


{kind=link}
{kind=link}
{kind=link}
半干旱地区菊芋品系植株表型与光合特性分析
[吕世奇, 寇一翾, 杨彬, 曾军, 赵长明*  ]
]
]
|
|


为了加快半干旱地区的新型资源作物菊芋品种的选育进程, 有必要探讨菊芋产量及相关性状表型和光合生理基础。选用本课题组收集和培育的4个品系( 2个高产, 2个低产品系), 设置随机区组试验, 比较菊芋高产和低产品系与产量形成相关的生物量累积、植株表型和光合特性的差异。结果表明, 高产菊芋品系的地上生物量、块茎产量及根生物量显著高于低产品系; 高产品系整体表现出明显的生长优势, 其株高、基径、叶面积和叶片数均显著高于低产品系。气体交换参数分析表明, 高产品系的光补偿点和暗呼吸速率显著小于低产品系, 而其他各项参数差异不显著。相关性分析表明, 菊芋块茎产量与株高、基径、叶面积、叶片数、地上生物量和根生物量呈显著正相关, 而与光补偿点和暗呼吸速率呈显著负相关。建议多的叶片数、大的叶面积和发达的根系作为半干旱地区菊芋高产品系田间选育指标, 而低的光补偿点和暗呼吸速率在高产品系选育中也具参考价值。
To speed up breeding process for Jerusalem artichoke in semi-arid area, it is necessary to investigate traits related to yield and photosynthetic characteristics. Four stable lines including two high-yield lines and two low-yield lines were screened as materials, agronomic traits and photosynthesis parameters were determined. The results showed that the tuber yield, shoot biomass and root biomass of Jerusalem artichoke high-yield lines were significantly higher than those of low-yield lines, and the plant height, stem diameter, leaf area, leaf number of high-yield lines were also significantly higher than those of low-yield lines, then compared with low-yield lines, the high-yield lines showed a stronger growth vigor. LCP and
菊芋( Helianthus tuberosus L.)是菊科向日葵属的一年生草本植物, 又名洋姜、鬼子姜等, 起源于北美, 后经欧洲传入我国。菊芋具有较高的生态和经济价值, 是一种极具发展潜力的新兴作物, 现已被广泛栽培。菊芋抗寒抗旱耐盐碱耐贫瘠, 在温度最低大于-40℃, 年降雨量大于150 mm, 土壤盐含量小于150 mmol L-1的情况下均可生长, 菊芋块茎产量高, 正常情况下每公顷鲜块茎产量可达45~90 t[ 1, 2, 3, 4, 5]。菊芋块茎富含菊糖, 占其干重的70%~90%, 菊糖是一种果聚糖平均聚合度在10 DP左右, 与其他常见的碳水化合物不同, 菊糖不能被人体降解, 可降低能量摄入, 起到膳食纤维的作用, 已在医药和食品领域得到广泛应用, 块茎通过深加工可开发出低聚果糖、高聚果糖浆等具有较高附加值的生物基产品, 菊糖经水解后, 为发酵乙醇的良好原料[ 6, 7, 8]。
