
 Essex
Essex and
and{kind=link}
{kind=link}
大豆生物量与产量组分间的相关及关联分析
晁毛妮1, **  , 郝德荣
, 郝德荣2, ** , 印志同3 , 张晋玉1 , 宋海娜1 , 张怀仁1 , 褚姗姗1 , 张国正1 , 喻德跃1, *
, 郝德荣
|
|


* 通讯作者(Corresponding author): 喻德跃, E-mail:dyyu@njau.edu.cn, Tel: 025-84396410, Fax: 025-84395405
第一作者联系方式: E-mail:2011201044@njau.edu.cn
** 对本文有相同贡献.
生物量与后期的籽粒产量存在紧密联系, 是决定作物经济产量的主要因素之一。本研究利用自然群体中的1142 SNP在2年环境下通过全基因组关联分析检测大豆基因组中与生物量及产量组分显著关联的SNP。结果表明: (1)生物量、百粒重和单株籽粒产量在自然群体中存在广泛的表型及遗传变异, 并存在极显著的正相关, 其中生物量与单株籽粒产量之间的相关略高于与百粒重; (2)两年环境下共检测到41、56和29个SNP分别与生物量、百粒重和单株籽粒产量显著关联, 其中仅有6、19和1个SNP在2个环境中都被检测到; (3)共检测到15个SNP同时控制2个或2个以上性状, 其中位于第19染色体上的BARC-029051-06057位点被检测到同时与生物量、百粒重和单株籽粒产量3个性状显著关联, 表明有共同的遗传基础, 同时也解释了性状间相关的遗传原因; (4)鉴定到的多个SNP与先前我们对叶绿素荧光参数及多个环境下产量相关性状的定位结果共位。这些显著关联SNP位点的鉴定, 有助于理解生物量及产量相关性状的遗传机制, 从而促进利用分子标记辅助选择聚合有利基因, 实现未来大豆高产育种计划。
, HAO De-Rong
** Contributed equally to this work
Biomass, one of the main factors that determine the effective economic yield, has an important effect on the final seed yield. In this study, a genome-wide association analysis was conducted to detect key single-nucleotide polymorphisms (SNPs) associated with biomass and yield components using 1142 SNPs in a soybean landraces panel. There existed abundant phenotypic and genetic diversities and significant correlations among biomass and yield components in the population, and the correlation between biomass and seed yield was slightly higher than that between biomass and seed weight. Genome-wide association analysis using a mixed linear model detected 41, 56, and 29 SNPs associated with biomass, seed weight and seed yield respectively. Among them, 6, 19, and 1 SNPs were detected in two environments. In addition, 15 SNPs were found co-associated with two or more different traits and BARC-029051-06057 on chromosome 19 was associated with the three traits, which implies a partially common genetic basis for the three traits. Many SNPs detected in our study were found co-associated with soybean chlorophyll, chlorophyll fluorescence parameters and yield components in our previous study. The identification of these significant SNPs will be helpful to better understand the genetic basis of biomass and yield components, and facilitate the pyramiding of favorable alleles for future high-yield breeding by marker-assisted selection in soybean.
