 g cm-2 in untreated plants to 46.56
g cm-2 in untreated plants to 46.56 {kind=link}
燕麦对碱胁迫的阳离子响应机制
[萨如拉1  , 刘景辉
, 刘景辉1, * , 刘伟1 , 白健慧1 , 王占海2 ]
, 刘景辉, 刘伟|
|


以耐碱性燕麦品种Vao-9和碱敏感性品种白燕5号为试验材料, 采用盆栽法, 用25、50、75和100 mmol L-1碱浓度(Na2CO3和NaHCO3按摩尔比1∶1混合)进行短期(14 d)和长期(28 d)胁迫处理, 观测两品种根、茎、叶中Na+、K+、Ca2+、Mg2+吸收及分配特点, 并从离子平衡吸收与分配角度, 探讨燕麦对碱胁迫的生理适应机制。胁迫处理14 d后, 燕麦体内Na+增加, K+下降, Ca2+和Mg2+变化不大, 且两品种间各器官中4种离子的分配比例差异不显著。胁迫处理28 d后, 两品种各器官中Na+增幅较大, K+、Ca2+和Mg2+降幅较大。Vao-9植株体内Na+、Ca2+含量大于白燕5号, 但K+、Mg2+含量与白燕5号无显著差异, 但两品种间各器官中4种离子的分配特点不同; 当胁迫浓度达到100 mmol L-1时, 与白燕5号相比, Vao-9叶片中少分配5.9个百分点Na+, 多分配13.5个百分点K+、28.9个百分点Ca2+、10.9个百分点Mg2+, 茎中多分配5.4个百分点Na+, 少分配9.8个百分点K+, 根中少分配28.9个百分点Ca2+、10.9个百分点Mg2+, 因而Vao-9叶片中Na+ /K+、Na+/Ca2+、Na+/Mg2+值较白燕5号低。可见, 燕麦通过提高阳离子选择吸收及器官分配能力以适应碱胁迫。
第一作者联系方式: E-mail:sarula_0922@126.com, Tel: 15848101483
In a pot experiment, an alkali-tolerant oat variety, Vao-9, and an alkali-sensitive variety, Baiyan 5, were exposed to 25, 50, 75, and 100 mmol L-1 of alkali stress (molar ratio of Na2CO3:NaHCO3 = 1:1). The contents of Na+, K+, Ca2+, and Mg2+ absorbed by oat seedling and their distribution characteristics in root, stem, and leaf were measured after a short-term (14 d) or a long-term (28 d) stress treatment. The objective was to understand the physiological adaptation to alkali stress in oat in the view of ion balance absorption and distribution. After short-term stress, the cation contents in oat plants showed the variations of increased Na+, decreased K+, and minor changes in Ca2+ and Mg2+. Besides, the distribution proportions of the four ions in various organs were not significantly different between the two varieties. Compared to short-term stress, long-term stress resulted in larger increase of Na+ content and larger decreases of K+, Ca2+, and Mg2+ contents in all organs of both varieties. Vao-9 absorbed more Na+and Ca2+ than Baiyan 5, and the absorptions of K+ and Mg2+ were not significantly different between varieties, but the distribution characteristics of the four ions in various organs were different between the two varieties. Under 100 mmol L-1 alkali stress, 5.9 percent Na+ in leaf were lower distributed and 13.5 percent K+, 28.9 Ca2+, 10.9 Mg2+ more distributed in Vao-9 than in Baiyan 5, 5.4 percent Na+ in stem higher and 9.8 K+lower in Vao-9 than in Baiyan 5, 28.9 Ca2+, 10.9 Mg2+ in root were lower in Vao-9 than in Baiyan 5, As a result, the Na+ content and the ratios of Na+/K+, Na+/Ca2+, and Na+/Mg2+ in leaf were lower in Vao-9 than in Baiyan 5. Clearly, oat plant has the mechanism of selective absorption and distribution of various cations in different organs in response to alkali stress.