菊芋作为生产乙醇和生物柴油的重要生物原料, 具有更为广阔的发展前景。由于我国人多地少的现状, 国家在发展非粮能源植物时提出了“不与人争粮, 不与粮争地”的原则, 因此, 在保障国家粮食安全的前提下, 发展非粮生物质必须利用我国大量边际土地。菊芋适于边际土地种植、生态适应性强、生物量大, 不涉及粮食安全问题、管理成本低、易转化, 是一种理想的非粮能源植物, 我国西北半干旱地区有大量边际土地适宜菊芋生长, 种植菊芋不仅可获得巨大的经济效益而且具有良好的生态效益[ 9, 10, 11, 12, 13]。目前国内外菊芋的研究主要集中在抗旱、抗盐、抗重金属的生理生态特性与优良品种传统选育及田间栽培技术[ 14, 15, 16, 17], 而对菊芋高产品系产量形成的表型和光合生理基础鲜见报道, 尤其在高产品系选育过程中缺少可靠的选育标准, 使菊芋在高产品种选育方面滞后于甜高粱等其他能源作物, 从而限制了菊芋作为能源植物的大面积规模化推广种植。因此, 本文在西北半干旱地区通过对菊芋高产品系植株表型性状与光合生理特性分析, 以期探讨菊芋产量形成的表型和光合生理基础, 明确适于菊芋高产品系选育的直观可靠标准, 为生产实践中菊芋高产品系的选育提供理论和技术支撑。
兰州大学榆中校区植物种质资源圃(35°56′N, 104°09′E), 年均温7.1℃, 年均降雨量300 mm, 年均蒸发量1450 mm, 无霜期130 d, 年均日照时数2666 h, 土壤类型为黒垆土, 耕层含有机质12.6 g kg-1、全氮1.05 g kg-1、全磷0.79 g kg-1、全钾8.32 g kg-1、速效氮91.9 mg kg-1、速效磷23 mg kg-1、速效钾168.5 mg kg-1, pH 8.14、容重1.25 g cm-2 [ 18]。
2011—2012年种植本课题组收集和选育的稳定高产菊芋品系(LZJ119和LZJ004)和低产菊芋品系(LZJ188和LZJ018)。按完全随机区组设计, 将平整好的地块分为12个小区, 每个品系3个重复, 每个小区面积45 m2, 播种株行距为40 cm×40 cm。两年均在3月20日选取具有1~2个芽眼的30 g左右的块茎播种, 播深为10~15 cm; 苗期适时除草, 且在形态建成期和块茎膨大期各浇水一次, 每次浇水量为300 m3 hm-2。
7月下旬晴天9:00至11:30, 选取菊芋顶部第5片叶子利用便携式光合作用测定系统(Li-6400, Li-COR USA)测定光响应曲线和CO2响应曲线。测定时选取3~5片叶子重复, 设定叶室温度30℃和流速500 µmol s-1。测定光响应曲线时设定光强为1600、1200、1000、800、600、400、200、150、100、80、50、25和0 µmol m-2 s-1外加CO2钢瓶设定参比室CO2浓度为380 µmol mol-1; 测定CO2响应曲线时光强设置为1500 µmol m-2 s-1, CO2浓度依次设定为400、200、150、100、80、50、200、400、600、800、1000、1200、1600和2000 µmol mol-1。通过非直角双曲线模型[ 19, 20]拟合光响应曲线和CO2响应曲线并且结合光强小于200 µmol m-2 s-1以下线性拟合和CO2浓度小于200 µmol mol-1以下线性拟合, 得到光饱和下的最大光合速率( Pmax)、CO2饱和下的最大光合速率( Amax)、光饱和点(LSP)和CO2饱和点(CSP); 暗呼吸速率( Rd)、表观量子效率(AQY)和光补偿点(LCP), 光呼吸速率( Rp)、羧化效率(CE)和CO2补偿点(CCP)。
