
{kind=link}
{kind=link}
{kind=link}
利用9311来源的粳型染色体片段代换系定位控制稻米糊化温度的微效QTL
[刘鑫燕1, 2 , 朱孔志1 , 张昌泉1 , 洪燃1 , 孙鹏1 , 汤述翥1 , 顾铭洪1 , 刘巧泉1, *  ]
]
]
|
|


糊化温度(gelatinization temperature, GT)是评价稻米蒸煮与食味品质的重要因素之一, 除受一主效基因控制外, 还受多个微效基因的影响。本研究利用粳稻品种日本晴和籼稻品种9311作为受体和供体来源的38个染色体片段代换系为研究对象, 于2010—2011年连续2年分别于2个环境内种植, 测定各株系稻米的糊化温度(碱消值), 利用
Gelatinization temperature (GT), one of the determinants for rice cooking and eating quality, is controlled by not only a major gene but also several minor genes. Previously, we used the
随着人们生活水平的提高, 稻米品质日趋受到重视。糊化温度(gelatinization temperature, GT)是指大米淀粉蒸煮后淀粉粒发生不可逆转膨胀、双折射现象消失时的温度, 它是评价稻米蒸煮与食味品质的一项重要指标[ 1]。GT一般分为低( <70℃)、中(70~ 74℃)和高等( >74℃) 3个等级[ 2]。因测定困难, 通常用碱消值(alkali spreading value, ASV)间接表示。ASV的6~7级对应于低糊化温度, 3~5级对应于中糊化温度, 1~2级对应于高糊化温度。稻米糊化温度直接反映煮饭时米的吸水率、膨胀容积和伸长程度。一般高糊化温度的稻米相对于低糊化温度的稻米需更多的水分及更长的蒸煮时间。由于育种家的定向选择, 目前主要栽培品种多为中、低GT类型, 少数为高GT类型; 大多数粳稻品种属于低GT类型, 少数为中GT类型, 很少有高GT的粳稻品种。
糊化温度的遗传, 多数研究认为是受一主效基因和若干微效基因共同控制。日本学者Kudo等[ 3]最早通过细胞遗传学将控制GT的主效基因定位在水稻第6染色体上。He等[ 4]对DH群体的QTL分析显示, GT主要受第6染色体的 ALK主基因及该染色体上一个微效QTL控制。利用KDML105和CT9993衍生的重组自交系群体, Lanceras等[ 5]检测到1个位于第2染色体上的QTL和2个第6染色体上的QTL共同控制GT。严长杰等[ 6]利用GT上具有显著差别的籼稻品种南特号和粳稻品种Balilla杂交, 并与Balilla回交得到的142株回交群体为研究对象, 通过SSR标记定位了一个第6染色体上的主效QTL (即 ALK位点)和分别位于第2、第3、第6、第9和第11染色体上的5个微效QTL。纵观前人研究, GT除受第6染色体上的主效QTL (即 ALK位点)控制外, 还有一些微效QTL的影响。