盐渍化土壤严重影响作物生长和产量[ 1]。Na+是造成植物盐害及产生盐渍生境的主要离子。由于Na+和K+有相似的离子半径和水合能[ 2], 高浓度Na+竞争K+, 使植物体内K+高亲和力AKT1等离子通道及HKT1等转运蛋白结合位的K+吸收及转运受到抑制[ 3]。高浓度Na+可置换质膜和细胞内膜上结合的Ca2+, 破坏膜系统完整性, 增加膜透性, 扰乱离子的选择运输能力。在盐碱胁迫下, 由于阳离子竞争效应, 植物对Mg2+的吸收和向地上部运输受Na+、K+、Ca2+抑制[ 4, 5], 而这4种离子的平衡吸收选择与分配能力与其耐盐碱性有关[ 6]。
燕麦( Arena sativa)作为粮饲兼用的一年生草本植物, 具有抗寒、耐贫瘠和耐盐碱等特性, 被称为盐碱地改良的先锋作物[ 7]。研究表明, 燕麦在盐碱胁迫下合成和积累有机溶质(脯氨酸和可溶性糖)以及吸收和积累无机离子(Na+)来进行渗透调节, 以适应盐碱胁迫, 且不同时期、不同燕麦品种间存在差异[ 7, 8, 9]。盐碱胁迫下, 植物对Na+、K+、Ca2+、Mg2+吸收选择及分配能力不仅与胁迫时间和胁迫程度有关, 且与品种类型有关[ 10, 11, 12, 13]。目前, 不同燕麦品种不同器官中Ca2+、Mg2+对燕麦耐碱性影响的研究报道较少。本试验选用耐碱性不同的2个燕麦品种, 通过不同浓度碱溶液处理, 观测短期和长期胁迫后不同器官中Na+、K+、Ca2+、Mg2+平衡吸收与分配规律差异, 为揭示燕麦对碱胁迫的阳离子响应机制提供依据。
耐碱性燕麦品种Vao-9由加拿大东部谷物与油料作物研究中心提供, 碱敏感性品种白燕5号由吉林省白城市农业科学院提供。
2012年4月至7月, 在内蒙古农业大学燕麦产业研究中心日光温室内进行盆栽试验。将种子播于直径22 cm、高30 cm的塑料盆内, 每盆装7.5 kg细沙, 定苗20株。每隔2 d浇一次Hogland营养液, 每次500 mL。待燕麦长至二叶一心时进行短期(14 d)和长期(28 d)胁迫处理。
按摩尔比1∶1混合2种碱性盐(Na2CO3、NaHCO3), 配成25、50、75和100 mmol L-1浓度碱液, 分别加入Hogland营养液(共500 mL), 作为胁迫处理液; 对照只加Hogland营养液500 mL。3次重复。为避免盐激反应, 各处理每天递增预定浓度的1/4, 每隔2 d加一次碱溶液, 使各处理同一天达到预定浓度, 此时作为胁迫第1天, 分别于处理后第14天和第28天取样。
取出每盆中所有植株, 用自来水冲洗干净, 再用蒸馏水冲洗, 并用吸水纸吸去表面附着的水分。将根、茎、叶分开, 105℃杀青后80℃烘干至恒量。样品磨碎后准确称取干样0.3000 g, 放入50 mL的塑料管中, 加1 mol L-1盐酸25.0 mL, 盖紧管口, 振荡2 h后用定量滤纸过滤。用火焰光度计测定K+(直接用滤液)和Na+(滤液稀释5倍后)含量。
取0.5000 g干样放入150 mL三角瓶, 加HNO3-HClO4 (4∶1)溶液20 mL, 放通风厨里在电热板上加热至140~150℃, 消煮过夜, 至冒白烟为止。把消煮液倒在 25 mL的比色管里定容, 在原子吸收光谱仪上测Ca2+和Mg2+含量。
采用Microsoft Excel绘制柱形图和SPSS 17. 0软件进行试验数据的方差分析(one-way ANOVA)。
