

 -carotene,
-carotene,  -carotene desaturase, an enzyme catalyzing an early step in the carotenoid biosynthetic pathway. If the QTL identified in this study are confirmed, particularly those associated with candidates genes, they could be used in an efficient marker-assisted selection program to facilitate increasing levels of carotenoids in maize grain.
-carotene desaturase, an enzyme catalyzing an early step in the carotenoid biosynthetic pathway. If the QTL identified in this study are confirmed, particularly those associated with candidates genes, they could be used in an efficient marker-assisted selection program to facilitate increasing levels of carotenoids in maize grain.{kind=link}
{kind=link}
甜玉米乳熟期籽粒维生素A源和维生素E组分的变异
[冯发强1  , 王国华
, 王国华1 , 王青峰1 , 杨瑞春1 , 李小琴1, 2, * ]
, 王国华]
|
|


第一作者联系方式: E-mail:fengfq@scau.edu.cn, Tel: 020-85285430
维生素A和维生素E是人与动物必须从食物中摄取的微量营养物质。甜玉米是人类直接食用的一种鲜食玉米类型。本研究利用高效液相色谱法测定了47份甜玉米自交系籽粒乳熟期的维生素A源和维生素E各组分的含量。结果表明, 不同基因型甜玉米材料间维生素A源和维生素E各组分及α/γ-生育酚存在显著差异。非维生素A源的叶黄素和玉米黄质为维生素A组分中的主要成分, γ-生育酚为维生素E组分中的主要成分。相关分析表明多数性状间表现为显著正相关或不相关, 其中β-胡萝卜素极显著正相关于玉米黄质、β-隐黄质和α-类胡萝卜素, 维生素E各组分两两极显著相关, α/γ-生育酚极显著负相关于δ-生育酚和γ-生育酚, 极显著正相关于玉米黄质和α-生育酚, β-胡萝卜素与α-生育酚呈正相关。这些数据为进一步研究甜玉米籽粒中维生素A源和维生素E的代谢和微营养品质的改良提供了有益的表型数据。
Vitamin A and Vitamin E are two kinks of micronutrients that must be acquired regularly from dietary sources. Sweet corn is a type of fresh corn for direct consumption by human beings. The content of provitamin A and vitamin E components at milk stage of kernels development were measured by HPLC in 47 sweet corn lines. The results showed that the contents of carotenoids and tocopherols and the α/γ tocopherol ratio were extremely different among 47 sweet corn lines. The non-provitamin A, including lutein and zeaxanthin, are the main components of the total carotenoids. The main component of vitamin E is γ-tocopherol. The pearson correlation analysis showed that significantly positive correlation and non-correlation were detected among most traits. β-carotene was significantly positive correlated with zeaxanthin, β-cryptoxanthin and α-carotenoids, respectively. Three isomers of vitamin E were significantly correlated between each other. The ratio of α/γ tocopherol had a significantly negative correlation with δ-tocopherol and γ-tocopherol and a significantly positive correlation with zeaxanthin and α-tocopherol. A significantly positive association was found between β-carotene and α-tocopherol. These results provide useful information for the research on provitamin A and vitamin E metabolism and nutritional improvement in sweet corn.