高振宇等[ 7]利用图位克隆的方法, 分离克隆了控制水稻GT的主效基因 ALK, 序列分析表明其编码可溶性淀粉合酶IIa (SSSIIa); 同时研究了不同品种中该基因DNA序列与碱消值之间的关系, 推测出糊化温度的高低与该基因编码区序列中ATG下游G264变为C264有关, 这一碱基变换使所在密码子由编码谷氨酸变为天冬氨酸, 是导致糊化温度降低的主要原因。对GT微效QTL的研究, 在不同群体中所获结果有所不同, 其中主要的原因是传统的群体如杂种F2/F3、低世代回交群体(BC1)、加倍单倍体(DH)和重组自交系(RIL)等初级作图群体有众多的遗传因子同时分离, 个体间遗传背景相差很大, 难以准确鉴别单个 QTL的效应, 克隆就更困难[ 4, 5, 6]。为克服初级群体在QTL研究中的缺点, 一系列的高级作图群体(次级群体), 如导入系(introgression lines, Ils)、代换系(substitution lines, SLs)或近等基因系(near isogenic lines, NILs)等被发展, 并已被用于稻米品质性状相关微效QTL的定位[ 8]。
本实验室在前期研究中, 分别以全基因组测序的粳稻品种日本晴和籼稻品种9311为受体和供体, 构建了一套染色体片段代换系群体, 并借助高通量全基因组重测序的方式对所有染色体片段代换系进行了精确的基因型鉴定[ 9]。高振宇等[ 7]的研究表明, 9311和日本晴2个品种中控制GT的主效基因相同, 即 ALK基因编码区ATG下游第264位碱基都为C; 碱消值表型鉴定也都属C型, 即在碱液中米粒能完全溶解, 均表现为中糊化温度类型。因此, 本研究利用由这2个亲本来源的染色体片段代换系群体对GT进行定位研究, 以期定位控制GT的微效QTL。通过2年各2个环境的数据分析, 共定位到4个与GT相关的微效QTL, 其中2个QTL能稳定遗传, 为进一步精细定位并克隆相应QTL奠定了基础。
2010年和2011年籼稻( O. sativaL. subsp. indica)品种9311和粳稻( O. sativaL. subsp. japonica)品种日本晴(Nipponbare), 以及以日本晴为受体、9311为供体构建的38个经重测序的染色体片段代换系群体[ 9]于扬州大学文汇路校区内试验基地(简称为“校内”)和扬州市郊杭集镇的试验基地(简称为“杭集”)种植, 每个环境设置3个重复。每个系种植4~6行, 每行10个单株, 株行距分别为13 cm和17 cm。均按常规水肥管理。38个染色体片段代换系中的代换片段分布于水稻的12条染色体上, 覆盖了水稻全基因组的87.46%[ 9]。染色体代换系群体抽穗期持续时间约为8 d, 灌浆成熟期间的条件较为一致, 抽穗后22~25 d成熟收获。为减少生育期差异对稻米品质的影响, 本研究剔除了个别生育期滞后的家系。
收获亲本及水稻染色体代换系植株上成熟种子, 采用Little等[ 10]方法测定每粒精米的碱消值(ASV), 按1~7级记分。每个株系测定3个单株, 每个单株均测定18粒, 以其平均数代表亲本或代换系各株系的GT表型值。按农业部标准米质测定方法NY147- 88测定稻米中的直链淀粉含量和胶稠度。
