级(HS)[1]。
用7份高抗外引材料分别与感病自交系获白杂交, 产生F2分离群体, 在玉米13叶期时人工接种相应的F2群体和感病对照获白, 乳熟后期调查每个F2植株玉米果穗的上方叶片和下方3叶的总体发病状况[1]。计算各群体抗、感个体的分离比例, 用SAS 8.2软件(SAS Institute, Raleigh, NC, USA) 计算分离比例的χ 2值和概率值, 进行分离比例的适合性测验。
1.6.1 幼苗的准备 种子经2%的次氯酸钠表面消毒处理后, 在25℃下催芽3~4 d。待萌动后, 分别播于10 cm× 10 cm 的花盆内, 每盆播种4粒, 出苗后每盆保苗3株[13]。
1.6.2 接种液制备 将生理小种0、1、2、N、123N纯培养接种到高粱粒培养基上扩繁, 待产生分生孢子后用纯净水冲洗制成孢子悬浮液, 孢子浓度1× 105~1× 106个 mL-1, 滴加0.1%吐温振荡摇匀即可用于接种[13]。
1.6.3 人工喷雾接种 待幼苗长至8~10叶期时进行喷雾接种。接种后保湿24 h, 然后温室进行正常管理, 室内温度控制在25℃左右。接种2周后进行发病调查[13]。
1.6.4 病斑调查 R型: 条状萎蔫褪绿斑或褐色窄条形的坏死斑, 有时有黄色晕圈, 不产生或很少产生孢子; S型: 梭状大型斑, 中间明显坏死, 病斑周围有明显的萎蔫中毒区, 条件适宜时产生大量褐色霉层[13]。
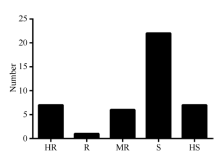
2014年利用孢子悬浮液喷雾接种法对43份外引自交系进行了抗大斑病人工接种鉴定, 感病对照获白病斑占叶片总面积的比率为90%, 而抗病对照Mo17病斑占叶片总面积的比率为15%, 抗感对照抗病性差异显著, 能够有效区分待鉴定材料的真实抗性, 表明人工接种鉴定结果有效。2014年, 从43份鉴定资源中共筛选到高抗材料7份, 抗病材料1份, 中抗材料6份, 高感材料7份, 感病材料22份(图1)。
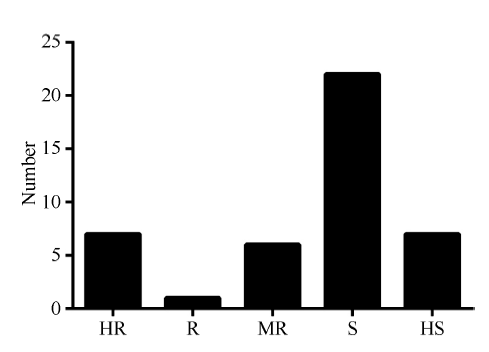 | 图1 外引材料2014年大斑病抗性级别分布 HR: 高抗; R: 抗; MR: 中抗; S: 感; HS: 高感。Fig. 1 Distribution of northern corn leaf blight resistance score for foreign maize inbred lines in 2014 HR: highly resistant; R: resistant; MR: moderately resistant; S: susceptible; HS: highly susceptible. |
从上述鉴定结果可以看出, 引进的43份国外资源, 在大斑病抗性方面具有广泛的变异, 来源于美国的自交系A04高抗大斑病, 来源于德国的自交系G03高感大斑病(图2), 其中的抗性材料对丰富我国玉米抗大斑病种质资源、拓宽其遗传基础具有重要利用价值。
 | 图2 外引材料高抗和高感大斑病田间表型 A04、G03: 自交系; HR: 高抗; HS: 高感。Fig. 2 Field phenotype of foreign maize inbred lines highly resistant and highly sensitive to northern corn leaf blight A04, G03: inbred lines; HR: highly resistant; HS: highly susceptible. |
2015年和2016年对14份抗大斑病材料, 又进行了连续2年的重复鉴定, 感病对照获白病斑占叶片总面积的比率分别为90%和95%, 抗病对照Mo17病斑占叶片总面积的比率分别为25%和20%, 抗感对照充分发病, 人工接种鉴定有效。结果发现, 这些材料年际间大斑病抗性表现稳定, 反应了其真实抗病性, 具体鉴定结果如表1所示。这些材料可作为重要的抗大斑病资源, 用于改善我国玉米抗大斑病种质遗传基础狭窄的现状。
| 表1 14份外引材料对玉米大斑病多年抗性鉴定 Table 1 Identification of resistance to northern corn leaf blight of 14 foreign inbred lines across years |
7份供试自交系对大斑凸脐蠕孢的反应型均表现为高抗, 而感病亲本获白则为高感。根据各供试材料与获白配制的杂种F2植株对大斑凸脐蠕孢的反应型, A04/获白、F02/获白、F05/获白和R01/获白这些F2群体抗、感植株比例都符合3∶ 1的分离比, 而A11/获白、A24/获白和G02/获白的F2群体抗、感植株比例不符合3∶ 1的分离比(表2), 说明A04、F02、F05和R01对大斑凸脐蠕孢的抗性可能受1对显性单基因或少数主效基因控制。
| 表2 7个F2群体对大斑凸脐蠕孢的抗性反应 Table 2 Reactions of seven F2 populations to Exserohilum turcicum of northern corn leaf blight |
