

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
栽培模式对杂交粳稻常优5号根系形态生理性状和地上部生长的影响
[褚光1  , 周群
, 周群1 , 薛亚光1, 3 , 颜晓元2 , 刘立军1 , 杨建昌1, * ]
, 周群]
|
|


第一作者联系方式: E-mail:chuguang19880210@163.com
以杂交粳稻常优5号为材料, 设置未施氮肥处理(0N)、当地高产栽培(对照)、超高产栽培和氮肥高效利用栽培等4种栽培模式, 观察其对水稻不同生育期根系形态生理和地上部生长的影响。结果表明, 不同栽培模式下水稻产量差异极显著。超高产栽培与氮肥高效利用栽培两年的平均产量分别为12.29 t hm-2和9.62 t hm-2, 平均分别较对照增产41.4%和10.7%。上述两种栽培模式的氮肥农学利用率(每kg施氮量增加的产量)分别较对照增加80.7%和76.8%, 灌溉水利用效率分别较对照提高62.1%和32.3%。与对照相比, 超高产栽培与氮肥高效利用栽培均增加了水稻地上部干物重、叶面积指数、根干重、根长, 提高了粒叶比, 改善了库源关系, 并提高了根冠比与根系伤流量。同时也提高了灌浆期剑叶净光合速率、根系氧化力、根系总吸收表面积与根系活跃吸收表面积, 生育中后期根系、叶片以及根系伤流液中的玉米素(Z)与玉米素核苷(ZR)含量、灌浆期籽粒中蔗糖合酶(SuSase)以及腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)活性。这说明, 通过栽培技术的集成与优化可以提高水稻灌浆期根系和地上部的生理活性, 促进水稻高产与水分养分高效利用。
The objective of this study was to investigate if a cultivation technique could coordinately increase both grain yield and nutrient use efficiency of rice through an improvement in morphological and physiological traits of roots. A field experiment was conducted using
近年来, 我国开展了作物超高产研究, 涌现了许多超高产记录。然而, 大多数高产记录是在高水、肥投入下实现的。部分高产记录重演性较差, 在地区间或年度间产量表现出严重的不稳定性[ 1, 2, 3]。究其原因, 可能与超高产水稻的生长发育规律不清楚、生育诊断指标不明确、超高产栽培技术尚未完全掌握等有关。
随着我国水稻产量的不断攀升, 氮肥的施用量也在持续增加。2010年, 我国化肥消费量达到5562万吨, 约占世界化肥总消费量的34%, 其中氮肥约为3200万吨, 为世界第一消费大国[ 4]。氮肥投入量高、利用效率低是我国目前水稻生产中的一个突出问题。有研究表明, 我国稻田氮肥利用率仅为30%~50%, 在部分高产和高投入区域, 氮肥利用效率则更低[ 5, 6, 7, 8]。如何在获得高产的同时, 实现水分与养分的高效利用?这是目前农业研究的一个热点问题。