GT数据由统计软件SPSS16.0分析完成。采用Eshed等[ 11]的方法稍作修改检测QTL。以受体亲本日本晴为对照, 参照张昌泉等[ 8]方法检测代换系与对照亲本间的差异显著性。通过 t测验比较每个CSSL与日本晴GT之间的差异, 以 α= 0.05为阈值, 即 P≤0.05, 则认为染色体置换片段上存在控制GT的QTL; 当 P> 0.05时则不存在控制GT的QTL。参考Eshed等[ 11]的方法估算各个QTL的加性效应值及加性效应贡献率。加性效应值=(染色体片段代换系的碱消值-日本晴碱消值)/2; 加性效应贡献率 = (染色体片段代换系加性效应值/日本晴碱消值) × 100%。
QTL代换作图参照何风华等[ 12]的方法并作适当修改。即2个或2个以上的染色体代换系在相互重叠的置换区段上均检测到控制GT的QTL, 而且遗传效应一致, 则认为QTL位于置换片段的重叠片段上; 如果在某一个CSSL的置换片段上检测到控制GT的QTL, 但在含有其置换片段某相应重叠区段的另一个或多个CSSL中未检测出, 则认为QTL位于非重叠的区段上。
待测样品的精米用三倍体积NaOH溶液(pH8.0~8.5)浸泡过夜后, 去除浮渣。随后用组织匀浆机(IKA-T RCT-Basic, Germany)以17 500转 min-1匀浆3 min, 匀浆液中按每克精米添加50 mg的碱性蛋白酶, 42℃磁力搅拌24 h, 将消化过的匀浆液过200目筛后弃掉残渣, 滤液经3600 × g离心20 min, 弃上清液, 用去离子水悬浮沉淀后3600 × g离心20 min。重复上述清洗过程5次以去掉淀粉中残留的离子。后用90%无水乙醇洗2次, 于真空干燥器中干燥, 过100目筛提纯出天然的淀粉, 随后用塑料袋密封放置于4℃冰箱备用。参照Zhu等[ 13]的方法将提纯的淀粉去分支, 利用异淀粉酶(EC3.2.1.68, Hayashibara Biochemical Laboratories, Inc, Okayama, Japan)处理提纯亲本及代换系成熟种子收获的淀粉, 获得去分支的淀粉。利用凝胶色谱仪(PL-GPC 220, Polymer Laboratories Varian, Inc. Amherst, MA)分析去分支的淀粉分子量分布。
双亲及染色体片段代换系群体的平均碱消值见表1。 t测验表明受体亲本日本晴和供体亲本9311的碱消值在2年2个地点均存在显著差异( P≤0.05), 且较稳定。双亲及染色体群体在2年2点的碱消值频率分布图见图1, 由图1可知, 染色体片段代换系间GT的变异范围较广, 且呈连续分布, 这也表明GT性状受多基因控制。
| 表1 双亲及染色体片段代换系群体的碱消值 Table 1 Profiling of the alkali spreading value among CSSLs and their two parents |
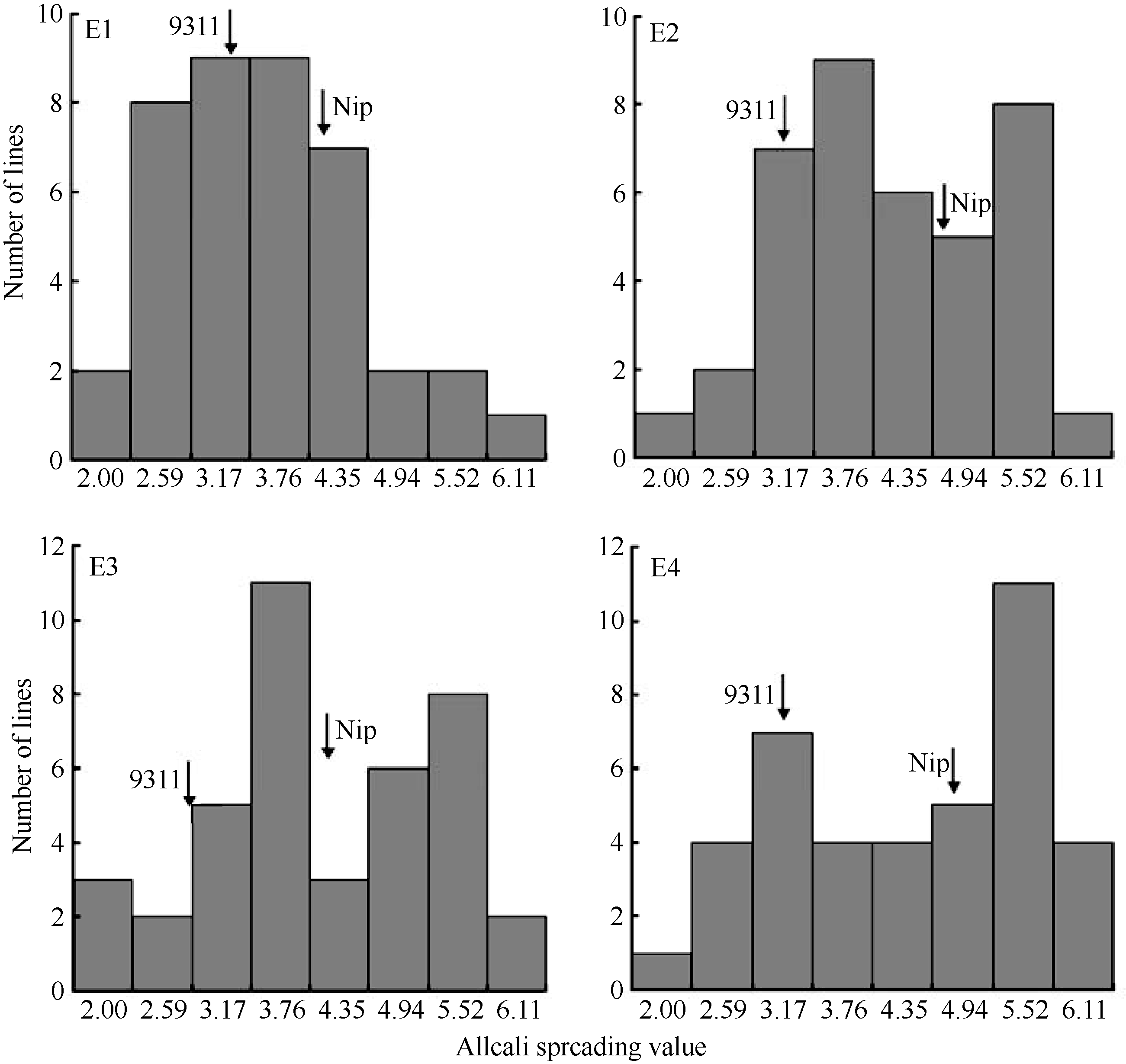 | 图1 双亲及染色体片段代换系在不同环境中的碱消值频率分布E1: 校内(2010); E2: 杭集(2010); E3: 校内(2011); E4: 杭集(2011)。Fig. 1 Histogram of alkali spreading value of CSSLs and their two parents in different environmentsE1: Campus (2010); E2: Hangji (2010); E3: Campus (2011); E4: Hangji (2011). |