大豆在农业生产中占有重要地位, 是人类重要的植物蛋白源[ 1], 提高大豆产量一直是大豆育种的核心问题[ 2]。生物量和收获指数是决定作物经济产量的2个主要因素[ 3]。以往通过传统的一些育种途径使得作物产量得到了很大提高, 但主要是通过农艺措施与收获指数的提高[ 4, 5, 6], 很少是由于生物量的增加。对世界的两大重要作物水稻与小麦而言, 收获指数的提高已经进入瓶颈阶段, 进一步提高作物产量迫切需要增加作物的生物量[ 7]。因此, 对生物量的遗传变异分析及其影响因素的研究逐渐成为目前育种家们选育高产品种的方向之一。
生物量是植株基因型C、N积累能力、栽培措施、环境因素的综合结果[ 8], 与后期籽粒产量有紧密的相互关系[ 9]。近年来, 大豆育种家已经鉴定许多与产量有关的QTL或SNP, 主要关乎成熟期, 株高, 百粒重和生长习性等与产量相关的性状[ 10, 11, 12, 13, 14, 15], 但关于生育期内大豆生物量积累的研究却很少, 仅有黄中文等[ 16]利用重组自交系群体作过初步研究, 需要强调的是对育种利用而言, 只有那些在多数环境和多数遗传材料中都稳定表达的QTL才具有应用价值[ 17]。产量性状是非常复杂的数量性状, 具有较低的遗传力且涉及大量基因的表达与调控, 更容易受环境影响[ 18]。在不同的遗传材料中, 或不同的环境条件下, 起主导作用的基因(QTL)可能不同, 因此有必要在在不同遗传背景和多环境下, 继续开展生物量和产量相关性状的定位研究。本研究通过关联分析的方法在自然群体中研究大豆生物量与产量相关性状的遗传关系, 分析其相关的遗传基础, 鉴定控制这些产量影响因子的SNP位点, 以期为改良大豆生物量, 改善“源”的状况, 实现大豆产量增加提供有价值的标记信息。
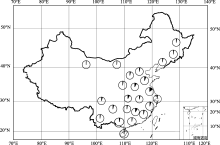
选自全国24个省份不同地理环境、不同生态类型的191栽培大豆地方品种组成的自然群体(图1), 由南京农业大学国家大豆改良中心种质资源库提供。
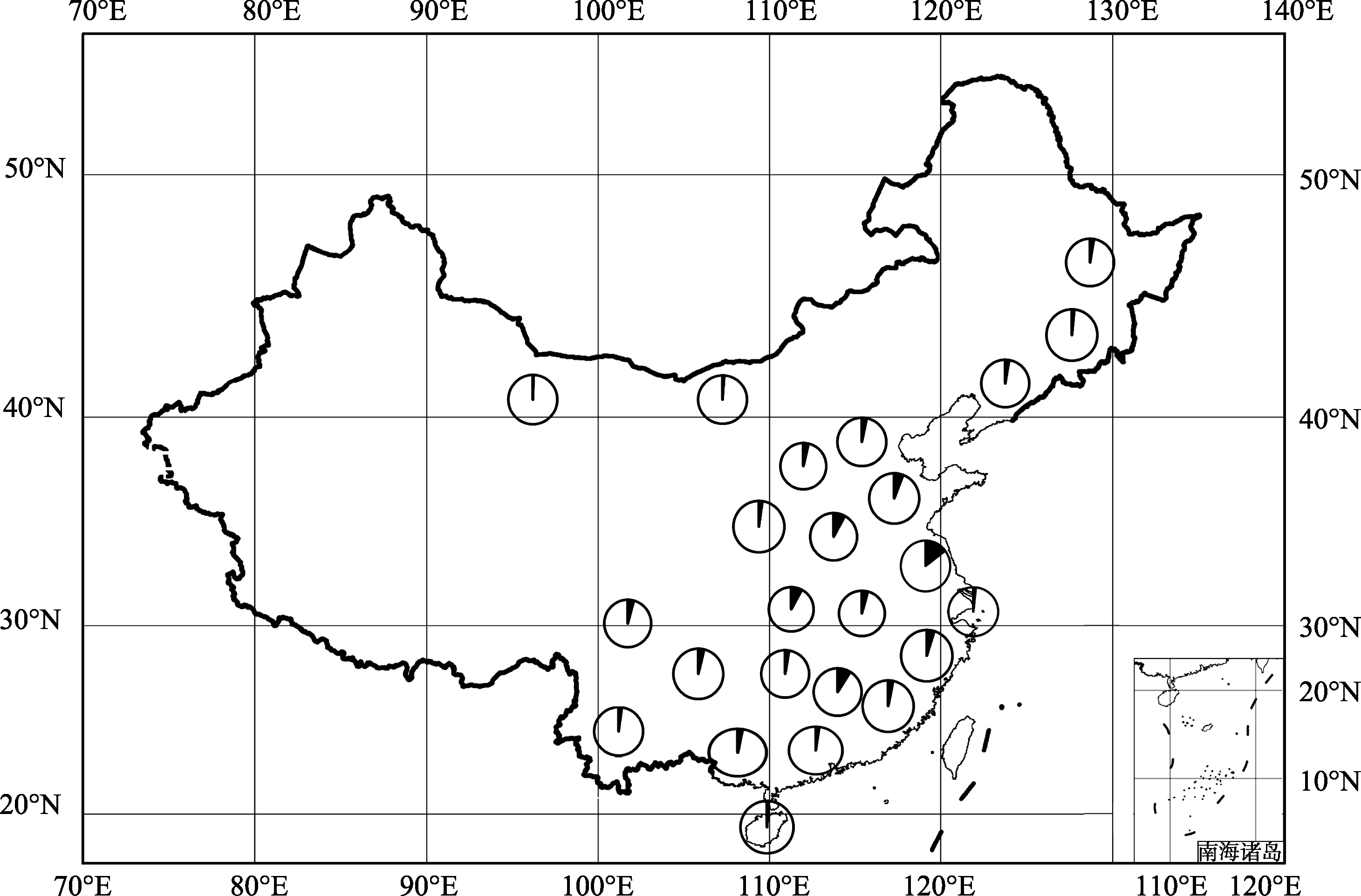 | 图1 191份栽培大豆地方品种地理分布每个圆圈代表不同的省份, 每个省份的材料数目用圆圈的实心部分表示。Fig. 1 Geographical distribution of 191 soybean landraces in our studyCircles represent different provinces, the number of soybean landraces per province is indicated by closed portion of the circle. |
在南京江浦实验站, 采用完全随机设计, 每个品种重复6次, 每重复1盆。为了控制环境对表型性状测定的影响, 根据前3年记载的各材料的熟期, 将试验材料分为4组。分期播种以保证所有材料在采集表型数据时处于类似的生育阶段。2011年播种日期分别为6月7日、17日、27日和7月7日, 2012年为6月8日、18日、28日和7月8日。每盆种9粒, 出苗后7 d, 定苗至2株。在试验材料的生长过程中, 保证充足的营养和水分供应, 以消除营养匮乏和干旱造成的不良影响。