根系作为水稻植株的重要组成部分, 不仅能够吸收养分和水分, 同时也可合成许多重要的生理活性物质, 因而在水稻生长发育的过程中起着十分重要的作用[ 9, 10, 11]。近年研究表明, 水稻根系的生长发育与各种环境条件密切相关, 其中尤以水分、肥料、耕作栽培措施等影响较大[ 12, 13, 14]。目前对超高产栽培和氮肥高效利用栽培下水稻地上部研究较多[ 15, 16, 17], 但对其地下部生长发育以及根冠相互作用机制的研究较少。本研究观察了杂交粳稻常优5号在超高产栽培与氮肥高效利用栽培下根系形态生理特征的变化及其与地上部生长发育的关系, 以期从根系生长发育角度进一步认识水稻的高产与水分养分高效利用的机制。
试验于2012—2013年在中国科学院南京土壤研究所常熟野外生态试验站进行。试验地前茬作物为小麦。耕作层含有机质3.16%~3.18%、全氮0.17%~0.19%、有效氮82.70~83.24 mg kg-1、速效磷26.5~27.3 mg kg-1、速效钾110.7~111.6 mg kg-1。供试品种为当地高产品种杂交粳稻常优5号。5月19日至20日播种, 6月17日至18日移栽, 双本栽插。于8月30日至9月3日抽穗, 10月25日至11月2日收获。
1.2.1 0N 不施氮肥, 仅在移栽前基施磷肥(过磷酸钙, 含P2O5 13.5%) 90 kg hm-2和钾肥(氯化钾, 含K2O 60%) 120 kg hm-2, 栽插株行距为20 cm×20 cm。除生育中期排水搁田外, 其余时期保持水层至收获前1周断水。
1.2.2 LHY即当地高产栽培(对照) 总施氮量(纯氮, 以下同)为300 kg hm-2, 按基肥∶分蘖肥∶穗肥 = 6∶2∶2施用, 磷、钾肥的用量、栽插株行距及水分管理方式同0N处理。
1.2.3 SHY即超高产栽培 氮肥较LHY增25%, 即375 kg hm-2; 磷肥较LHY处理增加40%, 施用量为126 kg hm-2; 钾肥较LHY增加50%, 施用量为180 kg hm-2; 基肥增施硅肥225 kg hm-2, 锌肥15 kg hm-2, 菜籽饼肥2250 kg hm-2。
1.2.4 HUEN即氮肥高效利用栽培(施用含有硝化抑制剂的尿素) 氮肥较LHY减25%, 即225 kg hm-2。磷肥和钾肥的施用量及时间同SHY处理。
各处理施肥时期与施肥量如表1所示。
| 表1 水稻不同栽培模式的施肥时期和施肥量 Table 1 Time and amount of fertilizer application for different cultivation patterns (kg hm-2) |
各小区用拖拉机旋耕, 耕深8~10 cm, 上水后耱平。每处理重复4次, 小区面积42 m2, 小区之间筑田埂并用塑料薄膜覆盖, 以防肥水串灌。
针对超高产栽培(SHY)和氮肥高效利用栽培(HUEN)采用以下关键技术。
I.稀播育壮秧, 增加栽插密度。湿润育秧, 播量较LHY处理减少15%。移栽时秧苗带分蘖2~3个, HUEN株行距为20 cm×15 cm, 栽插密度较LHY处理增加33%, SHY株行距与LHY相同。
II.前氮后移。氮肥按基肥∶分蘖肥∶促花肥∶保花肥 = 5∶1∶2∶2施用。磷肥于移栽前作基肥一次性施入。钾肥分基肥和拔节肥两次施用, 前后两次的比例为5∶5。
III.精确灌溉。从移栽至返青建立浅水层; 返青至有效分蘖临界叶龄期(N-n)前2个叶龄期(N-n- 2)进行间隙湿润灌溉, 低限土壤水势为-10 kPa; (N-n-1)叶龄期至(N-n)叶龄期进行排水搁田, 低限土壤水势为-20 kPa, 并保持1个叶龄期; (N-n+1)叶龄期至二次枝梗分化期初(倒三叶开始抽出)进行干湿交替灌溉, 低限土壤水势为-25 kPa; 二次枝梗分化期(倒三叶抽出期)至出穗后10 d进行间隙湿润灌溉, 低限土壤水势为-10 kPa; 抽穗后11 d至抽穗后45 d进行干湿交替灌溉, 低限土壤水势为-15 kPa。各生育期达到上述指标即灌2~3 cm浅层水, 用水分张力计监测土壤水势。