维生素A和维生素E均为脂溶性维生素, 是动物和人类必不可少的微量营养物质。维生素A在抗氧化及维持正常的视觉反应、上皮组织形态与功能、骨骼发育和皮肤细胞功能等方面具有重要的功能[ 1]。缺乏维生素A, 会使上皮细胞的功能减退, 导致皮肤弹性下降, 变干和粗糙, 失去光泽, 严重的维生素A缺乏症可导致失明甚至死亡[ 2]。维生素E在繁殖生育、免疫、抗衰老、抗氧化等方面具有重要功能[ 3, 4]。植物组织中并不直接含有维生素A, 而以维生素A源——类胡萝卜素(carotenoid)的形式存在, 维生素A源能在人体内转变为具有生理活性的维生素A。天然维生素E的结构包含一个芳香环和一个疏水性侧链, 根据侧链饱和度不同分为生育酚(tocopherol)和生育三烯酚(tocotrienol)两类。根据芳香环甲基数目和位置的不同又各分为4种生育酚(α、β、γ和δ)和生育三烯酚(α、β、γ和δ)[ 5]。
植物性食物中, 如绿叶菜类、黄色菜类以及水果中含有较丰富的维生素A源, 粮食作物中玉米含较丰富的维生素A源和维生素E。玉米籽粒包含5种主要的类胡萝卜素, 即α-胡萝卜素、β-胡萝卜素、β-隐黄质、叶黄素和玉米黄质, 95%~97%都位于胚乳中, 仅有1%左右位于胚中[ 6]。α-胡萝卜素、β-胡萝卜素和β-隐黄质具有维生素A源活性, β-胡萝卜素的活性是α-胡萝卜素和β-隐黄质的2倍, 无维生素A源活性的叶黄素、玉米黄质是眼睛视网膜黄斑区域形成的必要成分[ 7, 8]。玉米籽粒中的生育酚有α-生育酚、β-生育酚、γ-生育酚和δ-生育酚, 主要位于胚中[ 6], 其中β-生育酚的含量可以忽略不计[ 9]。已有大量关于普通玉米维生素A源和维生素E的研究[ 10, 11, 12, 13], 而以甜玉米为材料的研究较少[ 14]。
玉米中与维生素A和维生素E相关的研究以普通玉米为多, 而对本应受到重视的鲜食玉米研究较少。本研究选用超甜玉米自交系( sh 2 sh 2)为材料, 采用高效液相色谱(HPLC)法分析了乳熟期籽粒中维生素A源和维生素E各组分的含量, 以期明确甜玉米自交系中维生素A源和维生素E各组分的变异, 为甜玉米营养品质改良提供依据。
选用47份甜玉米自交系(基因型为 sh 2 sh 2, 编号A1~A47), 来源于国内外引进的单交种选育的二环系, 由华南农业大学甜玉米课题组选育。2009年种植于华南农业大学增城教学科研基地, 每自交系种3次重复, 每重复种2行, 每行10株(株距30 cm, 行距 80 cm), 按常规方法进行田间管理。自交授粉后第21天(甜玉米最佳采收期), 从每行选取5个无病虫害感染的果穗, 于37℃烘干后脱粒、混匀。以四分法取50粒左右的样品, 万能粉碎机粉碎1 min 40 s, 将磨好的样品转移到自封袋中, 于4~6℃避光保存用于测定类胡萝卜素和维生素E各组分, 每样品测定3次。