9月初, 菊芋开花后人工用卷尺测定株高和节长, 用游标卡尺测定基径, 用Li-3000a (Li-COR USA)叶面积仪测定整株叶面积, 统计叶片数和分枝数。11月上旬, 待菊芋地上部分完全干枯后以单株为单位收获, 将地上部分和根部分于烘箱中85℃烘至恒重, 称其生物量干重, 而地下块茎部分收获时按其鲜重计算。
采用Microsoft Excel 2007和SPSS16.0分析处理数据, 应用LSD法分析差异显著性, 用Origin 8.0作图, 2年试验结果趋势一致, 本文主要分析2012年数据。
从表1可见2年试验的菊芋高产品系与低产品系在块茎产量、地上生物量和根生物量上表现一致且均有显著的差异( P<0.05)。高产品系2011年和2012年的单株块茎产量、地上生物量和根生物量分别比低产品系高36.05%、27.37%、48.57%和39.86%、38.85%、59.72%; 根生物量的增幅最大, 块茎产量次之而地上生物量最小, 可见发达的根系是菊芋块茎高产的关键指标, 其利于菊芋水分和养分吸收从而促进地上器官生长和增加光合产物积累。
| 表1 不同菊芋品系块茎产量、根和地上生物量特征 Table 1 Comparison of tuber yield, root weight and shoot weight in Jerusalem artichoke lines |
表2表明, 高产品系的株高、基径、分枝数、叶片数、叶面积以及节长均大于低产品系且差异显著。其中高产品系分枝数差异达到3.97倍, 株高的差异达到1.29倍, 多的分枝以及高的株高可以着生多的叶片极大地增加叶面积, 使其叶面积的差异达到了1.60倍, 叶片数的差异也达到1.23倍, 而且基径和节长的差异也分别达到了1.22倍和1.05倍。可见, 高产品系的植株表型性状值较低产品系均有显著优势, 呈现较强的生长势。
| 表2 菊芋品系的叶面积、叶片数、株高、基径、分枝数和节长特征 Table 2 Leaf area, leaf number, plant height, stem diameter, branch number, and internode length in Jerusalem artichoke lines |
由图1各菊芋品系净光合速率随光照强度和胞间CO2浓度变化响应及相关拟合参数(表3和表4)可见, 在CO2饱和浓度下 Amax无论菊芋高产品系还是低产品系都是光饱和下的 Pmax的2倍多, 可见在强光下菊芋光合速率很大程度上受到CO2供应的限制。高产品系的LCP和 Rd均显著小于低产品系, 其中高产品系的LCP和 Rd分别比低产品系低74.12%和66.44%, 其余各项参数差异并不显著, 但高产品系的AQY依然显示出增高的趋势和LSP降低的趋势。虽然菊芋高产品系的CE、CCP、CSP、 Amax和 Rp略低于低产品系(变异幅度均小于7%), 但各项参数间差异并不显著(表4)。表明对于菊芋高产品系较低的LCP和 Rd对于其提高光合产物的积累是十分有利的。
| 表3 菊芋各品系光响应曲线拟合的特征参数 Table 3 Fitting parameters of light response curve in Jerusalem artichoke lines |