遗传分析表明, 自交系A04、F02、F05和R01对玉米大斑病的抗性均受1对显性单基因控制。因此, 利用当前优势小种0和1号、2号生理小种、N号生理小种及强致病力小种123N对A04、F02、F05和R01进行了抗谱分析。生理小种0、1、2、N及123N对A04、F02、F05和R01均无毒性, 而强致病力小种123N对Ht1、Ht2、Ht3和HtN均有毒性, A04、F02、F05和R01对4个生理小种的反应型均不同于Ht1、Ht2、Ht3和HtN的反应型(表3)。说明自交系A04、F02、F05和R01对大斑病的抗性不同于已定位的抗大斑病基因Ht1、Ht2、Ht3和HtN, 可能携带新的抗病基因。
| 表3 4份玉米自交系对0、1、2、N和123N的反应型 Table 3 Infection types in the four foreign inbred lines to isolates 0, 1, 2, N, and 123N of northern corn leaf blight |
玉米对大斑病的抗性有两种遗传类型, 即质量遗传抗性和数量遗传抗性, 这两种抗性均广泛应用于玉米抗病育种中[11, 14]。
质量抗性, 也叫小种专化抗性, 是受显性单基因控制的垂直抗性。质量抗性基因在控制玉米大斑病危害方面取得过显著成效[15]。但长期大面积种植同一抗源选育的品种, 由于病菌与寄主互作具有普遍的基因对基因关系, 很容易使有效的抗病基因随着病菌对定向选择压力的适应性变异而失去作用, 形成所谓的“ 抗性” 丧失现象[5, 15, 16]。因此, 寻找更多、更有效、更易利用的抗病自交系或基因资源是当前我国玉米抗病育种的一项十分紧迫的任务。
自交系A04株型清秀, 年际间抗性稳定, 病斑面积占总叶片面积的比率为1%, 其最大特点是脱水快、适宜直收; F02和F05株高和穗位较低, 粒深轴细, 产量高, 年际间病斑面积占叶片总面积的比率有波动, 最高分别为3%和2%; R01抗低温能力较强, 年际间抗性稳定, 病斑面积占叶片总面积的比率为4% (表1)。以病情级别为唯一标准, 对A04、F02、F05和R01进行抗性遗传分析和多小种鉴定, 结果表明, A04、F02、F05和R01对大斑病的抗性受一对显性单基因控制(表2), 且可能携带有新基因(表3)。对A04、F02、F05和R01携带的抗病基因精细定位、克隆和功能验证, 以丰富我国玉米抗大斑病基因资源, 开发与抗病基因紧密连锁的分子标记, 利用分子标记辅助选择及回交转育等手段, 提高我国当前玉米生产上常用种质抗病性的同时, 也可改善目标材料的脱水性、耐冷性, 对于培育高产广适品种和资源创制具有重要指导意义。
数量抗性, 属水平抗性, 在遗传上受多基因控制, 对所有大斑病菌的生理小种均有效。大斑病数量抗性基因分布在玉米12条染色体上的整个基因组[11, 17, 18], 因其抗性持久且不易被新的生理小种克服而受到广泛重视[19]。通过构建不同的群体材料, 许多玉米抗大斑病QTLs位点被定位[20, 21, 22, 23], qNLB8.06DK888被精细定位在第8染色体0.46 MB的区间内, qNLB1.06被限定在3.60 MB的区间内[24, 25]。
A11和A24具有配合力高、耐密植等优点, 高抗大斑病且抗性稳定, 年际间叶片上病斑面积占叶片总面积的百分比分别为5%和0; G02硬粒型, 秆韧性好, 抗倒性极强, 高抗大斑病且抗性稳定, 年际间叶片上病斑面积占叶片总面积的百分比有波动, 分别为0、1%和0 (表1)。以病情级别为指标, A11/获白、A24/获白和G02/获白的F2群体抗、感植株比例既不完全符合3∶ 1的分离比, 也不符合15∶ 1的分离比例(表2)。如果综合考虑病情级别、病斑大小和年际间环境因素的影响, A11和G02对大斑病的抗性很有可能受数量性状控制。但A24三年鉴定没有发现病斑, 年际间叶片上病斑面积占叶片总面积的百分比均为0, 其抗性遗传规律有待做进一步的深入研究, 以期发掘具有持久抗性且耐密、抗倒的玉米优异资源。
尽管质量抗性基因和数量抗性基因有着明显的不同, 但是在某些情况下, 质量抗性基因更像是一个主效QTL[26, 27]。F02和F05年际间叶片上病斑面积占叶片总面积的百分比有波动, 分别为1%、3%、3%和2%、1%、1%, 而A04和R01年际间叶片上病斑面积占叶片总面积的百分比比较恒定, 分别为1%和4%(表1), 但病斑数量、病斑长度和宽度可能有差异。仅根据病情级别, A04、F02、F05和R01对大斑病的抗性可能受一对显性单基因控制, 而如果综合考虑病情级别、病斑大小、环境因素及多小种鉴定结果, A04、F02、F05和R01对大斑病的抗性也可能受少数主效QTL控制, 存在微效基因的作用。质量单基因或主效QTL定位将是下一步工作重点。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|