于成熟期收集百粒重和单株籽粒产量的表型数据。成熟后每品种人工收获6株, 手工脱粒并烘干至恒重用于测定百粒重和单株籽粒产量。于鼓粒期收集生物量的数据。每品种选取6株, 将大豆植株整个地上部(包括叶、叶柄、茎秆、根及后期的荚)剪下装入纱袋, 于烘箱中105℃杀青15 min, 85℃烘至恒重 (≥72 h), 称重得生物量。
取大豆苗期幼嫩叶片, 采用改良的CTAB法提取DNA。
从大豆SNP数据库(
在1536个SNP标记中, 有384个标记(25.7%)最小等位基因型频率(minor alleles frequency, MAF)小于10%, 因此本研究选取1142个SNP标记(MAF≥ 10%)用于进一步分析。
用SPSS 17.0的相应程序完成各性状的方差分析、相关分析、描述统计。用TASSEL软件的MLM (mixed linear model)程序完成目标性状的关联分析。MLM将各个体的 Q作为协变量, Kinship值作为随机效应, 将表型数据分别对标记变异进行回归分析, P<0.01时认为该标记与目标性状相关联。群体遗传多样性分析、连锁不平衡估算、群体结构及亲缘关系分析参见郝德荣等[ 15]的方法。
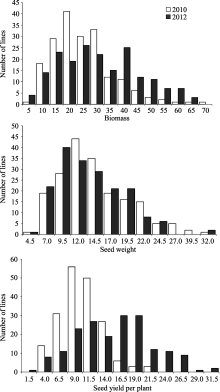
由表1可以看出3个性状在两年环境中均表现出广泛的表型变异, 且品种间差异极显著, 说明现有品种(系)在生物量及产量组分上存在较大的遗传差异, 这些参数的遗传变异可为大豆产量 的改良提供广泛的空间。另外, 生物量与产量组分在大豆品种(系)中, 均呈现连续性分布, 近似正态分布(图2), 表明生物量及产量组分可能是受多基因控制的数量性状。数量性状易受环境影响, 在对生物量及产量组分2个环境联合方差分析时, 发现这3个性状的基因型与环境互作均达到极显著水平。
| 表1 大豆自然群体中生物量与产量组分的描述统计 Table 1 Descriptive statistics of soybean biomass and yield components in soybean natural population |
各性状之间存在极显著相关, 且在不同年份中生物量与百粒重和单株籽粒产量的相关存在相同的趋势即生物量与单株籽粒产量之间的相关略高于与百粒重(表2)。在2011年, 最高的相关系数存在于单株籽粒产量与百粒重之间, 而在2012年最高的相关则存在于单株籽粒产量和生物量之间, 这种差异可能是由于生物量和单株籽粒产量遗传力较低, 易受环境影响, 而百粒重在不同环境下相对稳定。
| 表2 大豆生物量与产量组分间的相关系数 Table 2 Correlation coefficients between biomass and yield components in soybean natural population |
