


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
花后前期高温对玉米强弱势籽粒生长发育的影响
[赵丽晓, 张萍, 王若男, 王璞*  , 陶洪斌]
, 陶洪斌]
, 陶洪斌]
|
|


采用籽粒离体培养的方法, 研究花后高温对玉米强、弱势籽粒的影响。结果表明, 高温处理加快了强、弱势籽粒前期的灌浆速率, 但降低了中后期的灌浆速率, 导致粒重降低, 且对弱势粒影响尤为显著, 高温处理强、弱势粒成熟期粒重分别比对照低5.8%、17.4%; 高温显著降低了籽粒不同灌浆时期的淀粉合成相关酶活性, 从而使淀粉含量降低, 强势粒的淀粉含量降低幅度小于弱势粒; 与对照相比, 高温处理后强势籽粒中的3-吲哚乙酸(IAA)和玉米素核苷(ZR)含量显著下降, 赤霉素(GA3)含量则无显著差异, 而弱势粒IAA、ZR含量显著降低, 但GA3含量增加, 可能是导致弱势粒干重受损较大的原因。
The kernel culture
随着全球气候变暖, 高温胁迫对玉米的影响日益突出。黄淮海地区是我国最大的玉米产区, 吐丝开花期高温多雨的气候条件, 限制了产量的进一步增加[ 1, 2]。花期高温对产量的影响主要是通过影响玉米的花粉活力、花丝生长, 授粉受精过程, 或在籽粒建成期影响单位籽粒库容的形成, 导致穗上部籽粒败育率增加, 有效粒数减少, 且粒重降低[ 2, 3, 4, 5]。但不同阶段高温使玉米受损程度不同[ 6], 花后高温对玉米产量、叶片光合速率和抗氧化能力等的影响大于花前高温[ 7]。对于花后高温对籽粒生理生化的影响
已有大量研究, 但玉米穗部不同籽粒对高温的响应研究较少。因此, 本试验采用玉米籽粒离体培养方法, 在人工控温条件下, 研究玉米强、弱势籽粒对花后高温的响应机制, 为生产上提出有效的防控措施提供理论依据。
2013年在中国农业大学上庄试验站种植郑单958, 田间管理同常规田。在玉米吐丝前, 选取生长健壮、长势均匀一致的植株套袋, 人工辅助统一授粉。授粉后3 d取穗进行玉米籽粒体外培养, 试验分中部和顶部籽粒进行取样, 从顶部向下1 cm为弱势粒取样位, 从基部第20位籽粒为强势粒取样位。
籽粒授粉后第4天在昼夜25℃/25℃下暗培养1 d, 第5天开始温度处理。高温处理为昼夜35℃/25℃ (12 h/12 h), 对照为25℃/25℃。高温处理时间为5~15 DAP (days after pollination), 随后转入25℃/25℃培养至成熟。Q-35、Q-25分别指高温处理、对照的强势粒, R-35、R-25分别指高温处理、对照处理的弱势粒。共4个处理, 每个处理3次重复, 每个重复至少含12颗籽粒。
参考Gengenbach和Jones[ 8]方法, 按籽粒穗轴比为1∶9切取样穗[ 9], 经75%酒精灭菌后放入超净台进行操作。