| 表4 菊芋各品系CO2响应曲线拟合的特征参数 Table 4 Fitting parameters of CO2 response curve in Jerusalem artichoke lines |
 | 图1 菊芋各品系光合速率( Pn)随光照强度(PAR)和胞间CO2浓度( Ci)的变化黑色三角代表高产品系, 白色三角代表低产品系。Fig.1 Changes of net photosynthetic rate with different photosynthetically active radiation and intercellular CO2 concentrations in Jerusalem artichoke linesBlack triangles represent high yield lines, white triangles represent low yield lines. Pn: photosynthetic rate; PAR: photosynthetically active radiation; Ci: intercellular CO2 concentration. |
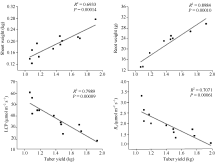
由图2可知, 菊芋基径、株高、叶片数和叶面积与块茎产量均呈显著的正相关, 且叶片数和叶面积与块茎产量拟合系数( R2=0.87和0.80)高于株高和基径拟合系数( R2=0.69和0.64)。地上生物量和根生物量与块茎产量的拟合发现(图3), 二者与块茎产量间也呈显著的正相关关系且根生物量拟合系数( R2)高达0.90, 而LCP和 Rd与块茎产量间呈显著负相关( P<0.01)。可见, 对于菊芋而言较大的叶片数和叶面积以及发达的根系和较低的LCP和 Rd对于其块茎产量的提高起重要作用。
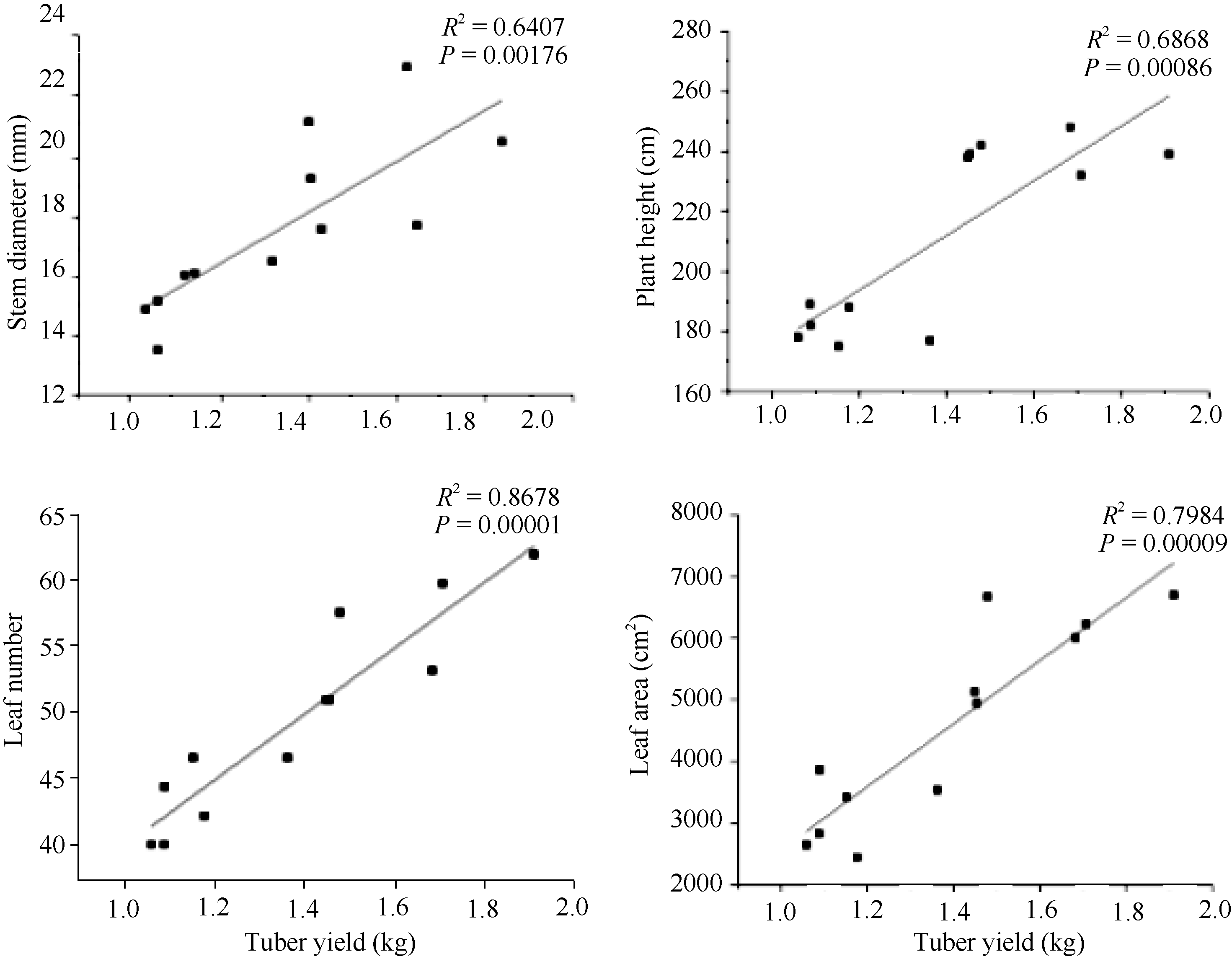 | 图2 块茎产量与基径、株高、叶片数和叶面积的相互关系Fig. 2 Relationship of tuber yield with stem diameter, plant height, leaf number, and leaf area |
作物产量是由物质生产总量和收获指数共同决定的, 目前学者们一致认为对于同一种作物当其收获指数达到较高水平时, 提高其生物量是获得高产的主要途径[ 21, 22], 且作物的表型性状在其产量的形成过程中具有重要的作用, 其中叶面积、叶片数、株高等与作物的光合有机积累关系比较密切表型特征对作物的产量影响较为显著, 并且有研究已表明在马铃薯中叶面积与产量间呈显著正相关, 小麦中顶三叶面积与生物量和经济产量呈显著正相关, 玉米中灌浆期的叶面积系数对产量的影响最大, 旱稻中剑叶长与产量的关系最为密切, 苜蓿中全年产草量与其第2次刈割时的生长高度之间的相关程度较高[ 23, 24, 25, 26, 27]。菊芋也不例外, 在块茎迅速膨大期之前光合产物暂时储存在茎杆中, 至后来的块茎快速膨大期运输到块茎中, 从而促进块茎膨大和菊糖的累积。高产品系在开花前的形态建成期旺盛的生长, 拥有较大的株高和基径, 在增加菊芋对菊糖暂时储存能力的同时还可以增强其抗倒伏能力, 较大的叶面积和叶片数可以增加其对光的截获能力, 增大光合叶面积有利于增加光合产物的生产, 对于植株产量的形成是十分重要的, 与前人研究结果一致[ 28, 29], 菊芋块茎产量和其株高、基径、叶面积和叶片数呈显著正相关(图2)。