为排除环境的影响, 本实验对2年各2个环境的碱消值进行了分析, 以同一年份2个地点都能检出的极显著差异位点作为一个稳定QTL, 共检测到4个QTL, 分别是 qGT2-1、 qGT7-1、 qGT8-1、 qGT12-1 (表2)。其中 qGT2-1和 qGT8-1分别只在2010年和2011年检测到, 在年份间的稳定性较差, 而 qGT7-1和 qGT12-1在2年2地均检测到, 遗传效应的趋势也一致, 是很稳定的2个QTL。在2010年, qGT7-1和 qGT12-1在校内环境下对GT的加性效应值分别为-0.92和-1.00, 相应的加性效应贡献率分别为21.53%和23.53%; 2个QTL的加性效应值之和超出了亲本间的差异, 其原因可能存在着一定的上位性。在杭集试验点2个QTL对GT的加性效应值分别为-0.50和-0.50, 相应的加性效应贡献率分别为11.31%和11.31%。2011年, qGT7-1和 qGT12-1在校内环境下对GT的加性效应值分别为-0.71和-1.38, 相应的加性效应贡献率分别为14.84%和28.95%; 在杭集试验点2个QTL对GT的加性效应值分别为-0.79和-1.13, 相应的加性效应贡献率分别为18.59%和26.47%。 qGT12-1对水稻GT的加性效应贡献率最大, 含有 qGT12-1置换片段的代换系N53所对应的GT为高或中GT。
| 表2 利用染色体片段代换系对水稻糊化温度QTL分析 Table 2 Analysis of QTL for gelatinization temperature in rice CSSL |
本研究所用的38个染色体片段代换系均已全基因组重测序分析, 基因型图谱已发表[ 9]。通过染色体代换片段的重叠分析, 可进一步缩小QTL的定位区间。部分家系的ASV见表3。在代换系N9中检测到 qGT2-1, 初步将其定位在第2染色体上, 约1.10 Mb (图2-A); 在代换系N30中检测到 qGT7-1, 考虑N26和N27重叠部分, 将其定位在第7染色体上, 约13.44 Mb (图2-B, F); 在代换系N49中检测到 qGT8-1, 考虑N46重叠部分, 将其定位在第8染色体上, 约6.28 Mb (图2-G)。在代换系N53中检测到 qGT12-1, 定位在第12染色体中, 约11.04 Mb (图2-D, H)。
家系N53携带来自第4、第8和第12染色体上的3个染色体片段, 其中第8和第12染色体上的2个代换片段正好是 qGT8-1和 qGT12-1所在位置。按上述分析, 本研究定位到的 qGT12-1在2年均稳定存在, 而2011年只检测到 qGT8-1。为进一步分析这2个QTL的效应及其真实性, 分别进行代换作图(图2), 并列出了相关代换系在2年4个环境下的碱消值(表3)。在2010年, N53家系稻米碱消值与受体亲本有极显著差异, 而含有第8染色体相同代换片段的N49家系稻米碱消值与受体亲本无显著差异, 因此认为N53家系中造成GT差异的原因是由于第12染色体上代换片段引起的。在2011年, N53和N49家系稻米碱消值都显著区别于受体亲本, 可以推测53家系中GT的差异可能是2个QTL qGT8-1和 qGT12-1共同作用的结果。