参见Cully和Gengenbach[ 10]方法, 稍作改动, 培养基中氨基酸组分和浓度(mg L-1)为谷氨酸627.5、蛋氨酸42.5、丙氨酸170、酪氨酸92.5、组氨酸57.5、苏氨酸70、脯氨酸222.5、精氨酸52.5、色氨酸72.5、丝氨酸122.5、甘氨酸60、缬氨酸117.5、亮氨酸360、赖氨酸32.5、谷氨酰胺105、苯丙氨酸137.5、异亮氨酸85、半胱氨酸57.5、天冬氨酸177.5、天冬酰胺107.5。将籽粒转入培养基后, 放入人工气候培养箱中恒温培养前, 使用内置紫外线灯灭菌20 min。
1.3.1 籽粒干、鲜重 在授粉后10、20、30、40和50 DAP取样, 测定籽粒的鲜重, 105℃杀青30 min, 80℃烘干, 测定干重。
1.3.2 籽粒淀粉含量 将烘干后的籽粒研磨, 参照何照范[ 11]方法测定淀粉含量。
1.3.3 淀粉合成相关酶活性 在15、20、25、30、40 DAP取鲜籽粒液氮冷冻后置-40℃冰柜中, 用于淀粉合成相关酶活性的测定, 酶的提取和测定参照Nakamura和Zinselmeier[ 12, 13]的方法, 稍作改动。
1.3.4 激素含量 在5 DAP和15 DAP取鲜籽粒液氮冷冻后置超低温冰箱中, 用于激素含量测定, 参照宋松泉[ 14]的酶联免疫法, 酶联免疫试剂盒由中国农业大学提供。
采用Microsoft Excel 2007和SAS 8.0 GLM (General linear model) 程序统计分析数据。
花后高温对强、弱势粒干重的影响呈现相同的变化趋势(图1)。在灌浆前期, 高温处理籽粒的干重均高于对照, 但随着培养时间的增加, 干重逐渐小于对照。高温加快了籽粒灌浆前期的干重积累, 在10 DAP, Q-35、R-35的干重分别比对照高23.4%和22.3%, 但降低了成熟期的籽粒干重, Q-35的干重在50 DAP比对照低5.8%, R-35低17.4%, 弱势粒干重受高温影响程度大于强势粒。此外, 由图1可见, 离体培养的强、弱势籽粒, 在充足的营养供应下有相似的干重积累趋势。
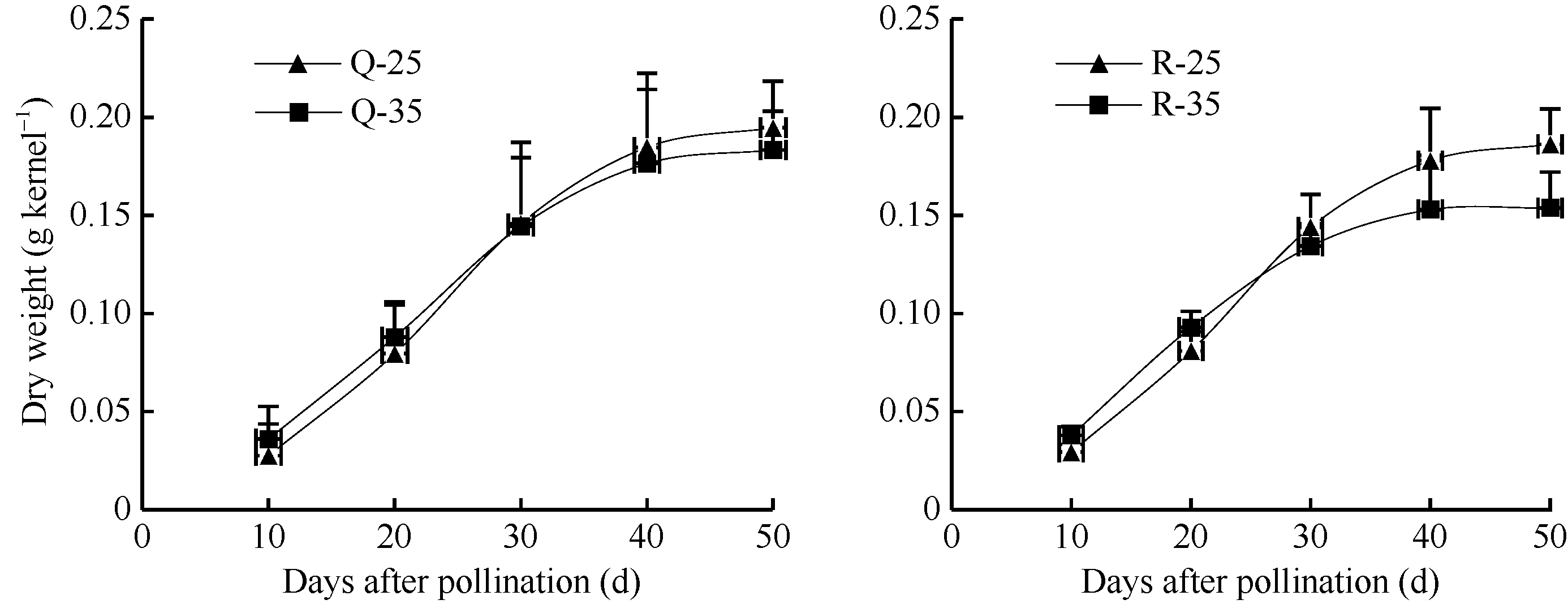 | 图1 花后高温处理下籽粒干重的变化Q-25: 强势粒常温; Q-35: 强势粒高温胁迫; R-25: 弱势粒常温; R-35: 弱势粒高温胁迫。Fig. 1 Changes of kernel dry weight under high temperature after floweringQ-25: superior kernels under normal temperature; Q-35: superior kernels under high temperature stress; R-25: inferior kernels under normal temperature; R-35: inferior kernels under high temperature stress. |