为矫正群体结构和遗传亲缘关系对关联分析的影响, 采用MLM(Q+K)模型对大豆生物量及产量组分与1142个SNP进行了全基因组关联分析。利用1142个SNP标记, 在 P≤0.01 (-lg P≥2.00)水平, 两年共检测到41、56和29个SNP与生物量、百粒重和单株籽粒产量显著关联(表3)。在所有检测到的SNP位点中, 大部分位点仅在特定环境中被检测到, 只有少部分在2个环境中均与生物量或产量组分关联。其中与生物量显著关联的SNP位点中, 有6个在2年环境中均被检测到; 与百粒重显著关联的SNP位点中, 有19个SNP在两年环境中均被检测到; 与单株籽粒产量显著关联的SNP位点中, 很少检测到在2年环境中都存在的SNP, 只有1个SNP在2年环境中均能被检测到。这与产量性状易受环境影响, 需要多年多点实验研究的结果一致。
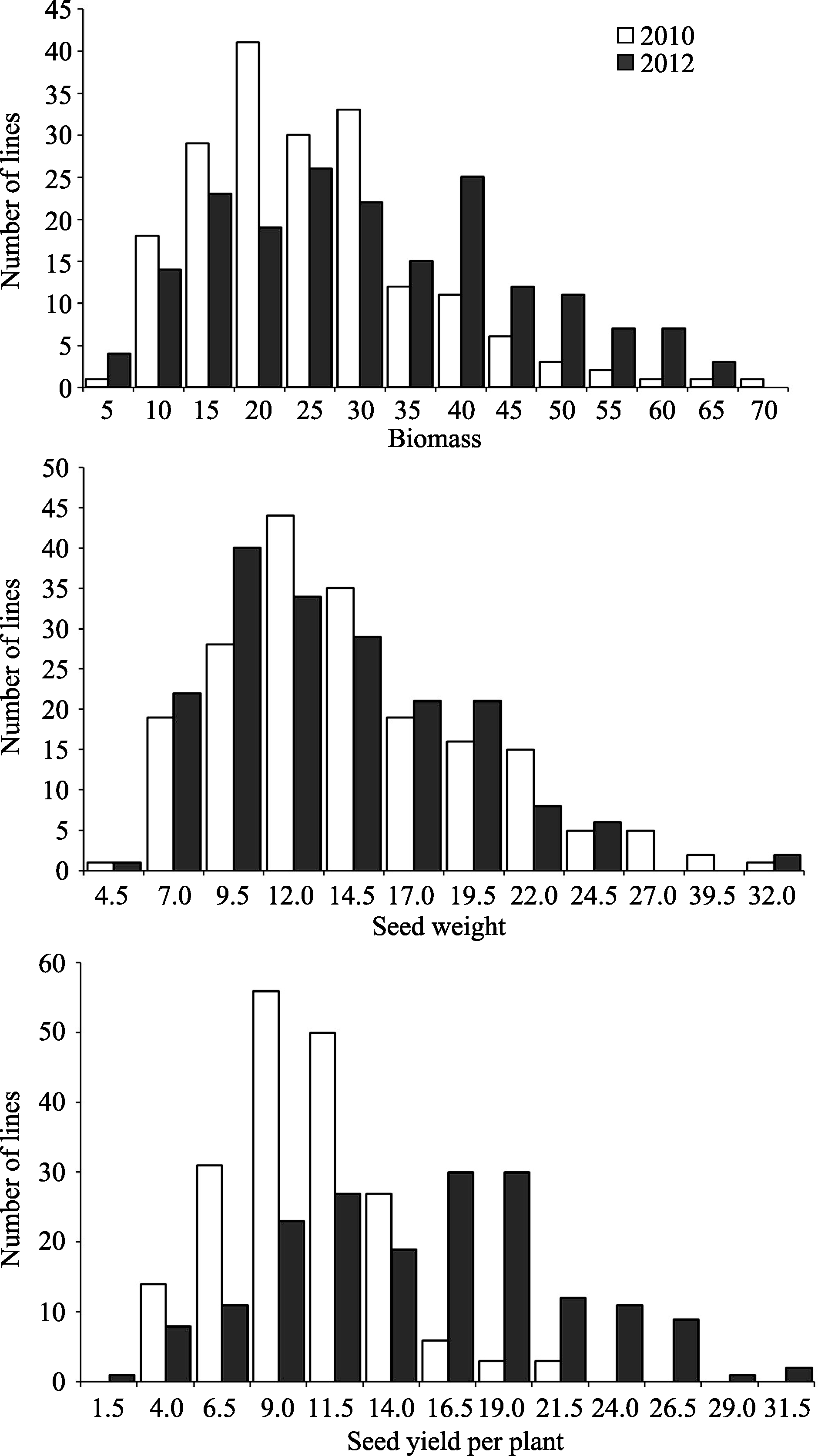 | 图2 大豆品种(系)生物量、百粒重和单株籽粒产量的频率分布Fig. 2 Frequency distribution of biomass, 100-seed weight, and seed yield for soybean varieties (lines) |
共检测到15个SNP同时控制2个或2个以上的性状: (1)与生物量关联的SNP中, 位点BARC- 013333-00485具有较低的 P值, 在2年环境中均能被检测到且与单株籽粒产量共位; 位点BARC- 029051-06057在2年环境均能被检测到, 且同时控制生物量、百粒重和单株籽粒产量3个性状; 位点BARC-030807-06945与单株籽粒产量共位, 但仅在一个环境中被检测到。这3个SNP可能包含同时控制生物量及产量组分的基因。(2)与百粒重关联的SNP中, 共有6个同时控制2个或2个以上的性状, 且均与单株籽粒产量共位。其中位点BARC- 028709-05992在两年环境中均被检测到与单株籽 粒产量共位, 对该位点的继续研究有助于鉴定同 时控制百粒重和单株籽粒产量的基因; 位点BARC- 029051-06057仅在一个环境被检测到, 但是同时与生物量和单株籽粒产量共位, 这个位点可能存在同时控制3个性状的基因; (3)与单株籽粒产量关联的SNP中, 有6个同时控制2个或2个以上的性状, 且大部分均在一个环境中被检测到与其他性状共位, 只有位点BARC-028709-05992在2年环境中均被检测到与百粒重共位。
参照郝德荣等对叶绿素荧光参数[ 19]及对多个环境产量相关性状[ 15]定位结果: (1)与生物量关联的SNP位点中, 3个与叶绿素含量共位, 2个与多个环境下被检测到与单株荚数和单株粒数的SNP共位。 (2)与百粒重关联的SNP位点中, 8个与叶绿素含量和快速叶绿素荧光参数显著关联的SNP共位; 8个与多个环境下的产量相关性状定位结果共位, 且均与多个环境下检测到与百粒重关联的SNP共位; (3)与单株籽粒产量关联的SNP位点中, 4个与叶绿素含量和快速叶绿素荧光参数关联的SNP共位; 1个与多个环境下检测到与单株荚数和单株粒数的SNP共位; 1个与多个环境下检测到与百粒重和产量的SNP共位。