1.4.1 叶片形态与生理特性的测定 分别于分蘖中期、穗分化始期、抽穗期、成熟期考察每个小区100穴植株的分蘖数, 按照平均茎蘖数取10穴植株, 分解为绿叶、枯叶、茎、鞘和穗(抽穗以后), 烘干测定干物质重。于以上时期各小区取2个8穴, 采用美国LI-COR公司生产的Li-Cor 3050型叶面积仪测定叶面积。并于抽穗期、抽穗后8、17、26 d, 选择晴朗上午, 于9:00采用美国LI-COR公司生产的LI-6400便携式光合测定仪测定稻株最上展开叶的光合速率。叶室CO2浓度为380 μmol mol-1, 使用红蓝光源, 光量子通量密度(PFD)为1400 μmol m-2 s-1, 温度28~30℃, 各处理重复测定8张叶片。
1.4.2 根系形态与生理特性的测定 分别于分蘖中期、穗分化始期、抽穗期、成熟期, 取5穴稻株, 每穴稻株以基部为中心, 挖取20 cm × 20 cm × 20 cm的土块, 装于70目的筛网袋中, 先用流水冲洗, 然后用农用压缩喷雾器将根冲洗干净, 放入装有一薄层去离子水的玻璃皿中(30 cm × 30 cm), 用镊子调整根的位置以尽量避免交叉重叠, 先用扫描仪(Epson Expression 1680 Scanner, Seiko Epson Corp. Tokyo, Japan)扫描图像, 后用WinRHIZO根系分析系统(Regent Instruments Inc. Quebec, Canada)分析和计算, 测得根长和根直径并于烘干后测干重。分别于抽穗期、抽穗后8、17、26 d, 用上述取根方法, 取3穴参照杨建昌等[ 18]方法测定根系氧化力。另取3穴采用甲烯蓝蘸根法[ 19]测定根系总吸收表面积与根系活跃吸收表面积。另分别于上述时期, 取代表性植株6穴, 于18:00在各茎离地10 cm处剪去地上部分植株, 将预先称重的脱脂棉放于茎的剪口处, 包上塑料薄膜, 于第2天早6:00取回带有伤流液的脱脂棉并称重, 挤出伤流液测定其Z+ZR含量。参照陈远平等[ 20]的高效液相色谱法并作改进提取、纯化和定量分析根系、叶片以及根系伤流液中激素: 用石油醚萃取去除样品中的叶绿素和脂肪等物质, 经Sep-Pak C18柱过滤以减少样品中杂质; 色谱条件改用Dubhe C184.6×250, 5 μm, 流动相为5% (v/v)乙腈、50% (v/v)甲醇、0.6% (v/v)冰乙酸, 流速为0.8 mL min-1, 采用梯度洗脱法, 检测波长254 nm; 柱温30℃, 进样量20 μL。样品回收率为85.5%±2.6%, 每一个样品至少重复4次。以外标法定量。
1.4.3 籽粒中蔗糖合酶(SuSase)和腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)活性测定 分别于抽穗后8、17和26 d, 取30~40粒去壳籽粒, 加3~5 mL 100 mmol L-1 Tricine-NAOH提取液[pH 8.0, 含10 mmol L-1 MgCl2, 2 mmol L-1 EDTA, 50 mmol L-1 2-mercap-toethanol, 12% (v/v) glycerol, 5% (w/v) PVP 40]于研钵中研磨(温度保持在0℃), 15 000 × g离心 5 min (4℃), 参照Yang等[ 21, 22]方法, 略作改进, 取上清液(粗酶液)测定各酶活性。各酶活性单位用μmol g-1 FW min-1表示。