由图2可以看出, 常温处理下, 强、弱势粒的灌浆速率有相似的变化规律, 即随着培养时间的增加而加快, 在30 DAP时达到最大, 随后呈下降趋势, 在50 DAP降到最低。高温处理加快了强、弱势粒10~20 DAP灌浆速率, 与籽粒干重变化同步(图1), 但降低了籽粒灌浆中后期的速率。Q-35的灌浆速率在30 DAP达到最大, 比对照的最大灌浆速率降低14.9%。高温处理弱势粒的最大灌浆速率比对照低12.9%, 在20 DAP达到灌浆峰值, 比对照提前10 d, 可见, 高温不但降低了弱势粒灌浆中后期的速率, 而且缩短了灌浆持续时间, 可能是导致干重下降较严重的原因。
 | 图2 花后高温处理下籽粒灌浆速率的变化Q-25: 强势粒常温; Q-35: 强势粒高温胁迫; R-25: 弱势粒常温; R-35: 弱势粒高温胁迫。Fig. 2 Changes of grain-filling rate per kernel under high temperature after floweringQ-25: superior kernels under normal temperature; Q-35: superior kernels under high temperature stress; R-25: inferior kernels under normal temperature; R-35: inferior kernels under high temperature stress. |
由图3可知, 强弱势籽粒在高温下淀粉含量变化趋势相似。在10 DAP, Q-35和R-35的淀粉含量均高于对照, 与高温处理籽粒的干重变化一致, 但R-35的淀粉含量与对照差异显著。在20 DAP后, 高温处理的淀粉含量均低于对照, 差异均不显著。在40 DAP, 高温处理的弱势粒淀粉含量比对照减少6.3%, 强势粒少3.1%, 弱势粒淀粉含量降低程度大。
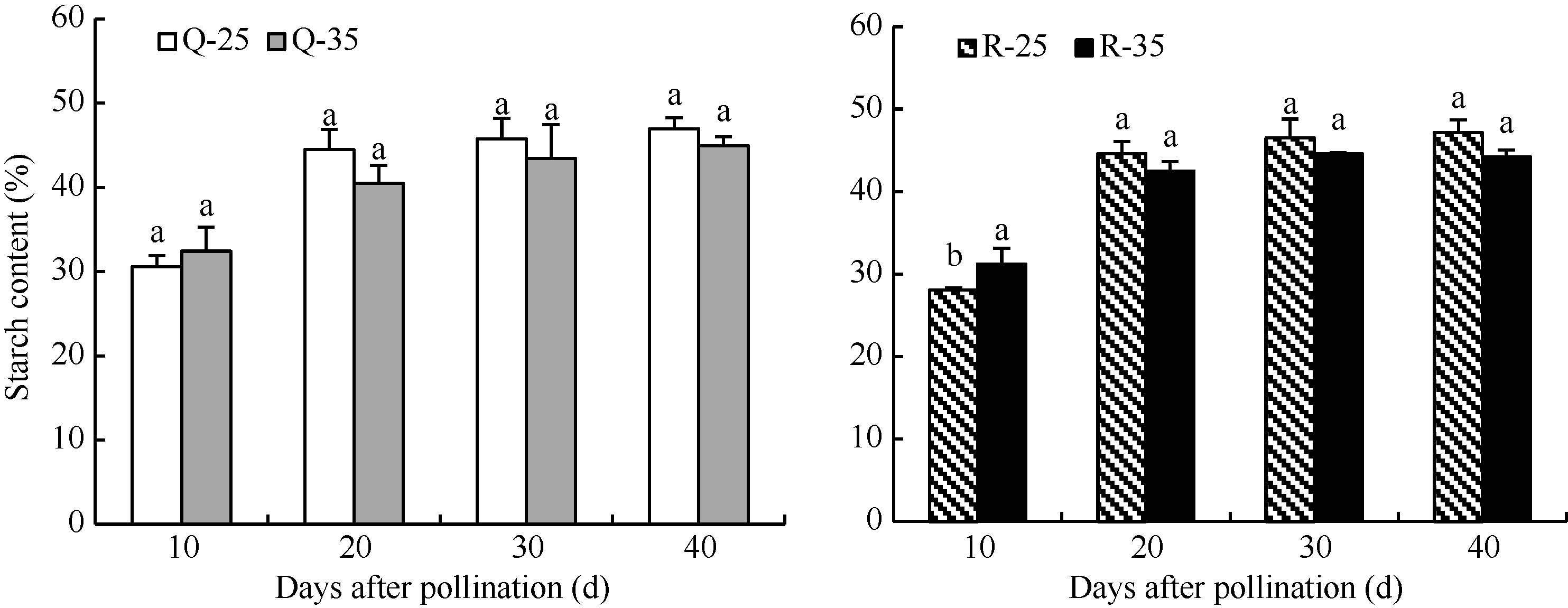 | 图3 花后高温处理下籽粒淀粉含量的变化Q-25: 强势粒常温; Q-35: 强势粒高温胁迫; R-25: 弱势粒常温; R-35: 弱势粒高温胁迫; 标以不同小写字母柱值在0.05水平上差异显著。Fig. 3 Changes of starch content under high temperature after floweringQ-25: superior kernels under normal temperature; Q-35: superior kernels under high temperature; R-25: inferior kernels under normal temperature; R-35: inferior kernels under high temperature; Bars superscripted by different lowercases are significant difference at 0.05 probability level. |