α-生育酚、γ-生育酚和δ-生育酚标样购自sigma公司, β-胡萝卜素、叶黄素、玉米黄质标样由HarvestPlust项目提供, α-胡萝卜素和β-隐黄质标样则是利用柱层析法分别从胡萝卜和青玉米中提取的。其他化学试剂均购自上海生工生物工程有限公司, 为分析纯。
在中国农业大学国家玉米改良中心提取及测定类胡萝卜素各组分(α-胡萝卜素、β-胡萝卜素、β-隐黄质、叶黄素和玉米黄质)和生育酚各组分(α-生育酚、γ-生育酚和δ-生育酚), 参照Chander等[ 13]实验方法。
称取0.6 g甜玉米粉于25 mL螺口玻璃离心管中; 加入6 mL 0.1%的BHT (0.1 g BHT溶于100 mL无水乙醇), 涡旋30 s, 85℃水浴15 min, 每5 min涡旋一次, 第一次水浴5 min后加入500~1500 μL 80%的KOH溶液, 取出后置冰上, 加入3 mL预冷的ddH2O (加入200 μL纯化后的内标apo-8’-carotenel, 涡旋), 再加入3 mL的正己烷, 涡旋混匀, 2700转 min-1离心1~2 min, 转移上层有机相于新的螺口离心管中, 再在原离心管中加入3 mL正己烷, 涡旋、离心; 重复转移上层有机相3次以上, 最后一次要使上层有机相完全转移; 加入3 mL ddH2O; 涡旋后静置, 转移上层有机相于20 mL敞口锥形底玻璃离心管中; 在水相中加入1~2 mL正己烷, 涡旋、离心, 转移上层有机相; 重复以上步骤2次; 于真空干燥离心机中浓缩1.0~1.5 h; 加入1 mL乙腈∶甲醇∶二氯甲烷(45∶20∶35, v/v/v)溶剂; 用1 mL一次性注射器使溶质完全溶解后, 通过0.22 μm的过滤器转移至 2 mL棕色进样小瓶中; 用于液相色谱分析。
采用高压液相色谱法分离样品类胡萝卜素各组分, 基于外标法构建的回归方程对样品类胡萝卜素各组分含量进行量化分析。采用日本Shimadzu公司2001高效液相色谱仪, 色谱柱为反相YMC类胡萝卜素C30 柱子(5 μm, 4.6 mm × 250.0 mm) (Waters Chromatography, Milford, MA)。流动相A为乙腈 (75 V), 流动相B为甲醇/二氯甲烷(20∶5, v/v), 含0.05% (在流动相总量中的比例)的BHT抗氧化剂。采用等度洗脱, 流速是2 mL min-1。柱温控制在30℃, 检测波长为450 nm和295 nm。
采用Agilent 1200色谱数据工作站记录和分析HPLC的测定结果, 以1/2 (β-隐黄质+α-胡萝卜素)+ β-胡萝卜素含量为维生素A源含量, 5种组分的总和为总类胡萝卜素含量; 维生素E含量为α-生育酚、γ-生育酚和δ-生育酚含量之和。用SAS8.0进行方差分析和相关分析。