 | 图2 控制稻米糊化温度的QTL代换作图基因型数据基于全基因组重测序, 黑色表示来自9311片段, 灰色表示日本晴背景。右边为CSSL的名称, 带*表示该CSSL稻米GT与受体亲本日本晴相比有显著差异。Fig. 2 Physical mapping of the QTLs for gelatinization temperature with developed CSSLsGenotypes of the CSSLs were constructed based on the whole-genome re-sequencing data. Black color indicates the substituted segments from 9311, are denoted by gray bars from Nipponbare. |
| 表3 几个家系碱消值的测定值 Table 3 Profiling of the alkali spreading value among several CSSLs (mean± SD) |
稻米理化品质是一个综合性状, 各性状间存在复杂的关系。同时对2年各2个环境直链淀粉含量的分析(表4)发现, 控制GT的 qGT2-1和 qGT8-1对AC也有一定的影响, 说明这2个区段存在同时影响GT和AC的QTL, 这些区段所含的QTL可能具有多效性。
为了探究造成GT差异的可能原因, 对受体亲本日本晴、供体亲本9311及与受体亲本碱消值差异最大的代换系N53株系成熟种子的淀粉分子结构进行了分析。利用凝胶渗透色谱仪(GPC)分析了样品提纯淀粉的分子量分布情况, 结果见图3。被酶解的淀粉经GPC分离, 出现了3个明显的峰, 左边2个峰F2和F1分别是被酶解的支链淀粉的一级支链峰和二级支链峰, 其中一级支链峰(F2)表示淀粉的长B链, 而二级支链峰(F1)表示短分支的A链和B1链; 右边第3个峰是未被酶解的直链淀粉。日本晴和9311在二级支链峰差异比较小, 而含 qGT8-1和 qGT12-1代换系N53的支链含量远远小于日本晴, 说明N53相对于日本晴而言, 含有较少的A链和短B链。长链B的组成在3个材料稻米淀粉间也存在差异, 代换系N53的长链B含量远远大于受体亲本日本晴。从图中可见, 代换系N53和日本晴在直链淀粉所对应的峰面积上并没有显著的差异。
| 表4 部分家系及受体亲本胶稠度(GC)、直链淀粉含量(AC)的测定值 Table 4 Gel consistency (GC) and amylase content (AC) in partial substitution lines and recipient parent (mean± SD) |