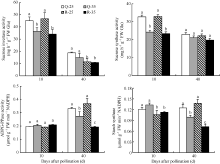
酸性蔗糖转化酶活性与同化物卸出及蔗糖水解密切相关, 而蔗糖合酶能催化降解细胞质中的蔗糖产生果糖和UDPG, 为淀粉合成提供底物, 因此二者与淀粉的合成密不可分。两种酶活性通常在籽粒灌浆早期相对较高, 在灌浆后期下降。从图4可以看出, 高温显著降低了籽粒灌浆前期的酸性蔗糖转化酶和蔗糖成酶的活性, 表明高温影响了籽粒灌浆前期淀粉合成过程中底物的供应。在10 DAP, Q-35的蔗糖转化酶、蔗糖合酶活性分别比对照降低19.1%、25.9%, 而R-35分别比对照降低27.1%、30.4%, 弱势粒的两种酶活性降低幅度均高于强势粒。在灌浆前期, 常温处理的弱、势籽粒的蔗糖转化酶和蔗糖合酶活性差异较小, 但在40 DAP, 强势粒处理的蔗糖转化酶活性显著高于弱势粒, 蔗糖合酶活性没有显著差异。
 | 图4 花后高温处理下籽粒淀粉合成相关酶活性的变化Q-25: 强势粒常温; Q-35: 强势粒高温胁迫; R-25: 弱势粒常温; R-35: 弱势粒高温胁迫; 标以不同小写字母柱值在0.05水平上差异显著。Fig. 4 Changes of activities of enzymes related to starch synthesis in kernel under high temperature after floweringQ-25: superior kernels under normal temperature; Q-35: superior kernels under high temperature; R-25: inferior kernels under normal temperature; R-35: inferior kernels under high temperature; Bars superscripted by different lowercases are significant difference at 0.05 probability level. |
淀粉合酶(SSS)和ADPG焦磷酸化酶(APGase)直接影响淀粉的合成, 在籽粒的灌浆前期活性较低, 灌浆后期活性较高。由图4可见, 在灌浆前期, 高温对强、弱势粒的SSS和APGase活性影响较小, 但灌浆后期两种酶的活性显著降低, 这可能是导致高温处理籽粒灌浆后期淀粉含量降低的原因。在40 DAP, Q-35和R-35的APGase活性分别比对照降低17.5%、47.7%, SSS活性分别降低20.8%、47.4%, 弱势粒的酶活性降低幅度大于强势粒, 且在10 DAP, 弱势粒处理的SSS活性显著小于强势粒。