图1-A表明, 在本实验所用的色谱条件下可以同时将类胡萝卜素的5种组分分开, 混合标样的洗脱依次为叶黄素、玉米黄质、β-隐黄质、α-胡萝卜素和β-胡萝卜素。标样的纯度依次为叶黄素89.7%、玉米黄质97.1%、β-隐黄质96.5%、α-胡萝卜素99.5%和β-胡萝卜素99.7%, 用外标法所建立的回归方程决定系数均在0.99以上, 说明标样可用, 定量准确。图1-B表明, 本实验所用的色谱条件可以同时将玉米中生育酚的各组分分开, 混合标样的洗脱依次为δ-生育酚、γ-生育酚和α-生育酚。用外标法所建立的回归方程决定系数均在0.99以上, 说明标样可用, 生育酚各分量定量准确。
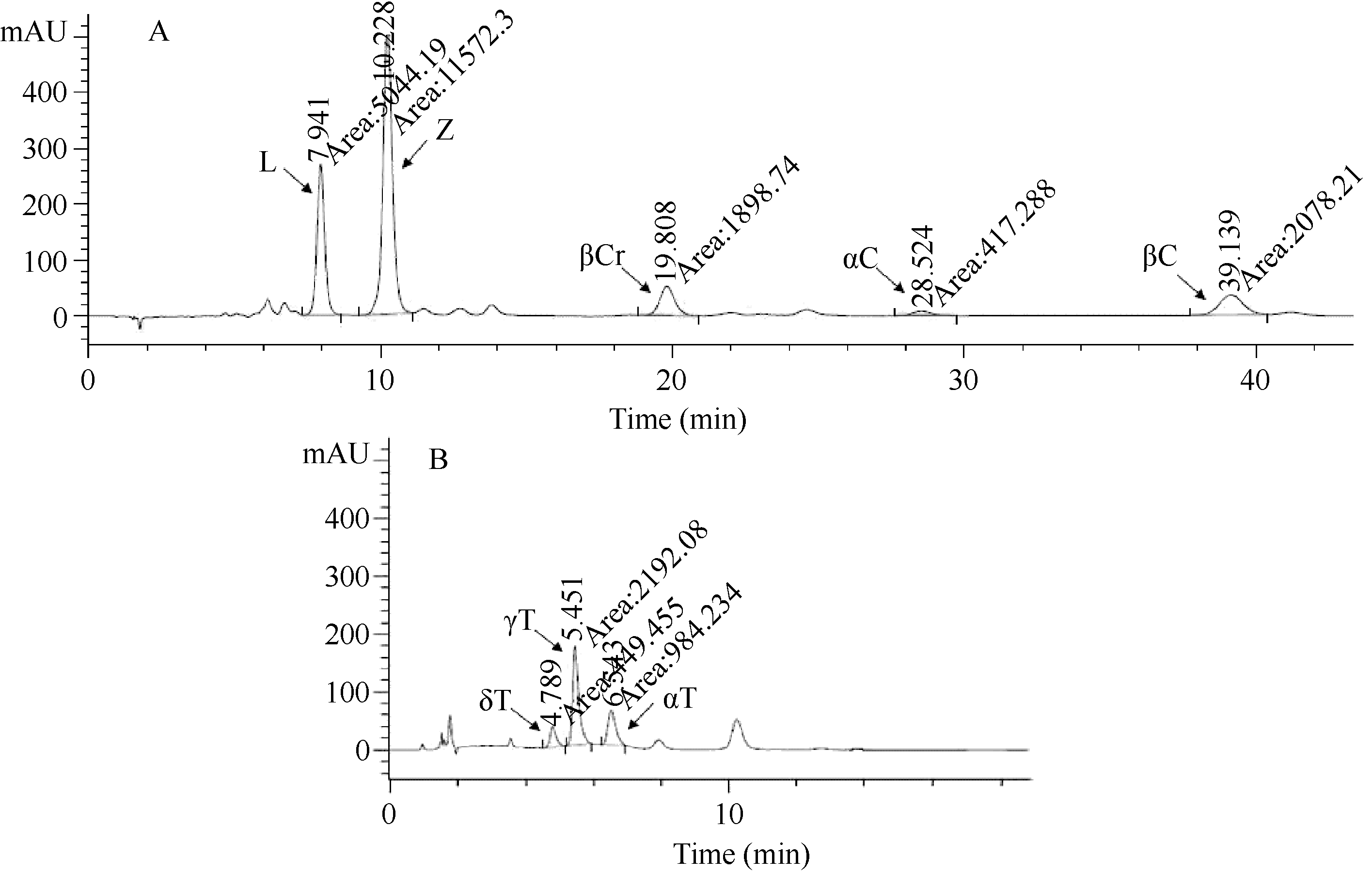 | 图1 类胡萝卜素和生育酚各组分的HPLC分析Fig. 1 HPLC analysis for carotenoids and tocopherols |
对47份甜玉米自交系乳熟期籽粒类胡萝卜素各组分的定量分析(表1和表2)表明, α-胡萝卜素、β-胡萝卜素、β-隐黄质、玉米黄质、叶黄素含量的均值分别为0.52、0.88、4.28、7.43和15.83 μg g-1, 变异范围分别是0.03~3.85、0.04~4.48、0.07~35.48、0.31~19.07、0.99~43.52 μg g-1。维生素A源含量均值为3.282 μg g-1, 变异范围为0.07~22.75 μg g-1, 变异系数达1.867, 最高与最低间相差307.5倍, 含量高于15 μg g-1的材料有2个, 即A5和A37。总类胡萝卜素含量均值为28.94 μg g-1, 变异范围为1.44~75.98 μg g-1, 最高与最低间相差52.7倍, 变异系数达0.548。
| 表1 47份甜玉米自交系维生素A源及类胡萝卜素各组分的含量平均值 Table 1 Average content of pro-vitamin A and carotenoid components in 47 sweet corn inbred lines (μg g-1) |
| 表2 47份甜玉米自交系中类胡萝卜素和维生素E各组分间差异分析 Table 2 Comparison of components of carotenoids and tocopherols in 47 sweet corn lines (μg g-1) |