此时, 高产品系地上部分的生长表现较强的生长势且具有较大的根系增强对水分和养分的吸收以促进植株光合产物的积累[ 30, 31, 32, 33]。我们的研究结果也充分印证了这一点, 高产品系与低产品系相比, 营养器官各部分增幅中根生物量的增幅最大, 可达到59.72% (表1), 且根生物量与块茎产量间的相关系数最高(图3)。因此, 对于菊芋而言较多叶片数和较大叶面积以及发达的根系是其高产的关键。
通常植物干物质的90%以上来自光合作用, 光合作用是作物产量形成的基础, 这就决定了作物的光合性能与其产量间一定是正相互关系。由于作物的产量受到光合生产能力、呼吸消耗、收获指数等众多因素的影响, 其中光合生产能力的大小又受光合速率高低、光合叶面积大小和光合功能期的长短的影响, 往往使光合性能与产量间的这种正相互关系被掩盖, 对在同一生境下生长的同一作物, 其光合特性的差异对于其产量差异的分析还是有重要的参考价值[ 34]。光合作用是一个极其复杂的生理过程, 受到各种因素的影响尤其是光和CO2浓度, 光是植物光合作用最终的能量来源, 而CO2是植物光合作用中碳的来源, 二者是影响光合作用的最重要因素, 作物在全生育期通过捕获和利用太阳能同化CO2以达到最大限度提高产量的目的[ 35, 36]。因此, 分析作物叶片光合速率对光和CO2响应过程是研究作物光合生理特性的重要手段之一。从表3和表4菊芋叶片光响应曲线和CO2响应曲线的拟合结果可见, 虽然菊芋高产品系叶片LSP、AQY、CCP、CSP、CE和 Rp等与低产品系差异不显著, 但高产品系的LCP和 Rd显著小于低产品系, 且从图3中可见菊芋叶片LCP和 Rd与块茎产量间呈显著负相关, 可见较低的LCP极大地提高了菊芋叶片对弱光的利用能力, 使高产品系叶片在较低的光强下能够有效进行光合作用, 这对于菊芋在大田高密度种植时尤为重要, 此时中下部叶片易受到相互遮挡使得光照变弱, 高产品系仍能保持较高的光合能力; 而较低的 Rd可以极大地减少植物对光合产物的消耗。总之, 菊芋高产品系较低的LCP和 Rd不仅可以使其在弱光下光合产物生产增加还可减少光合产物消耗, 从而增加对光合产物的积累[ 37, 38]。
此外, 虽然除LCP和 Rd外, 菊芋高产品系与低产品系其他各项光合特征参数差异均不显著, 但是在植株表型上高产品系的整株叶面积显著高于低产品系(1.6倍), 极大提高了整株光合有效叶面积, 再加上低的LCP和 Rd, 更有利于光合有机物的积累, 从而促进了高产。由此可见, 菊芋低的LCP以增加其对弱光的利用能力和低的 Rd以降低其对光合产物的消耗以及较大的叶面积对其产量的提高是十分必要的, 这与董志新等[ 39]和张振文等[ 40]在苜蓿和葡萄上的研究结果相似。最后, 无论菊芋高产品系还是低产品系饱和CO2浓度下 Amax均大于光饱和下的 Pmax且差异达到2倍多(表3和表4), 说明在饱和光强下菊芋光合速率很大程度上受到CO2供应的限制而且在饱和光强下提高其周围CO2浓度短期内能够显著提高叶片光合速率, 因此, 可尝试利用喷施CO2叶面肥提高菊芋叶片光合产物的累积, 但对于菊芋能否长期适应高CO2浓度环境并保持高的光合速率有待进一步研究。
在菊芋高产品系的选育过程中较大的叶片数和叶面积以及发达的根系可作为比较直观可靠的选育标准, 而且较低LCP和 Rd也具有较高的参考价值。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|