1.4.4 考种与计产 成熟期从各小区取50穴考察每穴穗数, 取10穴观察结实率(水漂法, 沉入水底者为饱粒)和千粒重。
氮吸收利用率(%)=(施氮区作物吸氮量-氮空白区作物吸氮量)/施氮量;
氮农学利用率(kg 稻谷 kg-1 N) = (施氮区产量-氮空白区产量)/施氮量;
氮生理利用率(kg 稻谷 kg-1 N) = (施氮区籽粒产量-氮空白区籽粒产量)/(施氮区植株吸氮量-氮空白区植株吸氮量);
偏生产力(kg 稻谷 kg-1 N) = 水稻产量/施肥量;
氮收获指数=成熟期单位面积植株穗部氮积累量/植株氮素积累总量;
水分利用效率(kg 稻谷 m-3用水量) = 水稻产量/灌溉用水量;
采用Microsoft Excel 2003、SPSS16.0和SAS统计软件分析试验数据, 用SigmaPlot 10.0绘图。
0N处理、当地高产栽培(对照)、超高产栽培以及氮肥高效利用栽培两年(2012年和2013年)的平均产量分别为5.82、8.69、12.29、9.62 t hm-2。超高产栽培和氮肥高效利用栽培分别较对照增产41.43%与10.70%。总颖花量的显著增加是其产量增加的主要原因。超高产栽培和氮肥高效利用栽培的总颖花数分别较对照增加43.07%与12.42%。0N处理下的结实率显著高于当地高产栽培(对照)、超高产栽培以及氮肥高效利用栽培, 但超高产栽培和氮肥高效利用栽培下的结实率与对照无显著差异(表2)。
| 表2 不同栽培模式下的水稻产量及其构成因素 Table 2 Grain yield and its yield components under different cultivation patterns |
超高产栽培和氮肥高效利用栽培下的氮肥的偏生产力、吸收利用率以及农学利用率均显著高于当地高产栽培(对照)。氮肥高效利用栽培的氮肥生理利用率也显著高于对照; 但超高产栽培的氮肥生理利用率显著低于氮肥高效利用栽培而且与对照差异不显著, 表明超高产栽培促进了稻株对氮素的吸收, 但吸收的氮素更多地集中在稻草中, 而转化为经济产量的效率仍然较低, 造成水稻对氮素的奢侈吸收。超高产栽培的氮收获指数低于其余3个处理, 也进一步证明这一点(表3)。
| 表3 不同栽培模式下水稻的吸氮量、氮收获指数与氮肥利用效率 Table 3 N uptake, N harvest index, and N use efficiency under different cultivation patterns in rice |
超高产栽培和氮肥高效利用栽培两年的平均灌溉用水量分别是4860 m3 hm-2与4661 m3 hm-2, 是当地高产栽培(对照)的87%和84%。超高产栽培和氮肥高效利用栽培的灌溉水分利用效率分别为2.53 kg m-3与2.06 kg m-3, 分别较对照增加62%与32% (表4)。
| 表4 不同栽培模式下的水稻灌溉水利用效率 Table 4 Water use efficiency (WUE) for irrigation under different cultivation patterns in rice |
超高产栽培和氮肥高效利用栽培在抽穗期的总叶面积指数、有效叶面积指数、高效叶面积指数均显著高于当地高产栽培(对照), 超高产栽培分别较对照高22.65%、39.75%和53.30%; 氮肥高效利用栽培分别较对照高4.44%、14.84%和18.85% (表5)。从表5可以看出, 超高产栽培与氮肥高效利用栽培两年抽穗期颖花数/叶面积(粒叶比)显著高于对照; 实粒数/叶面积与对照互有高低; 粒重/叶面积则与对照无显著差异。