图2和表2表明, α-生育酚、γ-生育酚、δ-生育酚含量的变幅分别为1.69~58.03、3.61~68.80和0~ 2.01 μg g-1, 维生素E含量的均值为51.15 μg g-1, 变异范围为18.50~99.17 μg g-1, 最高与最低间相差5.4倍, 变异系数为0.35, 含量最高的2个材料为A5和A6, 其总生育酚含量分别达90.46 μg g-1和99.17 μg g-1。α/γ-生育酚的均值为1.86, 变异范围为0.05~7.85, 变异系数为1.11。其中A5维生素A源和维生素E的含量均较高。
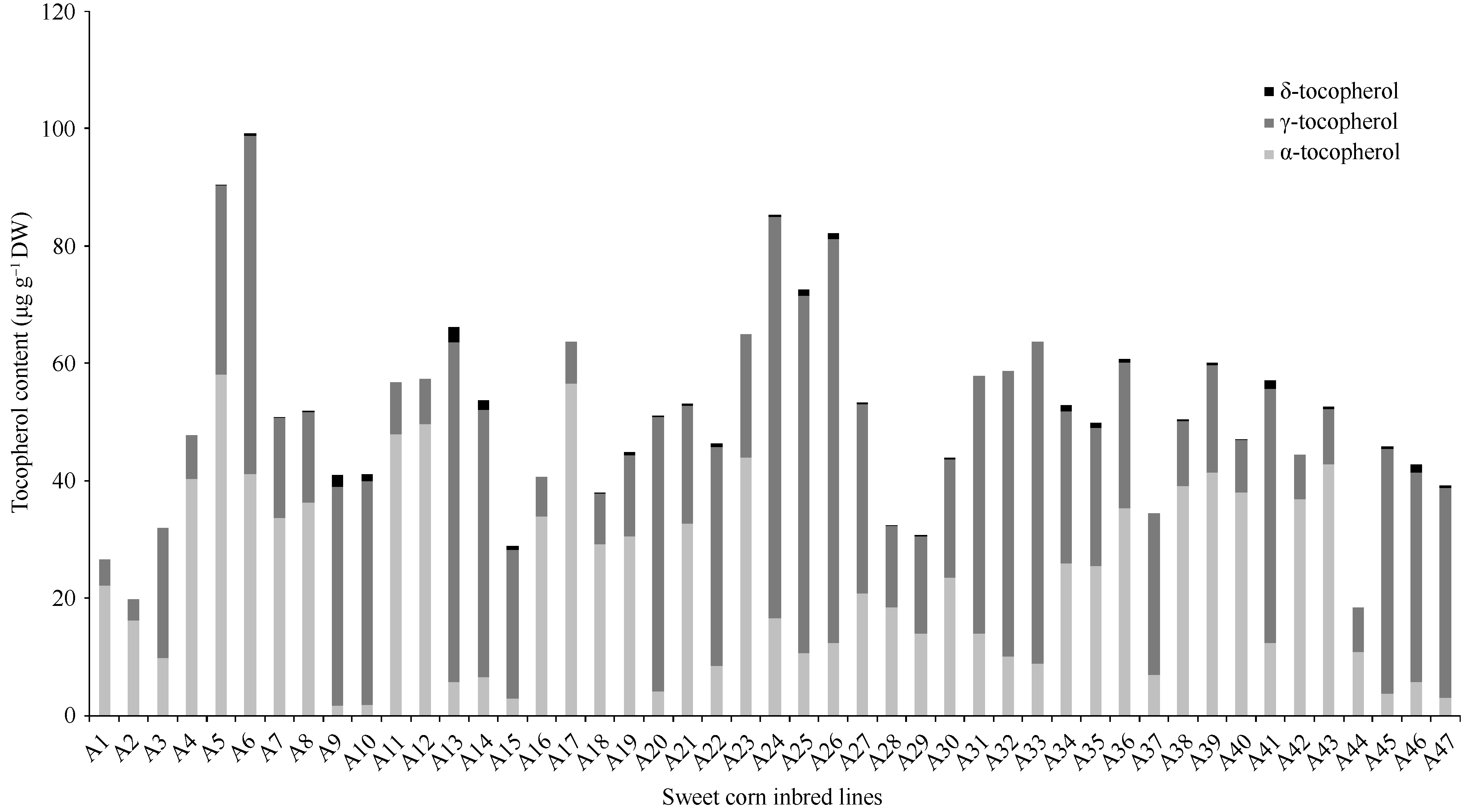 | 图2 47份甜玉米自交系籽粒乳熟期生育酚各组分的浓度Fig. 2 Content of tocopherols at milk stage of kernel development in 47 sweet corn lines |
从表2可知类胡萝卜素各组分含量从高到低依次为叶黄素、玉米黄质、β-隐黄质、β-胡萝卜素、α-胡萝卜素, 叶黄素和玉米黄质是类胡萝卜素的主要组成成分, 叶黄素、玉米黄质和β-隐黄质间的差异极显著, 而β-胡萝卜素和α-胡萝卜素之间差异不显著。维生素E各组分含量从高到低依次为γ-生育酚、α-生育酚、δ-生育酚, γ-生育酚和α-生育酚是维生素E的主要组成成分, 极显著高于δ-生育酚。偏度及峰度分析发现玉米黄质、叶黄素及总类胡萝卜素的偏度值和峰度值接近于1, 符合正态分布; 维生素A源和β-胡萝卜素不符合正态分布。维生素E各组分的分析发现α-生育酚、γ-生育酚和维生素E含量的偏度值和峰度值接近于1, 符合正态分布。
由表3可知, 各组分间差异极显著。类胡萝卜素组分中α-胡萝卜素的均方和最小, 为1.38, 总类胡萝卜素的均方和最大, 为753.49, 维生素A源的均方和为56.41; 维生素E各组分中δ-生育酚的均方和最小, 为1.05, γ-生育酚的均方和最大, 为1015.34, 维生素E的均方和为849.62。47份甜玉米自交系中维生素A活性最强的成分β-胡萝卜素的均方和仅为2.54, 表明现有甜玉米材料中的β-胡萝卜素变异极不丰富。
| 表3 47份甜玉米自交系类胡萝卜素和维生素E组分的方差分析 Table 3 ANOVA analysis of carotenoids and tocopherols among 47 sweet corn lines (μg g-1) |
表4显示, 在这47份甜玉米材料中, 维生素A活性最高的β-胡萝卜素极显著正相关于玉米黄质、β-隐黄质、α-类胡萝卜素、维生素A源和总类胡萝卜素, 仅与玉米黄质相关性不显著。维生素A源与其各组分(β-胡萝卜素、α-胡萝卜素和β-隐黄质)呈极显著正相关, 还与玉米黄质极显著正相关。所有类胡萝卜素组分均与总类胡萝卜素呈极显著正相关, α-胡萝卜素与叶黄素极显著正相关、与β-隐黄质显著正相关, β-隐黄质与玉米黄质极显著正相关。