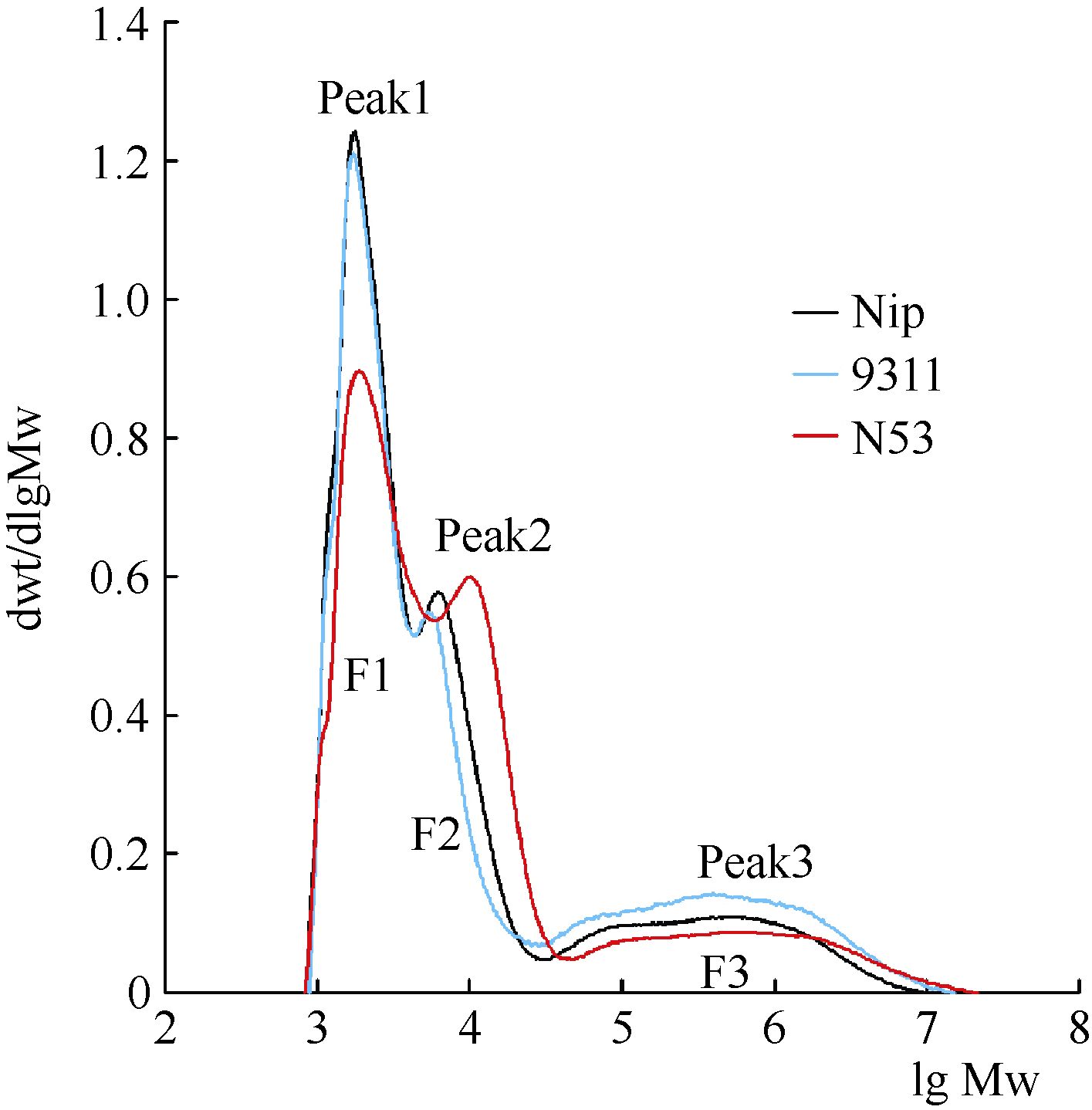 | 图3 受体亲本日本晴(Nip, 黑色)、供体亲本9311 (蓝色)和代换系N53 (红色)稻米淀粉经异淀粉酶去分支后的链长分布lg Mw:分子量的对数; dWt/dlg Mw: 分子量的微分分布。Fig. 3 Chain distributions of isoamylase-debranched starches from Nipponbare (Nip, black), 9311 (blue), and N53 (red) by Gel permeation chromatogramslg Mw: The molecular weight of logarithmic; dWt/dlg Mw: molecular weight distribution of calculus. |
稻米糊化温度因品种类型不同而有很大的差异, 一般糯米低于非糯米、粳米低于籼米, 籼米中还有高、中、低GT三种类型[ 14, 15]。对糊化温度遗传, 普遍认为是由一主效基因和若干微效基因共同控制[ 16, 17]。控制GT基因的分子定位研究开始于20世纪90年代, 此后许多分子遗传研究也都表明籼稻和粳稻间碱消值(ASV)的差异主要是由一主效基因或QTL ( ALK)控制[ 4, 5, 6, 7]。Umemoto等[ 18]利用粳稻日本晴和籼稻Kasalath及其杂交后代将控制GT差异的 ALK基因和 gel(t)基因、控制支链淀粉链长分布的 acl(t)基因和淀粉合酶基因 SSIIa定位在同一位点, 与分子标
记C1478和G200/R2147的遗传距离分别为0.70 cM和0.23 cM。高振宇等[ 7]通过图位克隆法克隆了 ALK基因, 并证明糊化温度的高低与该基因序列中ATG下游外显子中G264突变为C264有关, 这一碱基变换使所在密码子由编码谷氨酸变为天冬氨酸, 是导致糊化温度降低的主要原因。本研究所用的2个亲本 ALK基因序列中都是C264。高振宇等[ 19]通过15个水稻品种对 ALK基因进行了深入研究, 共发现了4个单核苷酸变异。两亲本除了在264位点相同外, 在ALK-SNP2-3797位点上也相同, 均为A。但在ALK- SNP3-3797位点上存在差异, 日本晴为A, 9311为G; 在ALK-SNP4-4327, 4328两位点上, 日本晴为GC, 而9311为TT。但这些差异碱基对糊化温度并没有产生影响[ 20], 说明本研究所用染色体片段代换系的2个亲本中控制GT的 ALK主效基因是相同的。
除主效基因 ALK外, 微效基因/QTL对GT的影响也起着较为重要的作用。Lanceras等[ 5]发现在水稻第2和第6染色体上存在微效作用位点, 分别解释了GT变异的12.22%和8.57%。由于不同的研究者所用的实验材料和分析方法各不相同, 并且性状本身在遗传表达上较复杂, 因此对GT的遗传控制有着不同的解释[ 4, 5, 6, 18]。本研究利用染色体代换系共鉴定出4个与GT相关的微效QTL, 分别为 qGT2-1、 qGT7-1、 qGT8-1和 qGT12-1, 部分QTL与前人报道的结果相同, qGT2-1与Lanceras等[ 5]定位在水稻第2染色体的QTL、严长杰等[ 6]定位到的 qASV2位于相同的染色体区段上, 此位点Govindraj等[ 20]在DH群体也定位到, 能解释GT变异的14.41%。 qGT8-1包含了Fan等[ 21]定位到的2个参与糊化温度控制的上位性位点。本研究中检测出的其他2个与GT相关的QTL尚未见报道。