大豆鼓粒期(R6)是决定大豆产量的关键时期[ 20, 21], 此时不仅光合速率及光化学活性高, 而且光合产物也明显增加, 更重要的是这些光合产物大量运往结实器官中[ 21]。张贤泽等[ 20]对大豆鼓粒期的群体光合速率研究发现其和产量呈高度相关, 相关系数 r = 0.9289。黄中文等[ 16]利用重组自交系群体对地下和地上部生物量与产量相关的动态的研究发现, 随生长进程, 相关系数逐渐增加, 至鼓粒期达到最大。但是关于生物量与产量之间也存在不相关的报道[ 22], 一个可能原因是所用的材料或群体大小不同。因此有必要在具有更大生物量和产量变异范围群体中开展这方面的研究。本文发现, 鼓粒期生物量与百粒重及单株籽粒产量呈极显著正相关, 这启示育种者提高鼓粒期生物量是大豆获得高产至关重要的因素, 提高该时期植株的生物量将有助于结实器官中光合产物的积累。
随着二代测序技术的迅速发展, 植物基因组数据海量增加, 显著增加了利用分子标记进行重要性状基因定位的有效性和利用MAS育种的可行性[ 23]。这些技术允许我们系统地分析来自于不同地理区域的品种, 利用高通量的测定工具和全基因组关联分析的方法, 鉴定新的目标基因用于作物改良。为了检测大豆基因组内与生物量, 百粒重和单株籽粒产量相关的SNP, 本研究在自然群体中, 利用关联分析的方法定位研究这些性状。在两年环境下共检测到个41、56和29个SNP与生物量、百粒重和单株籽粒产量显著关联, 其中仅有6、19和1个SNP在2个环境中均被稳定检测到分别与生物量、百粒重和单株籽粒产量显著关联。2年环境下, 与生物量和单株籽粒产量稳定关联的SNP较少可能与其遗传率较低、遗传基础复杂且易受环境影响有关[ 15, 16]。Fulton等[ 24]研究认为, 在多个不同的环境中都能检测到的QTL可能比那些对表型变异表现出高解释率、但只能在特定环境中检测到的QTL更有用。对照郝德荣等[ 15]在5个环境中对产量相关性状定位的研究结果, 3个以上环境与百粒重关联的SNP位点中, BARC- 018515-02927、BARC-018985-03048、BARC- 028709- 05992、BARC-029185-06106、BARC-030931-06978、BARC-040075-07652和BARC-040407-07733与本文在2个环境中均能稳定检测到的与百粒重关联SNP中共位; 3个环境中与产量关联的BARC-028709- 05992位点与在本研究的两个环境中均能检测到与单株籽粒产量显著关联。这些在多年环境中稳定表达的SNP在育种中可能具有较大的利用价值。另外, 本研究共检测到15个SNP同时控制2个或2个以上性状。位于第19染色体上的BARC-029051-06057位点被检测到同时与生物量、百粒重和单株籽粒产量3个性状关联; Thumma等[ 25]指出, 如果两个性状的QTL在染色体上共区间, 则表明这两个性状间很有可能存在因果关系。对这些共位SNP的鉴定表明, 这3个性状间存在紧密的联系, 该结果与3个性状间相关分析结果一致; 同时生物量与百粒重和单株籽粒产量的共位表明控制它们的基因可能存在一因多效现象。共区间内可能含有控制生物量的基因, 该基因的表达可同时影响百粒重及单株籽粒产量。这些结果对于我们通过提高光合性能, 增加鼓粒期生物量来增加作物产量提供了一定的理论基础。
| 表3 自然群体中与大豆生物量及产量组分显著关联的SNP位点 Table 3 SNPs with significant association signals (-lg P≥2.00, P ≤0.01) for soybean biomass and yield components detected in soybean natural population |