| 表4 47份甜玉米自交系类胡萝卜素和生育酚各组分的相关系数 Table 4 Pearson correlation coefficients between carotenoids and tocopherols |
维生素E各组分两两之间的相关性均达到极显著, γ-生育酚分别与δ-生育酚和α-生育酚呈极显著正相关, δ-生育酚与α-生育酚呈极显著负相关。总生育酚分别与α-生育酚和γ-生育酚显著和极显著正相关, 相关系数为0.3567和0.5959, 而δ-生育酚与总生育酚间不相关。α/γ-生育酚的比值与α-生育酚呈极显著正相关, 与δ-生育酚和γ-生育酚呈极显著负相关。
尽管类胡萝卜素主要存在于胚乳中, 维生素E主要存在于胚中, 但两者的合成途径中有共同的前体物质, 检测到玉米黄质分别与α-生育酚和α/γ-生育酚比值呈极显著正相关, 叶黄素与δ-生育酚呈显著负相关, β-胡萝卜素与α-生育酚呈显著正相关。
玉米是世界上播种面积最大、总产最高的粮食作物, 除了作为饲料, 还有相当多的人以玉米为主粮。甜玉米是一种鲜食玉米类型直接食用, 其种植面积逐年扩大, 营养品质与人的健康息息相关, 维生素A源和维生素E是甜玉米重要的微营养物质。本研究发现甜玉米中叶黄素和玉米黄质是类胡萝卜素的主要组成成分, 维生素A源较低, γ-生育酚和α-生育酚是维生素E的主要组成成分, δ-生育酚的含量较低, 本研究的结果与普通玉米的研究结果相同[ 7, 12, 13, 15, 16]。
在本研究中, 对授粉后第21天, 即最佳采收期的甜玉米籽粒的类胡萝卜素及维生素E各组分进行了分析, 而普通玉米提取的是成熟玉米籽粒的类胡萝卜素及维生素E含量[ 12, 13, 15], Chander等[ 13]的研究发现, 87份普通玉米成熟籽粒中β-胡萝卜素含量均值为0.449 μg g-1, 变异范围为0.016~1.736 μg g-1, 总类胡萝卜素含量均值为10.298 μg g-1, 变异范围为0.096~22.495 μg g-1, 本研究中甜玉米类胡萝卜素的含量明显高于普通玉米; 在Chander等[ 13]的研究中, 普通玉米的α-生育酚含量均值为23.98 μg g-1, 变幅为3.98~74.81 μg g-1, 总生育酚含量均值为59.55 μg g-1, 变幅为19.60~120.69 μg g-1, 与本研究中甜玉米生育酚含量相比差异不大。Kurilich等[ 14]以6份 sh 2 sh 2基因型的甜玉米为材料, 在授粉后20 d进行类胡萝卜素及维生素E各组分的定量分析, 除β-胡萝卜素略高于本研究中的群体均值外, 其余组分均低于本研究群体均值; 两种主要维生素E组分α-生育酚和γ-生育酚的含量均低于本研究群体均值, 本研究所用材料变异更为丰富。