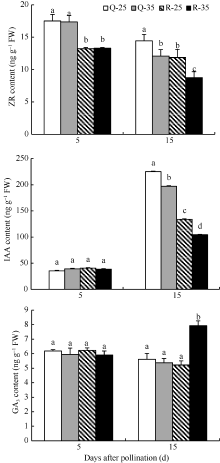
如图5所示, 高温处理前, 籽粒中的激素含量处于同一水平, 高温处理在15 DAP结束。在5 DAP, 强势粒中的ZR含量显著高于弱势粒, 可能是强势粒较弱势粒发育提前引起的。在15 DAP高温显著降低了籽粒的ZR、IAA含量, 强、弱势粒中的ZR含量分别降低了16.3%、26.2%, IAA含量分别降低了12.4%、22.1%, 弱势粒的降低幅度大, 且弱势粒处理的两种激素含量均显著低于强势粒。由此可见, 弱势粒中的部分激素含量不但低于强势粒, 而且高温敏感性较强。强、弱势粒的GA3含量变化不同, 高温对强势粒中的GA3含量影响较小, 没有显著差异, 却显著提高了弱势粒中GA3的含量, 这表明, 高温对弱势粒影响严重可能与GA3含量变化有关。
 | 图5 花后高温处理下籽粒中激素含量的变化Q-25: 强势粒常温; Q-35: 强势粒高温胁迫; R-25: 弱势粒常温; R-35: 弱势粒高温胁迫; 标以不同小写字母柱值在0.05水平上差异显著。Fig. 5 Changes of kernel hormone content under high temperature after floweringQ-25: superior kernels under normal temperature; Q-35: superior kernels under high temperature; R-25: inferior kernels under normal temperature; R-35: inferior kernels under high temperature; Bars superscripted by different lowercases are significant difference at 0.05 probability level. |
在玉米整个生育期间, 花期对高温最敏感, 研究多集中在开花后的高温。花后高温使玉米籽粒灌浆速率加快, 但灌浆时间缩短, 灌浆速率加快不能弥补灌浆持续期缩短对产量的负效应, 导致最终产量降低[ 15, 16, 17]。本研究发现, 花后连续10 d的高温处理(5~15 DAP)对籽粒灌浆速率影响较大, 籽粒发育前期(0~20 DAP)灌浆速率有一定的增加, 而影响了中后期的灌浆速率明显下降, 导致籽粒最终干重降低。产生这一结果的主要原因是高温在短时间内提高了籽粒胚乳细胞分裂速率[ 18], 促进了籽粒发育前期的生长, 从而引起前期灌浆速率和籽粒干重的增加; 但由于籽粒发育中后期以淀粉积累为主[ 19], 高温影响了籽粒淀粉合成相关酶的活性及激素含量, 使得淀粉合成受阻, 导致籽粒干重下降。有关不同部位玉米籽粒发育研究认为, 穗上部籽粒库容小, 导致干重积累少, 在本研究中发现, 强、弱势籽粒在充足的营养供应下有相似的干重积累, 与Hanft[ 20]的结果相同。但在高温条件下, 弱势粒受影响程度大于强势粒, 这与曹云英[ 21]在水稻上的研究结果相似。