| 表5 不同栽培模式下的水稻抽穗期叶面积指数和粒叶比 Table 5 Leaf area index and grain-leaf ratio at heading stage under different cultivation patterns in rice |
不同栽培模式下水稻的剑叶净光合速率在抽穗后均呈现下降的趋势。在抽穗期, 超高产栽培的剑叶净光合速率显著高于当地高产栽培(对照), 而氮肥高效利用栽培与对照无显著差异。自灌浆早期至灌浆后期, 超高产栽培和氮肥高效利用栽培模式的剑叶净光合速率均显著高于对照(图1)。
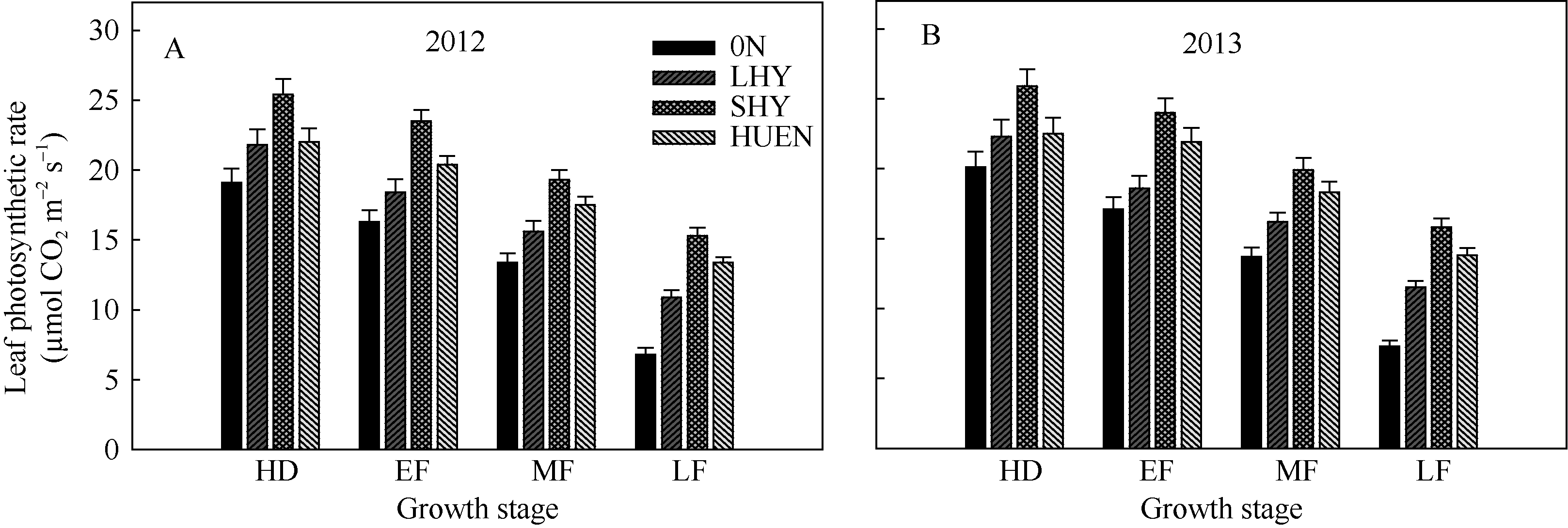 | 图1 不同栽培模式下水稻在不同生育时期的剑叶净光合速率Fig. 1 Flag leaf photosynthetic rate under different cultivation patterns in rice at different growth stages |
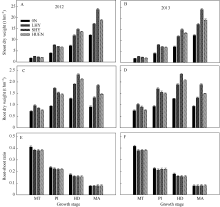
在所测定的生育期内, 0N处理的地上部干物重(图2-A, B)和根干重(图2-C, D)均显著低于当地高产栽培、超高产栽培和氮肥高效利用栽培3个处理, 而其根冠比(图2-E, F)则在分蘖中期显著高于其他3个处理。在分蘖中期和穗分化始期, 当地高产栽培(对照)的地上部干物重与根干重均显著或极显著高于超高产栽培或氮肥高效利用栽培; 在抽穗期和成熟期则极显著低于超高产栽培或氮肥高效利用栽培。不同栽培模式下水稻的根冠比均随生育进程逐渐减小, 在整个生育期超高产栽培、氮肥高效利用栽培的根冠比与对照无显著差异。
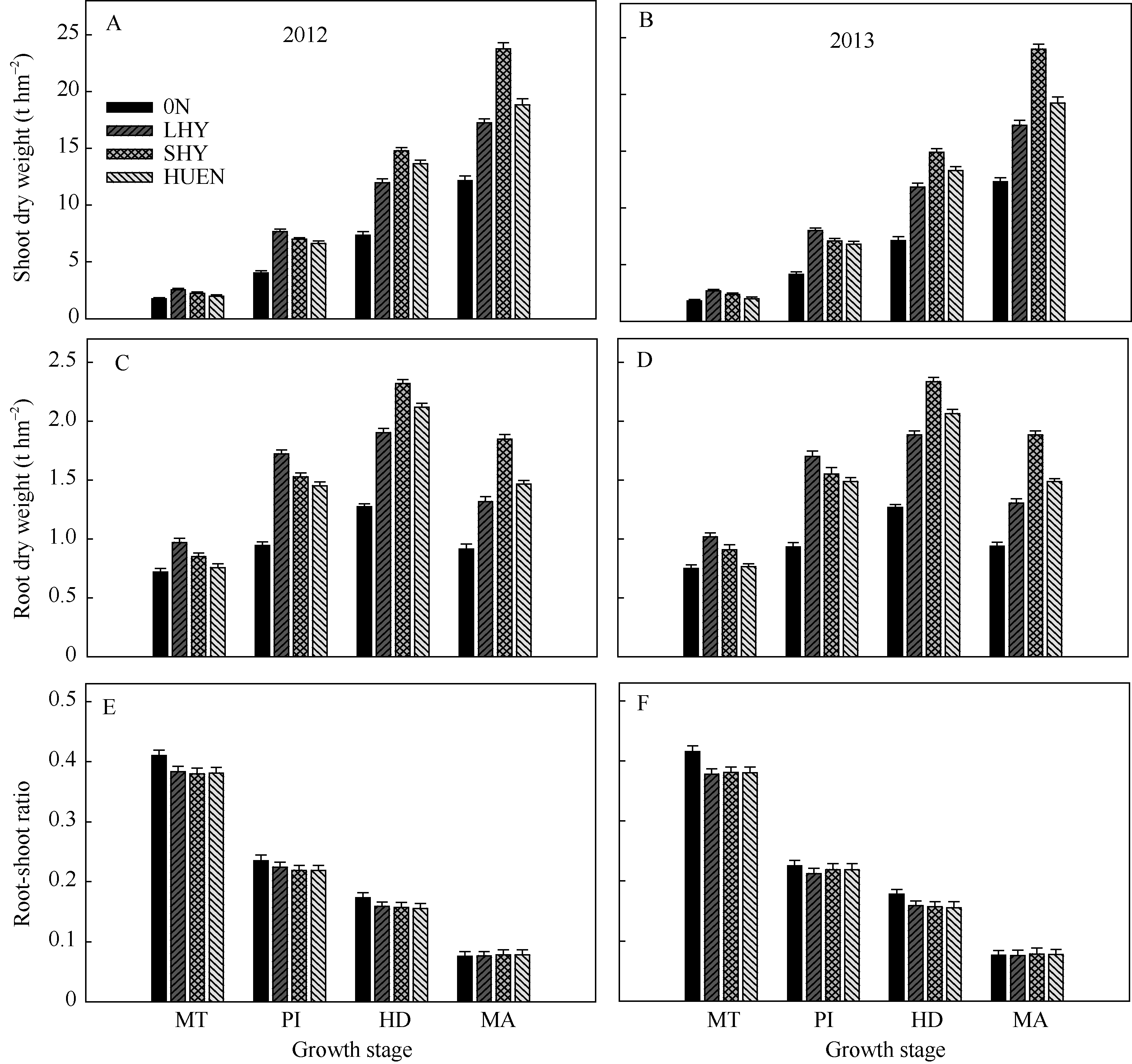 | 图2 不同栽培模式下的水稻在不同生育时期地上部干物重(A, B)、根干重(C, D)和根-冠比(E, F)Fig. 2 Shoot dry weight (A, B), root dry weight (C, D), and root-shoot ratio (E, F) under different cultivation patterns in rice at different growth stages |
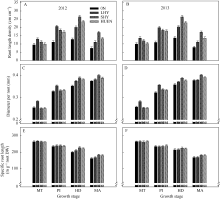
0N处理的根长密度(图3-A, B)与根直径(图3-C, D)均显著低于其他3个处理。在分蘖中期与穗分化始期, 当地高产栽培(对照)的根长密度、根直径以及比根长(图3-E, F)均显著高于超高产栽培或氮肥高效利用栽培。而在抽穗期与成熟期, 超高产栽培或氮肥高效利用栽培的根长密度及比根长显著高于对照; 在上述两个时期, 超高产栽培的根直径显著高于对照, 而氮肥高效利用栽培下的水稻根直径与对照无显著差异。
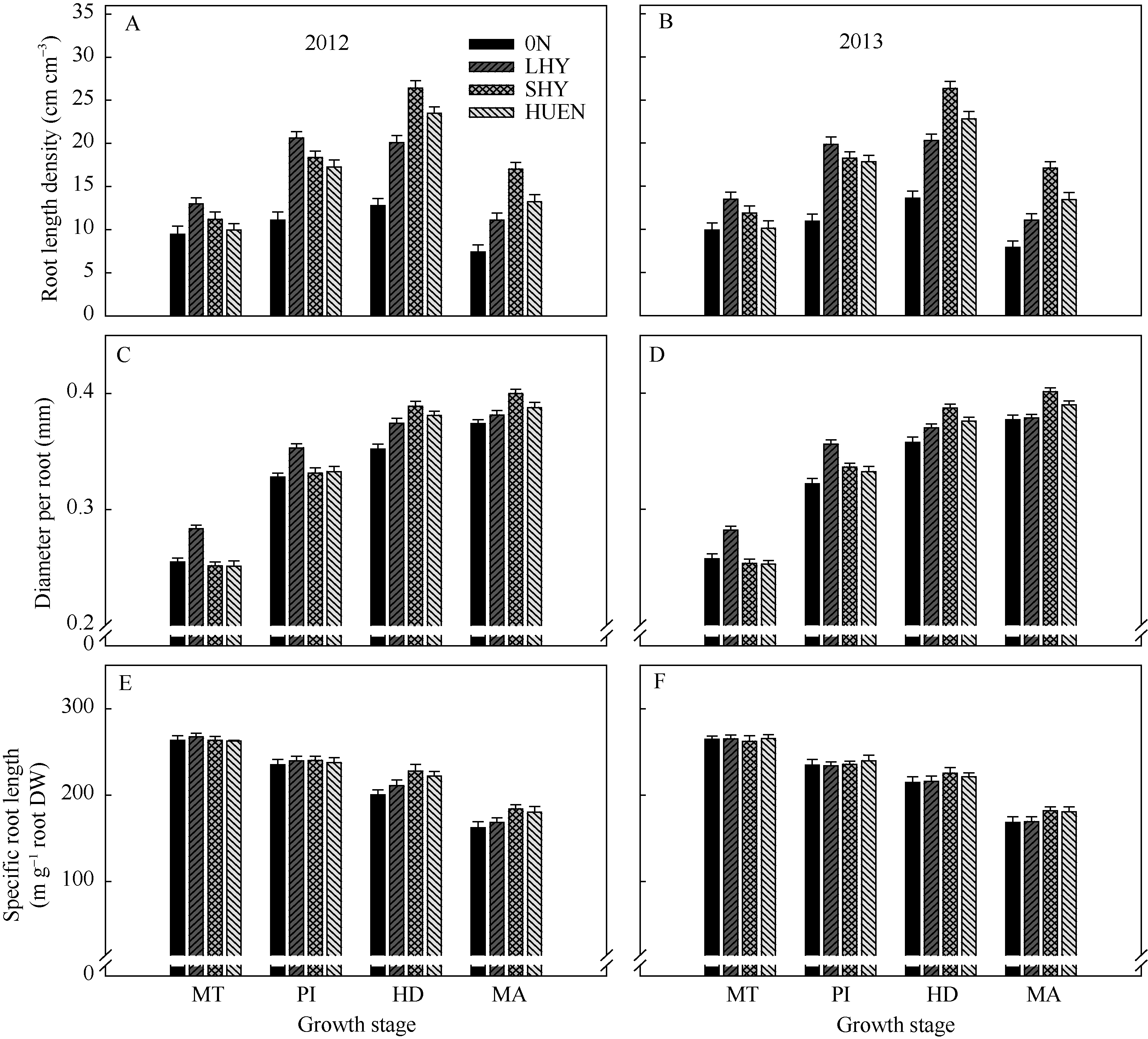 | 图3 不同栽培模式下的水稻在不同生育时期根长(A, B)、根直径(C, D)和比根长(E, F)Fig. 3 Root length (A, B), diameter per root (C, D), and special root length (E, F) under different cultivation patterns in rice at different growth stages |