对于育种家而言, 任何需改良的性状的变异越小越不利于改良, 均方和可用于评价变异的大小, 变异越大越有改良的空间, 否则须引入新材料或采用其他的方法来改良。在本研究中, 47份甜玉米维生素A源组分的均方和较小, 表明这些甜玉米材料中维生素A源变异并不丰富, 引进高维生素A源的材料显得尤为重要; 而维生素E组分中γ-生育酚和α-生育酚的均方和较大, 表明甜玉米中的维生素E的变异较丰富, 可在甜玉米材料中选育到高维生素E的材料。普通玉米中已经鉴定出高维生素A源和高维生素E的材料, 克隆了维生素A源合成的限速基因 LCYE[ 11]和 crtRB1[ 17]并开发了功能标记, 也鉴定了与维生素E合成相关的基因 ZmVTE4[ 18], 为强化甜玉米的维生素A源和维生素E奠定了坚实的基础。
维生素A源和维生素E是甜玉米的两类重要微营养成分, 理解类胡萝卜素和生育酚各组分间的相关性对改良甜玉米材料的营养性状十分重要。在本研究中检测到β-胡萝卜素分别与玉米黄质、β-隐黄质和α-胡萝卜素呈极显著正相关, 而α-生育酚、γ-生育酚和δ-生育酚两两之间均达到极显著相关。在普通玉米中, Chander等[ 13]未检测到β-胡萝卜素与α-胡萝卜素间及维生素E各组分间的相关性, 也未检测到β-胡萝卜素与α-生育酚的相关。β-胡萝卜素具有较高的维生素A源活性, 1分子的β-胡萝卜素可转化生成2分子的维生素A[ 7]; 维生素E各组分中α-生育酚具有更高的活性, 因为肝脏中有专化的α-生育酚转运酶[ 19], 而甜玉米中发现β-胡萝卜素和α-生育酚间存在正相关, 可见同时提高甜玉米中这两种微营养物质是可行的。
甜玉米中类胡萝卜素成分主要含叶黄素和玉米黄质, 而维生素A源含量较低, γ-生育酚和α-生育酚是维生素E的主要成分。不同材料间叶黄素、玉米黄质和β-隐黄质的变幅较大, α-胡萝卜素和β-胡萝卜素的变幅较小, 维生素E组分中α-生育酚和γ-生育酚的变幅较大, δ-生育酚的变幅较小, 不同材料间类胡萝卜素和维生素E组分均差异极显著。β-胡萝卜素极显著正相关于玉米黄质、β-隐黄质、α-类胡萝卜素、维生素A源和总类胡萝卜素, 与玉米黄质不相关, α-胡萝卜素与叶黄素、β-隐黄质与玉米黄质呈极显著正相关。δ-生育酚、α-生育酚和γ-生育酚两两相关极显著, 总生育酚与α-生育酚和γ-生育酚分别呈显著和极显著正相关, 而与δ-生育酚不相关, 还检测到β-胡萝卜素与α-生育酚呈显著正相关。这些结果为今后甜玉米维生素A源和维生素E营养品质改良提供了有益的表型数据基础。
致谢: 特别感谢中国农业大学国家玉米改良中心李建生教授、华中农业大学严建兵教授对本实验的指导与支持。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|