两品种的Na+含量均随碱浓度的升高而增加, 且处理间有极显著差异( P<0.01)。短期胁迫后, Vao-9 Na+总量大于白燕5号, 但Na+在各器官的分布品种间无显著差异(图1-A1)。长期胁迫后, 两品种Na+总量差异不大, 分别为67.7 mg g-1 DW和65.6 mg g-1 DW, 但Na+在不同器官中的分布品种间差异明显, Vao-9主要在茎中积累, 而白燕5号在叶片中有大量积累(图1-B1)。在100 mmol L-1长期胁迫下, Vao-9叶片中Na+分配百分比较白燕5号低5.9个百分点, 而根和茎中的分配百分比分别较白燕5号高0.5和5.4个百分点(图1-C1)。
K+主要分布在茎和叶中, 根中含量很低。随碱胁迫浓度增加和时间的推移, K+含量在各器官中均呈下降趋势, 且两品种表现一致(图1-A2, B2)。各器官中不同碱浓度处理的K+含量有极显著差异( P<0.01)。在短期胁迫下, Vao-9吸收的K+总量小于白燕5号, 但两品种不同器官中分布差异不显著(图1-A2); 长期胁迫下, Vao-9的K+总量为118.1 mg g-1 DW, 白燕5号为118.2 mg g-1 DW, 但K+在茎、叶中的分布两品种间不同, 尤其是75 mmol L-1和100 mmol L-1处理(图1-B2), 其中100 mmol L-1处理叶片中K+分配百分比, Vao-9较白燕5号高13.5个百分点, 而茎中则少9.8个百分点(图1-C2)。
短期胁迫下, 不同处理Vao-9各器官的Ca2+含量始终大于白燕5号, 且处理间变化幅度较小(图1-A3)。与短期胁迫相比, 长期胁迫使两品种根和叶中的Ca2+骤增, 而茎中减少。在长期胁迫下, 随碱浓度增加, 两品种茎、叶中Ca2+含量降低; 而根中Ca2+含量, Vao-9中先降低再升高, 白燕5号表现持续增加, 且各处理间差异显著(图1-B3)。Vao-9的Ca2+总量(36.5 mg g-1 DW)大于白燕5号(29.6 mg g-1 DW)。4种浓度处理的叶片Ca2+含量均低于对照, 但Vao-9的下降百分率(10.3%~22.2%)明显低于白燕5号(42.8%~79.6%)。100 mmol L-1长期胁迫下, Ca2+在茎、叶片中分配比例, Vao-9分别比白燕5号高1.5和28.9个百分点, 而根中比例则低30.4个百分点(图1-C3)。
短期胁迫下, 除Vao-9叶片中Mg2+含量随碱浓度的增加表现先上升后下降外, 其他均呈逐渐降低趋势, 但Vao-9的降幅小于白燕5号(图1-A4)。长期胁迫下, 除白燕5号根中Mg2+含量升高外, 其他均呈降低趋势; 两品种的Mg2+总量无显著差异, 但Mg2+在不同器官中的分布特点不同。两品种叶片中Mg2+含量随碱浓度的升高而下降, Vao-9较对照下降0~20.6%, 低于白燕5号的11.7%~36.7% (图1-B4)。与Ca2+相似, 100 mmol L-1长期胁迫下, Vao-9的茎和叶中比白燕5号分配了较高比例的Mg2+, 而根中Mg2+分配比例则低于白燕5号(图1-C4)。