在整个灌浆期, 超高产栽培、氮肥高效利用栽培的根系氧化力(图4-A, B)均显著高于0N处理及当地高产栽培(对照)。超高产栽培、氮肥高效利用栽培的根系伤流液强度(图4-C, D)所表现的趋势与根系氧化力相一致。
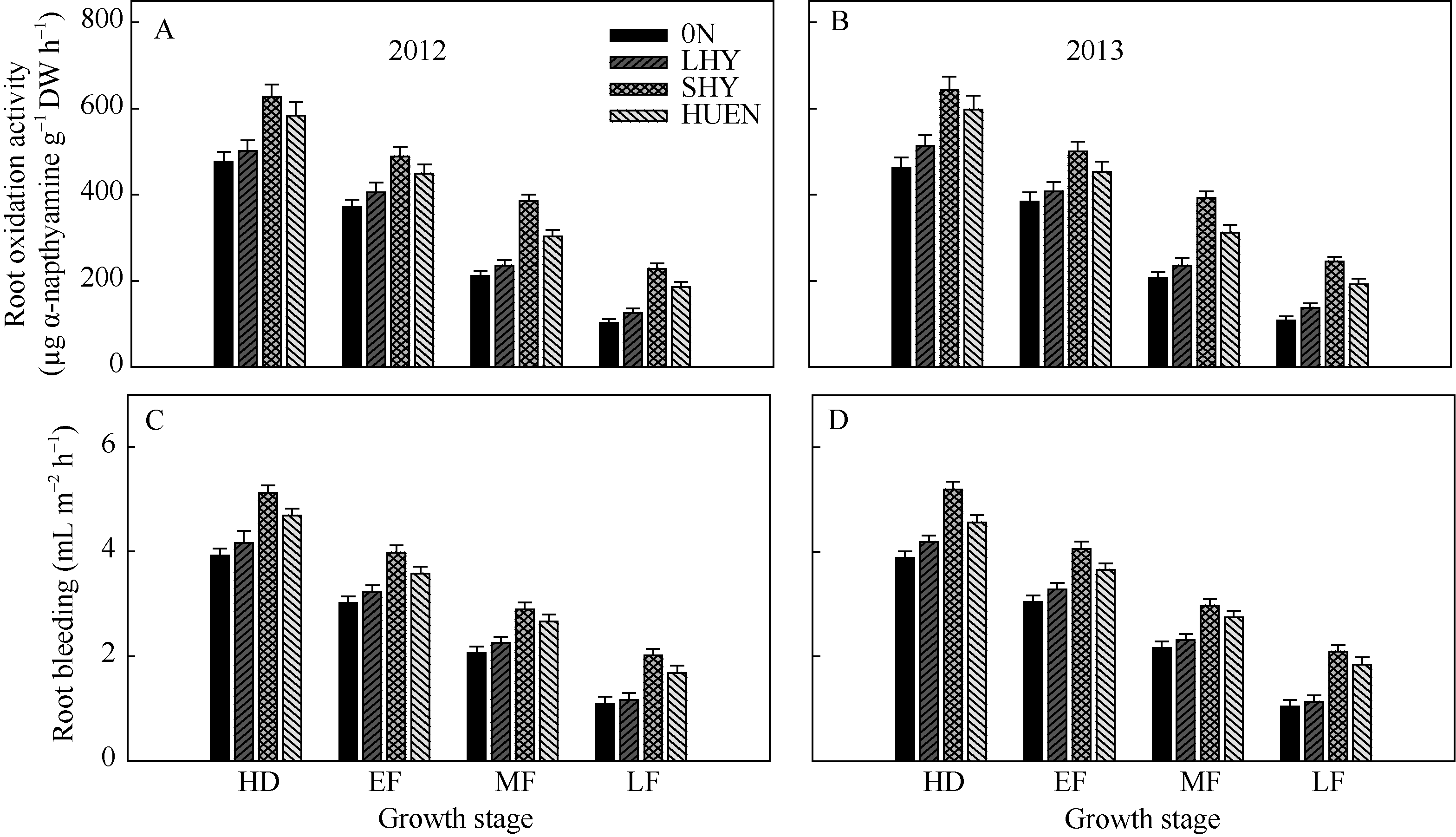 | 图4 不同栽培模式下的水稻在不同生育时期根系氧化力(A, B)和根系伤流液强度(C, D)Fig. 4 Root oxidation activity (A, B) and root bleeding amount (C, D) under different cultivation patterns in rice at different growth stages |
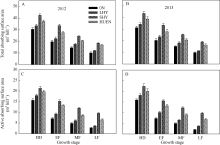
在整个灌浆期, 不同栽培处理下的水稻根系总吸收表面积(图5-A, B)和根系活跃吸收表面积(图5-C, D)均呈下降趋势, 超高产栽培、氮肥高效利用栽培均显著高于0N处理及当地高产栽培(对照); 抽穗期至灌浆后期, 超高产栽培或氮肥高效利用栽培的根系活跃吸收表面积下降幅度分别为37.39%、49.19%, 而对照为62.60%。
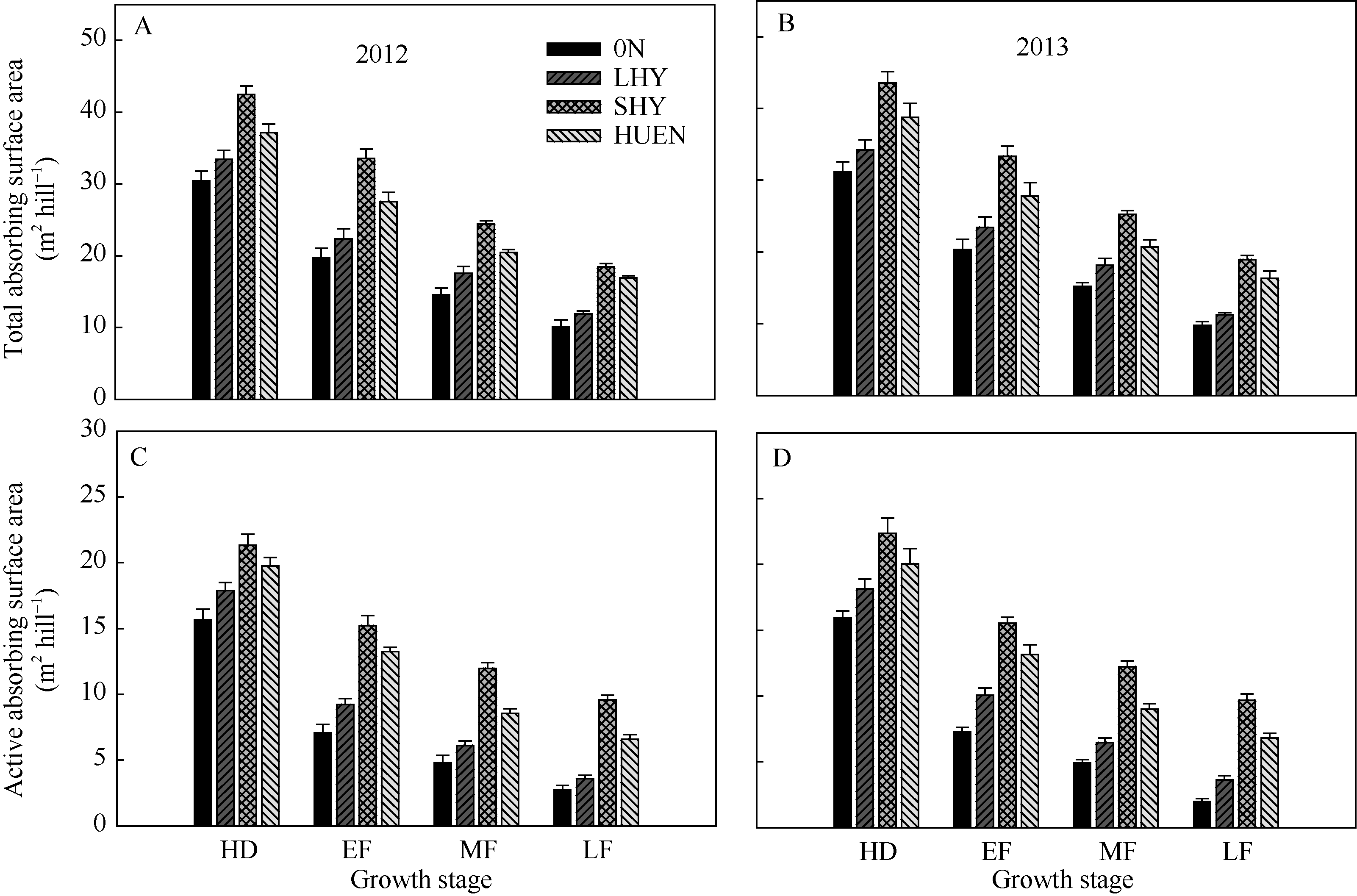 | 图5 不同栽培模式下的水稻在不同生育时期根系总吸收表面积(A, B)和活跃吸收表面积(C, D)Fig. 5 Total absorbing surface area (A, B) and active absorbing surface area (C, D) under different cultivation patterns in rice at different growth stages |
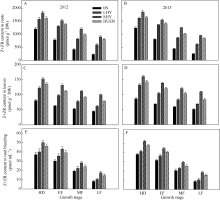
在整个灌浆期, 不同栽培模式下的水稻根系(图6-A, B)、叶片(图6-C, D)以及根系伤流液(图6-E, F)中的玉米素(Z)+玉米素核苷(ZR)含量均呈下降趋势, 超高产栽培以及氮肥高效利用栽培的根系、叶片以及根系伤流液中的玉米素(Z)+玉米素核苷(ZR)含量显著高于0N处理或当地高产栽培(对照)。
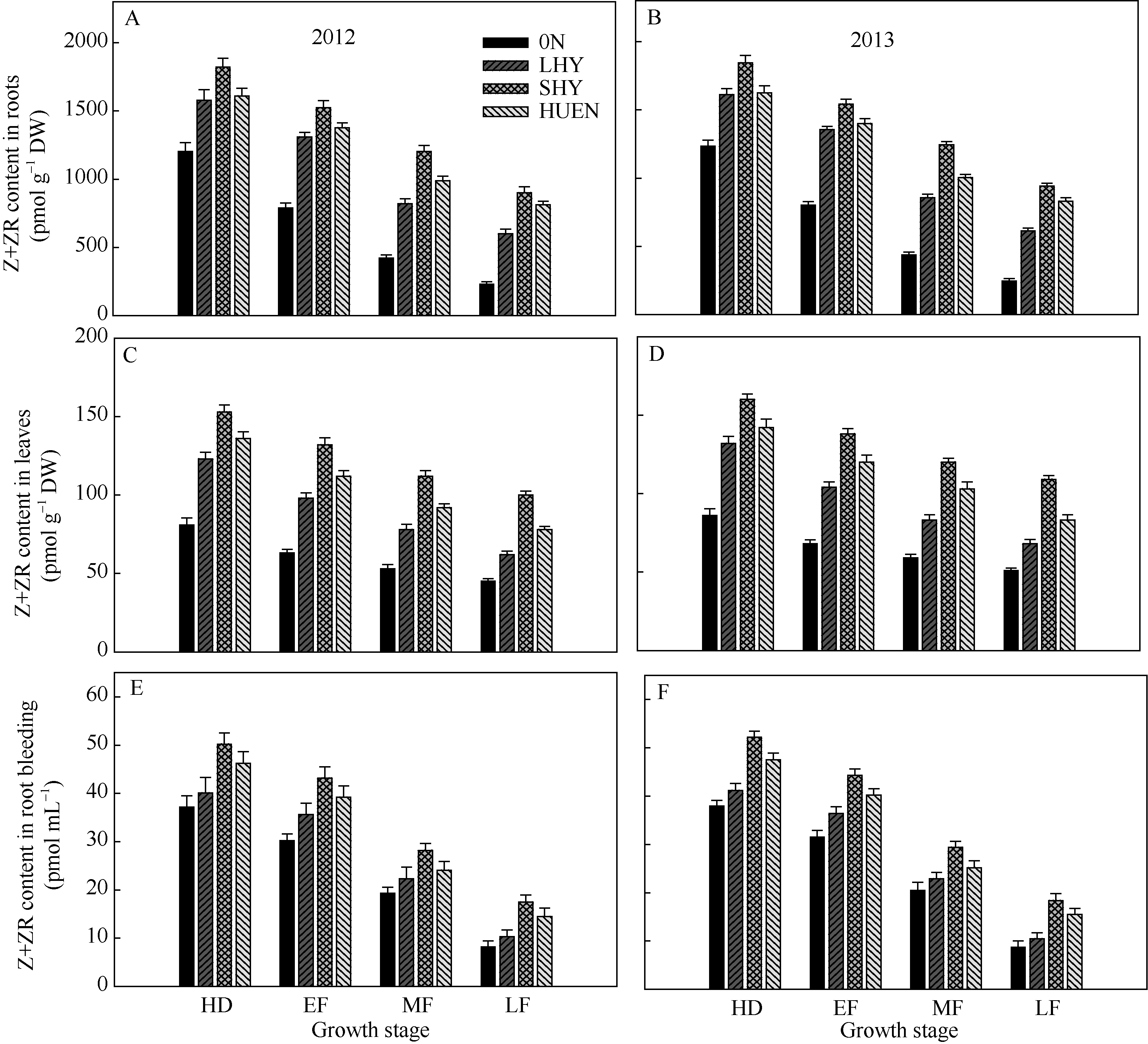 | 图6 不同栽培模式下水稻根系(A, B)、叶片(C, D)和根系伤流液(E, F)中玉米素(Z)+玉米素核苷(ZR)含量Fig. 6 Zeatin (Z) + zeatin riboside (ZR) content in roots (A, B), leaves (C, D), and root bleeding (E, F) under different cultivation patterns in rice at different growth stages |
当地高产栽培籽粒中的蔗糖合酶(SuSase)以及腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)活性在灌浆前期较0N处理低, 在灌浆中期与0N处理差异不显著, 而在灌浆后期则显著高于0N处理; 超高产栽培和氮肥高效利用栽培在整个灌浆期内2种酶活性均显著高于0N处理和当地高产栽培(表6)。