| (续表3-1) 自然群体中与大豆生物量及产量组分显著关联的SNP位点 Table 3(Continued) SNPs with significant association signals (-lg P≥2.00, P ≤0.01) for soybean biomass and yield components detected in soybean natural population |
| (续表3-2) 自然群体中与大豆生物量及产量组分显著关联的SNP位点 Table 3-2(Continued) SNPs with significant association signals (-lg P≥2.00, P ≤0.01) for soybean biomass and yield components detected in soybean natural population |
作物生物量的主要决定因素是整个生育期光合速率的累计量, 反映了植物源强度的高低, 也是植物光合效率的体现; 单株荚数和单株粒数则反映了植物库的大小及强度[ 26, 27], 库的强度则直接决定了同化物的分配; 而百粒重和单株籽粒产量事实上是光合有效辐射事件的产物, 是源、库关系协调的最终表现。大豆产量形成的实质是源库流互作的过程[ 27]。在本文鉴定到与生物量相关的SNP中, BARC-030807- 06945和BARC-042035-08159与郝德荣等[ 15]对单株荚数和株粒数的定位结果共位; 这些结果表明源、库之间可能具有共同的遗传基础, 且高度协调来完成整个生长过程,但是也存在单独控制源、流、库的基因或QTL, 对于某些具有较高收获指数的材料, 提高生物量及光合效率可能更有效果。
叶绿素是植物叶绿体内参与光合作用的重要色素之一[ 28], 快速叶绿素荧光参数则在对植物无损伤的情况下更加方便、准确地反映光合器官内各生理生化反应步骤上的能量传递情况[ 29], 因而, 叶绿素含量和快速叶绿素荧光参数均有可能影响光合速率的大小。参照郝德荣等[ 19]对叶绿素荧光参数定位结果, 本研究与生物量关联的SNP位点中BARC- 028289-05814、BARC-029969-06761和BARC- 029969-06762与郝德荣等对叶绿素含量的定位结果共位, 表明这些位点可能包含同时控制叶绿素含量和生物量的基因, 控制叶绿素含量的基因也可能通过影响光合效率而影响作物的生物量; 与百粒重和单株籽粒产量关联的位点中, 存在多个SNP与快速叶绿素荧光参数及叶绿素含量共位。这些位点的存在表明光合效率和大豆产量之间存在紧密联系。另外, 本研究中位点BARC-028709-05992在2个环境中均被检测到与百粒重和单株籽粒产量关联, 且与多个叶绿素荧光参数[ 19]及对多个环境产量相关性状[ 15]定位结果共位, 对该位点的继续研究有助于鉴定同时控制百粒重和单株籽粒产量的及光合相关性状的基因。有明确的证据表明C3植物的最大光合速率在自然群体中存在很大的变异[ 30], 鉴定那些决定这些变异的遗传基础, 将提供新的信息来制定改良作物产量的新策略。