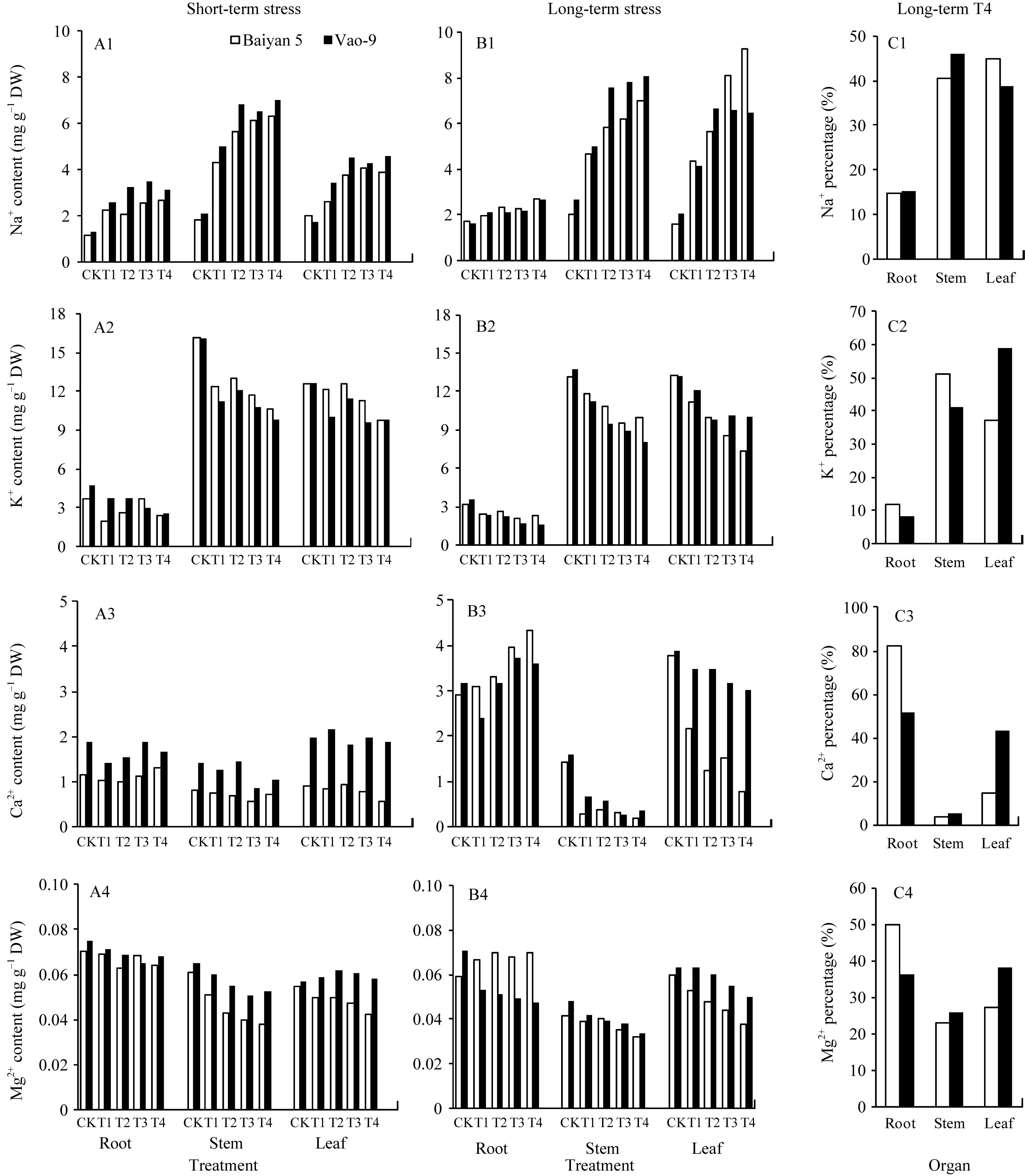 | 图1 不同浓度碱胁迫下燕麦根、茎、叶中4种阳离子含量与分配百分率短期(14 d)和长期(28 d)胁迫下, 各处理的碱浓度依次为0 (CK)、25 (T1)、50 (T2)、75 (T3)和100 mmol L-1 (T4)。Fig. 1 Contents and distribution ratios of four cations in root, stem, and leaf of oat under different concentrations of alkali stressAlkali concentrations in different treatments were 0 (CK), 25 (T1), 50 (T2), 75 (T3), and 100 mmol L-1 (T4) under both short-term (14 d) and long-term (28 d) stress. |
两品种的Na+/K+值随着胁迫浓度的增加而增加, 在根中最大, 这主要是根中K+含量低所致。短期胁迫下, Vao-9茎、叶Na+/K+值大于白燕5号, 根Na+/K+值在胁迫浓度达到50 mmol L-1时大于白燕5号; 长期胁迫下, Vao-9叶片Na+/K+值较白燕5号小, 且白燕5号各处理间均达极显著差异, 但根和茎的Na+/K+值较白燕5号大, 其中100 mmol L-1处理的根、茎、叶Na+/K+值, Vao-9分别为白燕5号的144.4%、140.8%和51.2% (表1)。
Na+/Ca2+值表现为茎>叶>根; 两品种茎和叶的Na+/Ca2+值呈现随着胁迫浓度增加而增加的趋势, 且白燕5号处理间存在极显著差异, 但短期胁迫下75 mmol L-1处理的茎Na+/Ca2+值高于100 mmol L-1处理, Vao-9叶Na+/Ca2+值在浓度达50 mmol L-1时基本稳定。无论短期还是长期胁迫, 白燕5号的茎、叶Na+/Ca2+值均高于Vao-9, 长期胁迫下的差异更为明显, 尤其是长期胁迫的100 mmol L-1处理, Vao-9的茎、叶Na+/Ca2+值分别只有白燕5号的60.4%和17.7% (表1)。
| 表1 短期和长期碱胁迫对燕麦不同器官中Na+/K+、Na+/Ca2+和Na+/Mg2+值的影响 Table 1 Effects of short-term and long-term alkali stresses on Na+/K+, Na+/Ca2+, and Na+/Mg2+ ratios in different organs of oat |
Na+/Mg2+值也表现为茎>叶>根。随着胁迫浓度提高, Na+/Mg2+值在各器官中也呈增加趋势, 但短期胁迫下, Vao-9根Na+/Mg2+值大于白燕5号, 茎、叶Na+/Mg2+值小于白燕5号。长期胁迫下两品种的茎、叶Na+/Mg2+值均大于短期胁迫的Na+/Mg2+值。长期胁迫下, Vao-9的叶Na+/Mg2+值远小于白燕5号, 且处理间差异极显著, 而根、茎Na+/Mg2+值则表现为Vao-9大于白燕5号; 当100 mmol L-1长期胁迫时, Vao-9根、茎、叶中Na+/Mg2+值分别为白燕5号的145.6%、109.9%和53.1% (表1)。
渗透调节在作物耐盐碱机制中有重要作用[ 14, 15, 16]。参与渗透调节的阳离子主要是Na+、K+和Ca2+, 其在植物器官中的分配比例因物种、品种和生育时期而异[ 17, 18, 19]。通过调节阳离子在不同器官、组织或细胞内的区域(隔)化的分布, 有助于减轻离子毒害[ 20]。本研究中, 长期和短期胁迫后, 两品种的Na+含量均显著增加, 且向茎、叶中大量积累; 两种胁迫强度下, Vao-9的Na+总量总是大于白燕5号, 长期胁迫下, Na+在Vao-9茎中的积累及分配较高, 而在叶片中的积累相对较低。这一方面是因为Vao-9通过地上部积累大量离子, 增大地上部和根部的渗透势差, 促进水分从根部向地上部运输, 改善地上部的水分状况, 促进生长[ 21, 22]; 另一方面是通过把吸收的大部分Na+分配在茎部, 从而减少叶片受伤害[ 23, 24]。