| 表6 各处理的籽粒中蔗糖合酶及腺苷二磷酸葡萄糖焦磷酸化酶活性 Table 6 Activities of sucrose synthase (SuSase) and adenosine diphosphate glucose pyrophospgorylase (AGPase) in grains |
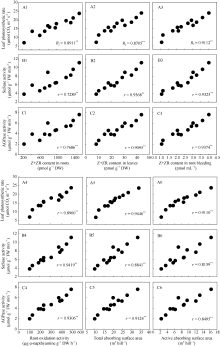
回归分析表明, 在不同栽培模式下水稻灌浆期根系、叶片以及根系伤流液中Z+ZR的含量、根系氧化力以及根系总吸收表面积和根系活跃吸收表面积与灌浆期剑叶净光合速率、籽粒中蔗糖合酶(SuSase)以及腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)活性均呈极显著正相关关系( r = 7289**~0.9419**)(图7)。
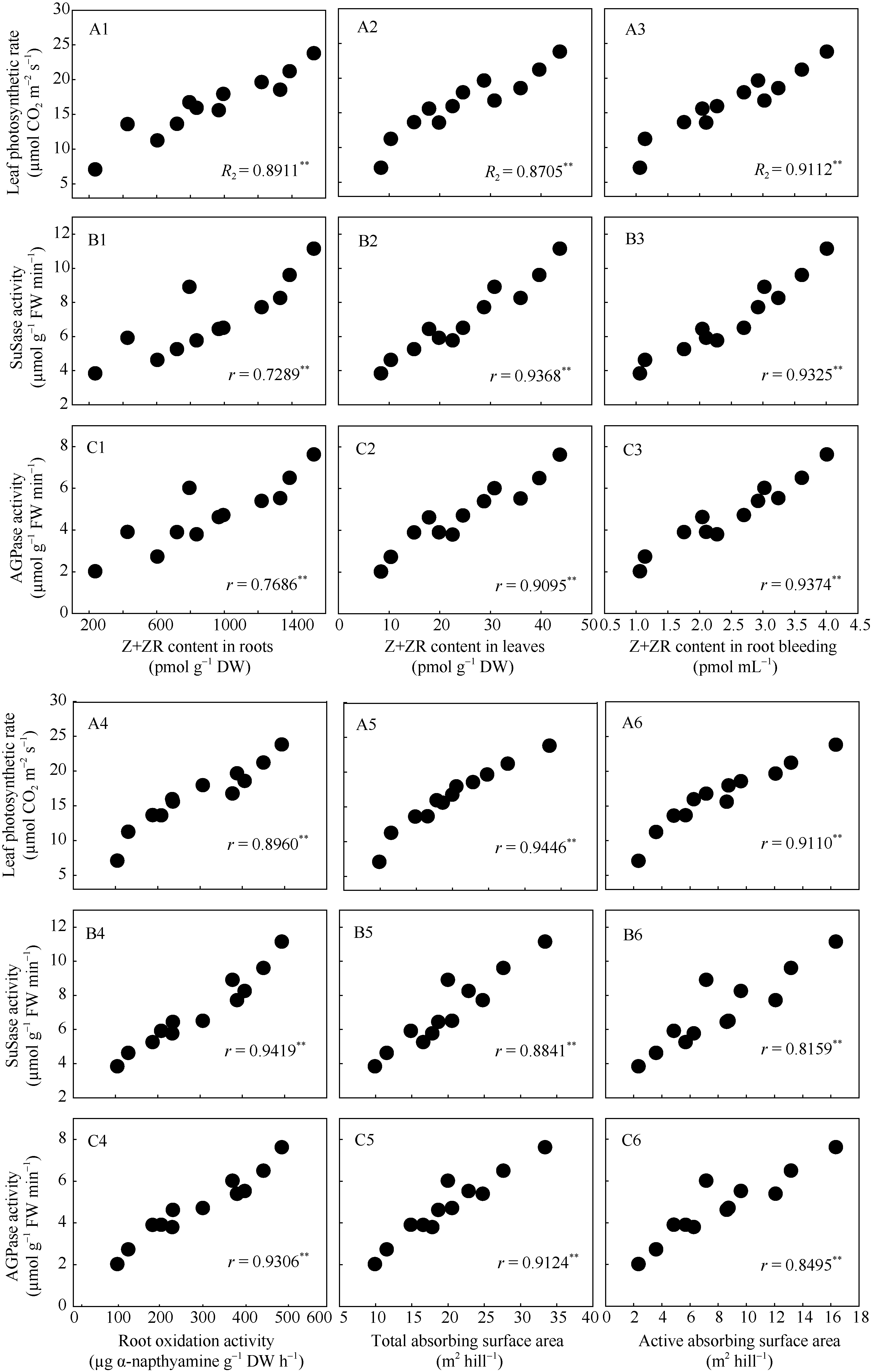 | 图7 水稻剑叶净光合速率(A1~A6)、蔗糖合酶活性(B1~B6)和腺苷二磷酸葡萄糖焦磷酸化酶活性(C1~C6)与根系(A1~C1)、叶片(A2~C2)和根系伤流液(A3~C3)中Z+ZR含量、根系氧化力(A4~C4)、根系总吸收表面积(A5~C5)以及根系活跃吸收表面积(A6~C6)之间的关系Fig. 7 Correlation of flag leaf photosynthetic rate (A1-A6), the activities of sucrose synthase (SuSase) (B1-B6) and adenosine diphosphoglucose pyrophosphorylase (AGPase)(C1-C6) with zeatin (Z) + zeatin riboside (ZR) content in roots (A1-C1), leaves (A2-C2) and root bleeding (A3-C3), root oxidation activity (A4-C4), total absorbing surface area (A5-C5), and active absorbing surface area (A6-C6) |
持续提高作物产量是否必须依赖于养分资源的大量投入?作物高产与资源高效利用能否协同?这一直是国内外关注的热点, 也是学术界仍在争论的重大科学命题[ 23, 24, 25]。根系作为水稻养分吸收和运输的主要器官, 其功能的发挥与根系形态和生理特性密切相关[ 26]。一般认为, 高产与氮高效水稻根系具有发根力强、根系发达; 根系生物量、根系体积、根系总吸收表面积和活跃吸收表面积大; 粗分支根比例、根系伤流强度、根系氧化力高; 对NH4+的亲和力强等特征[ 27, 28, 29, 30, 31]。但也有研究认为, 杂交水稻的根系氧化力、根系ATP酶活性、单株根重和吸氮量、单位根重吸氮量均显著高于亲本, 但氮素吸收利用率与亲本无显著性差异[ 32]。本试验结果表明, 在超高产栽培与氮肥高效利用栽培下, 根系生物量、根系氧化力、根系总吸收表面积与活跃吸收表面积、根系伤流液强度均显著高于当地高产栽培(对照); 根系、叶片以及根系伤流液中Z+ZR含量以及灌浆期籽粒中蔗糖合酶(SuSase)和腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)活性也均显著高于对照。说明通过栽培措施培育健壮根系是水稻高产与水分养分高效利用的重要途径。