对大豆而言 εi(有效辐射的截获效率)和 εp(生物量转换为种子的部分即收获指数)已经最大化( εi = 0.9 / εp = 0.6), 只有很少的改良空间[ 18], 但大豆 εc值(有效辐射转换为生物量的效率)却远远低于理论值。试图改变光合代谢来改变 εc值对于增加大豆生物量及最终实现大豆单株产量的提高是很有意义的。需要指出的是, 光合速率的微小增加(6%~10%)可以导致生物量的巨大增加(30%)[ 31]。因此, 阐明光合作用的机制, 提高光合效率是增加作物产量的一个新目标[ 18]。同时, 高光效种质的发掘及从种质资源库中寻找更有效的等位基因进行遗传转化可能是今后植物高产研究的一个重要方向。
对照前人用连锁分析检测到的与生物量和产量相关性状的QTL, 本研究检测到的SNP位于这些QTL内或与其紧密相邻[ 16, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42]。例如, 本研究检测到第11染色体上2个紧密相邻的SNP位点(BARC- 040407-07733和BARC-040075-07652)在所有环境中均与百粒重显著关联, 这2个标记位于Specht等[ 35]检测到的百粒重QTL位点Sat_095相同的基因组区域; 而位于同一染色体上的与生物量稳定关联的BARC-016279-02316位点则与黄中文等[ 16, 33]检测到的R6期生物量和收获期生物量QTL ( qSBB1-2、 qBMB1-1)处于相同QTL内。与生物量关联的BARC- 022037-04263 (位于第17染色体)则与Kim等[ 34]利用2个重组自交系群体检测到的叶长QTL (Satt372)紧密相邻。这些结果表明, 本研究利用SNP进行的全基因组关联分析可以有效地检测控制大豆生物量及产量组分的QTL。这些一致性SNP/QTL位点将是进一步研究大豆生物量及产量组分遗传机制的候选区域, 对这些关键位点候选基因的挖掘与研究, 对于今后提高大豆的生物量及最终的作物产量将提供重要分子信息。在本研究中, 为了保证试验条件的一致性, 最大程度缩减环境因素对产量的影响, 试验分两年在南京进行, 突出了遗传差异, 所发掘的SNP位点具有普适性。但是, 大豆是光温敏感作物, 将各生态区材料集中在一个地方种植有可能并不能真实地表现出其产量特性, 因此, 在以后的研究中, 为了进一步估计环境因素对结果的影响及鉴定那些在多个环境中能够稳定表达的SNP, 今后仍需在异地及多年环境安排全部材料再做统一鉴定, 以验证本研究结果。
大豆鼓粒期的生物量与百粒重和单株籽粒产量存在极显著的正相关及共同的遗传基础。在自然群体中两年共鉴定到41、56和29个SNP分别与生物量、百粒重和单株籽粒产量显著关联, 其中仅有6、19和1个SNP在2个环境中均被稳定检测到。共检测到15个SNP同时控制2个或2个以上性状。鉴定到的多个SNP与郝德荣等定位的叶绿素荧光参数及多个环境产量相关性状共位, 这些位点的存在对于挖掘产量及光合作用有关的基因提供了有价值的标记信息。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|