淀粉是玉米籽粒的主要组成成分, 有研究认为, 高温降低了籽粒中淀粉含量, 不是因为光合产物供应不足, 而是影响了淀粉合成过程中的酶活性[ 22, 23]。Wilhelm等[ 24]发现授粉后15 d至成熟期高温处理对籽粒中大部分酶影响很小, 但显著降低了ADPG焦磷酸化酶、葡萄糖激酶、蔗糖合酶和淀粉合酶活性, Keeling等[ 25]则认为可溶性淀粉合酶、ADPG焦磷酸化酶和淀粉分枝酶对高温均呈现高度敏感性。可见, 淀粉合酶和ADPG焦磷酸化酶在玉米抗高温响应中有重要作用, 在本试验中, 花后前期高温影响了籽粒灌浆后期的ADPG焦磷酸化酶和淀粉合酶, 使其活性显著降低, 影响籽粒淀粉合成。此外, 试验中花后前期高温处理显著降低了籽粒灌浆前期的蔗糖转化酶和蔗糖合酶可能影响了籽粒淀粉合成过程中底物的供应, 导致籽粒最终淀粉含量降低, 是造成籽粒发育中后期灌浆速率降低并最终引起籽粒干重下降的重要原因。强、弱势粒相比, 弱势粒本身含有较低的酶活性(酸性蔗糖转化酶、淀粉合酶), 且高温条件下弱势粒的4种酶活性下降程度明显高于强势粒, 导致弱势粒的淀粉含量减少大于强势粒。
大量研究表明, 胁迫条件影响作物籽粒中植物激素的含量, 进而对籽粒的生长发育产生不利影响[ 26, 27]。据王丰等[ 28]研究, 在水稻灌浆初期高温处理后, 籽粒中的IAA、ZR和GA3含量明显低于适温处理, Cheikh等[ 29]认为, 高温不是影响籽粒中单一激素的含量, 而是改变籽粒中ABA与CTK含量的比值, 从而影响籽粒的发育。本试验结果显示, 花后连续10 d (5~15 DAP)的高温显著降低了强、弱势粒中的ZR和IAA含量, 弱势粒的下降程度较大。IAA含量与籽粒淀粉含量有很好的一致性[ 30], 在15 DAP强、弱势粒IAA含量的显著降低, 可能是导致籽粒在20 DAP淀粉含量下降的原因。ZR促进籽粒中胚乳细胞的分裂, 在高温处理前, 强势粒的ZR含量显著高于弱势粒, 与籽粒取样穗部不同有关。徐云姬等[ 31]研究发现, 玉米胚乳细胞活跃增殖和籽粒活跃灌浆期与籽粒中的GA3含量呈显著负相关。本试验中高温处理后弱势粒的GA3含量显著增加, 但在强势粒中却无明显变化。由此推测, GA3含量的增加进一步影响了弱势粒的胚乳细胞增殖和灌浆过程, 从而导致籽粒产量损失加大。
花后前期高温处理加快了籽粒发育前期的灌浆速率, 且灌浆中后期速率受到影响, 导致籽粒干物质积累减小。花后前期高温处理影响淀粉合成中底物的供应及淀粉的合成过程, 导致籽粒淀粉含量的下降。高温处理后籽粒中激素的含量变化也是导致籽粒干重减小的原因。强、弱势籽粒在高温处理后有相似的生理变化响应, 但高温对弱势粒的影响大于强势粒。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|