Tian等[ 22]研究发现, 决定稻米蒸煮与食味品质的淀粉合成相关基因通过影响直链淀粉含量、胶稠度和GT形成一个精细的调控网络。本研究也证实了这一观点, qGT2-1和 qGT8-1这2个QTL同时影响着稻米的AC, 具有多效性。有关GT与AC的关系, Radhika等[ 23]报道, 在不同类型的品种中, AAC与GT显著正相关。此外, 利用本群体也进行了RVA分析, 结果发现 qGT7-1与控制米粉的消减值 qSBV7.1位于同一位置[ 8], Bao等[ 24]在该位置也检测到了控制消减值的QTL。
研究表明, 稻米GT是一个复杂的性状。除受品种的遗传因素影响外, 亦受环境因素的影响。本研究所用的CSSL群体中, 共检测到4个影响GT的微效QTL, 其中2个QTL( qGT7-1和 qGT12-1)在不同年份和不同环境下都能稳定表达。进一步对其淀粉的精细结构分析表明, 含有来自9311的 qGT8-1、 qGT12-1的染色体片段代换系稻米淀粉含有较少的A链和短B链, 推测短分支的支链淀粉A链和B1链含量的减少可能会引起GT的升高。Zhu等[ 13]对3个糯稻品种的研究, 也证实了这一推测。扬辐糯的糊化温度比苏御糯和广陵香糯都高, 支链淀粉链长分布结果显示, 扬辐糯含有较多的支链淀粉A链和B1链; Bao等[ 25]提出了SSIIa (即 ALK编码的酶)决定GT高低的可能机制, 认为SSIIa的活性决定了支链淀粉的链长分布, 而链长的分布决定了GT的高低。一般而言, 低GT品种的支链淀粉含有较多的短链, 高GT品种的支链淀粉含有较多的长链。本研究中, 通过凝胶渗透色谱(GPC)对2个亲本的链长分析发现, 2个品种的淀粉经过异淀粉酶处理后, 淀粉低分子量的短分支链差异不大, 可能与两亲本 ALK基因序列差异不大有关, 使定位控制GT的微效QTL更加精确。另一方面, 淀粉合成是一个复杂的网络, 淀粉合成相关基因如控制直链淀粉、胶稠度等基因对支链淀粉的合成也有一定的影响。本研究所用的染色体片段代换系的2个亲本含有相同的 Wxb等位基因, 可排除 Wx基因对GT的影响, 可更加准确地鉴定控制GT的微效QTL。
基于分子标记检测技术, 我们成功构建了染色体片段代换系, 并经全基因组重测序方式进行基因型鉴定。这套遗传群体中代换片段平均长约5.6 Mb, 导入片段覆盖率仅为87.4%, 因此可能还会遗漏部分QTL。因此构建高覆盖率的染色体单片段代换系是本研究的下一步目标, 此项工作正在进行中。另外, 将定位到携带QTL的染色体片段代换系进一步与受体亲本回交, 以期产生分离群体来验证并精细定位相关的QTL。
共鉴定出4个与GT相关的微效QTL, 分别为 qGT2-1、 qGT7-1、 qGT8-1和 qGT12-1, 分别位于水稻第2、第7、第8和第12染色体上。其中 qGT7-1和 qGT12-12个QTL的遗传较为稳定, 其加性效应贡献率为11.31%~28.95%。推测支链淀粉中短链含量的减少可能会引起GT的升高。研究结果为进一步精细定位和克隆相应微效QTL及开展稻米品质改良的分子育种奠定了基础。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|