以往研究表明, 作物根-冠之间存在着显著的互作效应[ 33, 34, 35]。本研究观察到, 超高产栽培与氮肥高效利用栽培显著增加了叶片的光合速率和叶片中细胞分裂素(Z+ZR)含量。地下部的生长(根系氧化力、根系伤流液强度、根系总吸收表面积与根系活跃吸收表面积、根系和根系伤流液中Z+ZR含量)与地上部的生长(叶片光合速率、籽粒中蔗糖淀粉代谢途径关键酶活性)极显著正相关。一方面, 超高产栽培与氮肥高效利用栽培显著增加了整个灌浆期的根系活性, 健壮的根系促进了地上部生物产量和库容的形成, 为地上部提供了充足的养分、水分和植物激素, 改善了地上部的生长; 另一方面, 地上部良好的生长可以保证充足的碳水化合物向根部输送, 为培育健壮根系提供营养保证。说明超高产栽培与氮肥高效利用栽培促进了根系生长, 也有利于其他生理过程, 这是在这两种栽培模式下获得高产高效的重要原因。
通过何种栽培技术可以培育健壮根系, 实现高产高效?这是作物生产上研究的一个热点。有关水稻高产高效栽培技术的研究, 前人已作了较多的报道, 包括群体质量栽培技术、精确施肥技术、精确灌溉技术等[ 36, 37, 38]。本研究在前人研究基础上, 对有效改善根-冠互作效应的栽培技术进行了集成和创新。本试验除采用培育壮秧、增加栽插密度、在氮肥高效利用栽培模式中减少氮肥施用量、前氮后移以及施用硅肥和锌肥等措施外, 关键栽培技术有以下几点。(1)施用含有硝化抑制剂的氮肥。硝化抑制剂可以使土壤中的氮肥较长时间以铵态氮的形式存在, 以减轻氮素的流失, 提高水稻吸氮效率, 实现对氮肥的高效管理与利用[ 39]。在本研究中, 含有硝化抑制剂氮肥的施用显著提高了氮肥利用效率, 提高了灌浆期根系的生理活性, 延缓了根系的衰老, 确保了灌浆期特别是灌浆中后期根系吸收到充足的养分和水分, 以满足地上部生长和籽粒灌浆的需要, 从而为水稻高产稳产奠定基础。(2)有机肥与化肥配施。这是维持和提高地力, 培肥土壤, 提高肥料利用率的重要途径, 长期施用有机肥或化肥有机肥配施, 不仅有利于协调土壤碳、氮库的平衡, 而且可显著提高土壤有机质含量, 进而提高土壤系统的生产力[ 40, 41, 42]。有机肥与化肥的配施可以为水稻根系营造一个良好的土壤环境, 从而促进根系的生长和根系活性的提高。(3)轻干湿交替灌溉。该技术以水稻生育过程中土壤保持水层和自然落干相互交替为特征, 大大减少灌溉用水量, 提高水分利用率。但是干湿交替灌溉对产量的影响因品种、土壤水分条件和灌溉方式而有较大差异。本研究采用了轻干湿交替灌溉技术, 控制土壤的落干程度, 使植物体内水分状况不受严重的影响。该技术不仅提高水分利用效率, 而且有效地提高水稻的根系活性和叶片的光合功能, 提高籽粒中蔗糖淀粉代谢途径关键酶活性, 增加抽穗后的物质生产量, 并促进光合同化物向籽粒的运转[ 43, 44, 45]。从而在扩大库容(增加总颖花量)的基础上, 保持较高的结实率和粒重。
与当地高产栽培相比, 超高产栽培及氮肥高效利用栽培可以有效提高水稻产量、水分以及氮肥利用效率。较好的根系性能(灌浆期较大的根系生物量, 灌浆期较高的根系氧化力、根系吸收表面积与根系活跃吸收表面积、根系伤流液强度、根系中以及根系伤流液中Z+ZR含量)和地上部植株较强的生理活性(抽穗期较大的叶面积指数、抽穗后较高的叶片光合速率和Z+ZR含量、籽粒中较高的SuSase和AGPase活性)是杂交粳稻常优5号在超高产栽培和氮肥高效利用栽培下高产和水分及氮肥高效利用的重要生理基础。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|