盐碱土壤中高浓度Na+经常阻止高亲和力K+转运蛋白导致作物体内Na+内流增加、K+内流减少, 使植物细胞膜去极化而减少净K+的吸收, 因此盐渍环境中植物叶片中K+减少、Na+/K+值升高。在胁迫下植物细胞质保持高K+/Na+的能力是植物适应盐碱胁迫的决定因子之一。本试验中, 长期胁迫下, 两品种吸收的K+总量比短期胁迫的降幅, Vao-9小于白燕5号, 且长期胁迫下两品种不同器官中K+分配差异显著, Vao-9叶片中积累及分配较高K+含量, 尤其是75 mmol L-1处理下叶片中积累K+能力更强, 使叶片中保持较低Na+/K+值。在长期胁迫下, 虽然与Vao-9吸收的K+总量无差异, 但白燕5号积累的大部分K+分配在茎部, 使胁迫下叶片中Na+/K+值较高。这与赵旭等[ 25]在小麦耐盐性研究中的结果一致。
Ca2+不仅对于保持植物细胞膜的完整性和稳定性有重要作用, 而且与植物的耐盐碱能力有重要关联[ 26], 由于胁迫下高活性Na+置换质膜上的Ca2+, 细胞中Ca2+浓度降低, 导致质膜上Ca2+失去平衡, 失去保护细胞的作用, 使细胞膜的稳定性和选择性下降而破坏作物体内离子平衡。本研究中, 碱胁迫降低了两品种的Ca2+含量, 但随着胁迫时间的延长, Ca2+总量增加, 这与范远等[ 11]在白燕2号上试验结果不一致, 可能不同基因型品种间存在差异导致适应胁迫的机制有所不同。碱胁迫条件下, Vao-9积累的Ca2+总量大于白燕5号, 且长期胁迫各处理下Vao-9根、叶中积累及分配较多Ca2+, 而白燕5号叶片中分配的Ca2+远少于根中, 使胁迫下Vao-9可以保持叶片中较低的Na+/Ca2+值。这有利于降低叶片细胞膜透性, 保持叶片中离子平衡和细胞正常代谢。同时与长期胁迫相比, 短期胁迫的各处理中, 两品种Ca2+含量始终保持较低水平和较小变幅, 说明碱胁迫下燕麦吸收及分配的Ca2+除基因型差异外还受胁迫时间的影响。
Mg2+是叶绿素分子的重要组成部分, 而盐碱胁迫下作物Na+增加Ca2+降低进一步限制了K+吸收, Ca2+吸收的降低和Na+的大量积累进一步限制Mg2+吸收, 由于K+与Mg2+间存在拮抗作用, 且主要发生在由根系向地上部的运输过程中[ 4, 5], 所以在盐碱胁迫下作物叶片中保持较低Na+/Mg2+值是衡量植物耐盐碱的重要指标之一[ 27, 28]。本研究中, 碱胁迫降低燕麦Mg2+含量, 且随着胁迫时间的延长而减少, 但Vao-9中降幅小于白燕5号。长期胁迫下两品种各器官中分布差异显著, Vao-9在叶中积累较多Mg2+, 而白燕5号则在根中积累较多Mg2+, 因而Vao-9叶片中保持较低的Na+/Mg2+值。这证实了不同刺槐品种存在Mg2+吸收选择能力差异的结果[ 29]。在棉花、碱地肤上的研究表明, 在盐、碱胁迫下, 作物对Na+的区域化分配, K+、Ca2+、Mg2+的吸收, 与叶片细胞质膜H+-ATPase、液泡膜H+-ATPase、H+-PPase和Na+/H+逆向运输蛋白等活性有关[ 30, 31]。燕麦的耐碱机制是否与上述研究结果一致, 需进一步探讨。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|